Activation of Human Dendritic Cells by Ascophyllan Purified from Ascophyllum nodosum

 , , , and
, , , and 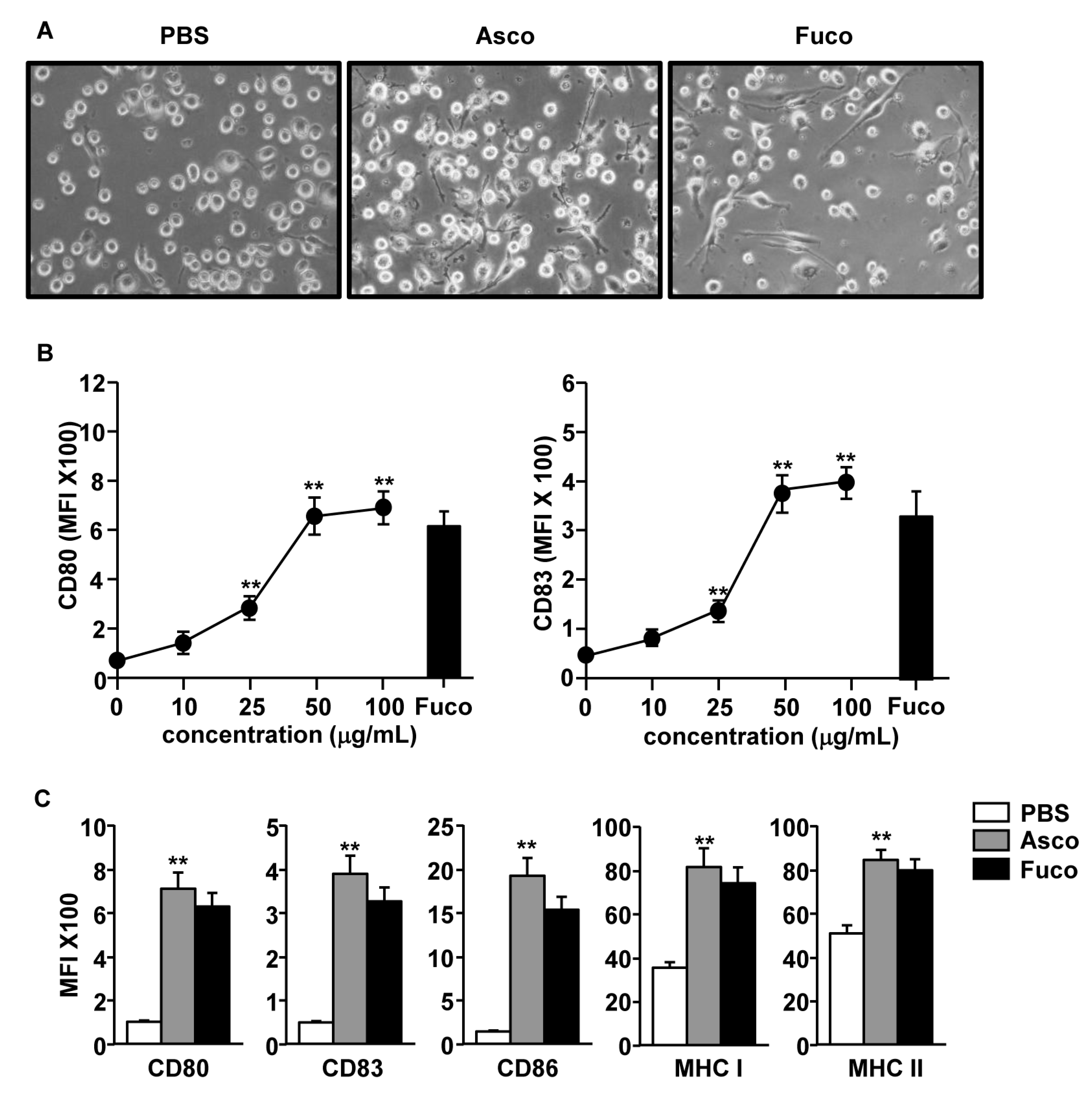{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Ascophyllan from Ascophyllum Nodosum Induced Activation of Monocyte-Derived Dendritic Cells (MDDCs)
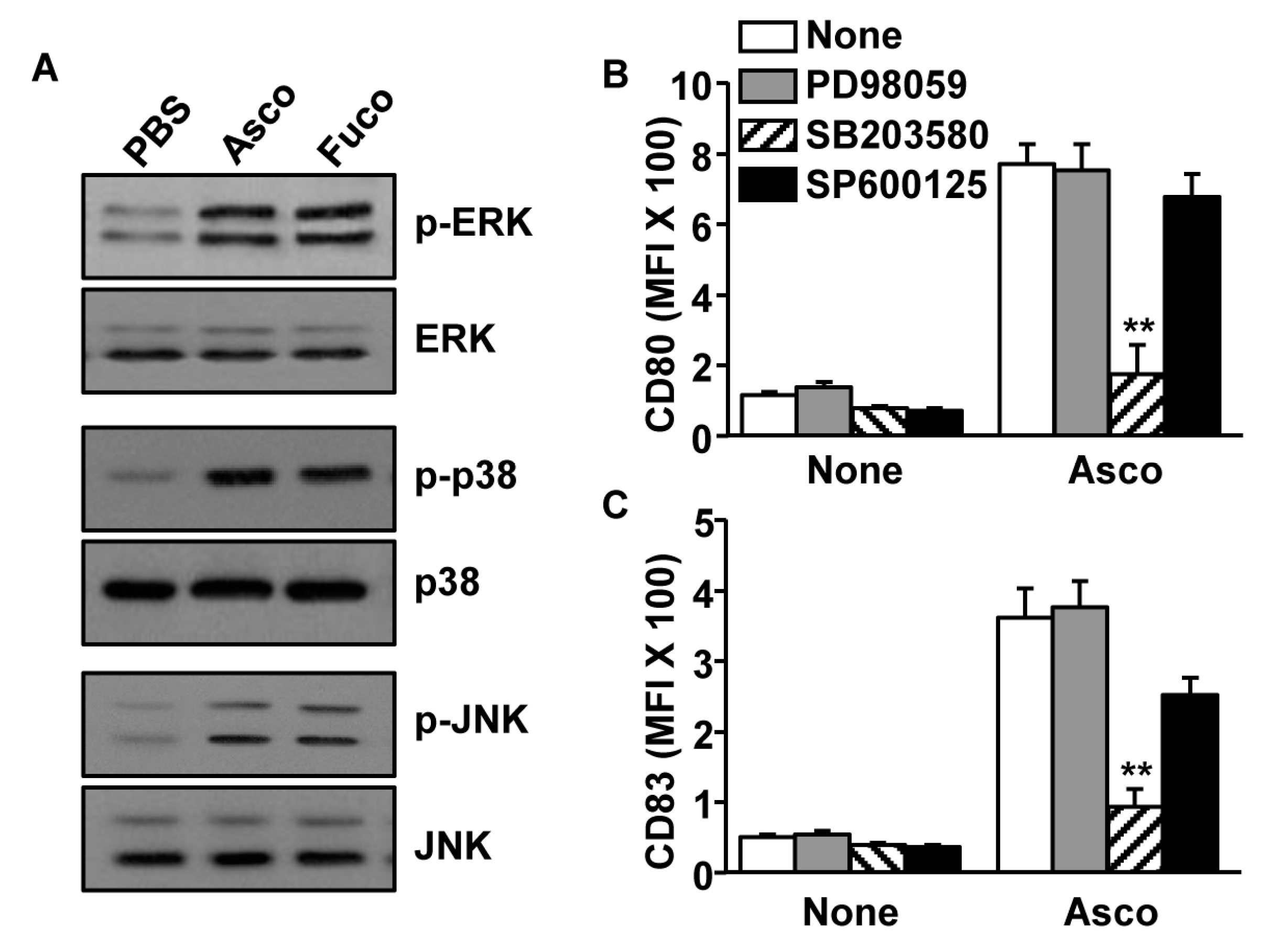
2.2. Activation of MDDCs by Ascophyllan Was Dependent on p38 Phosphorylation
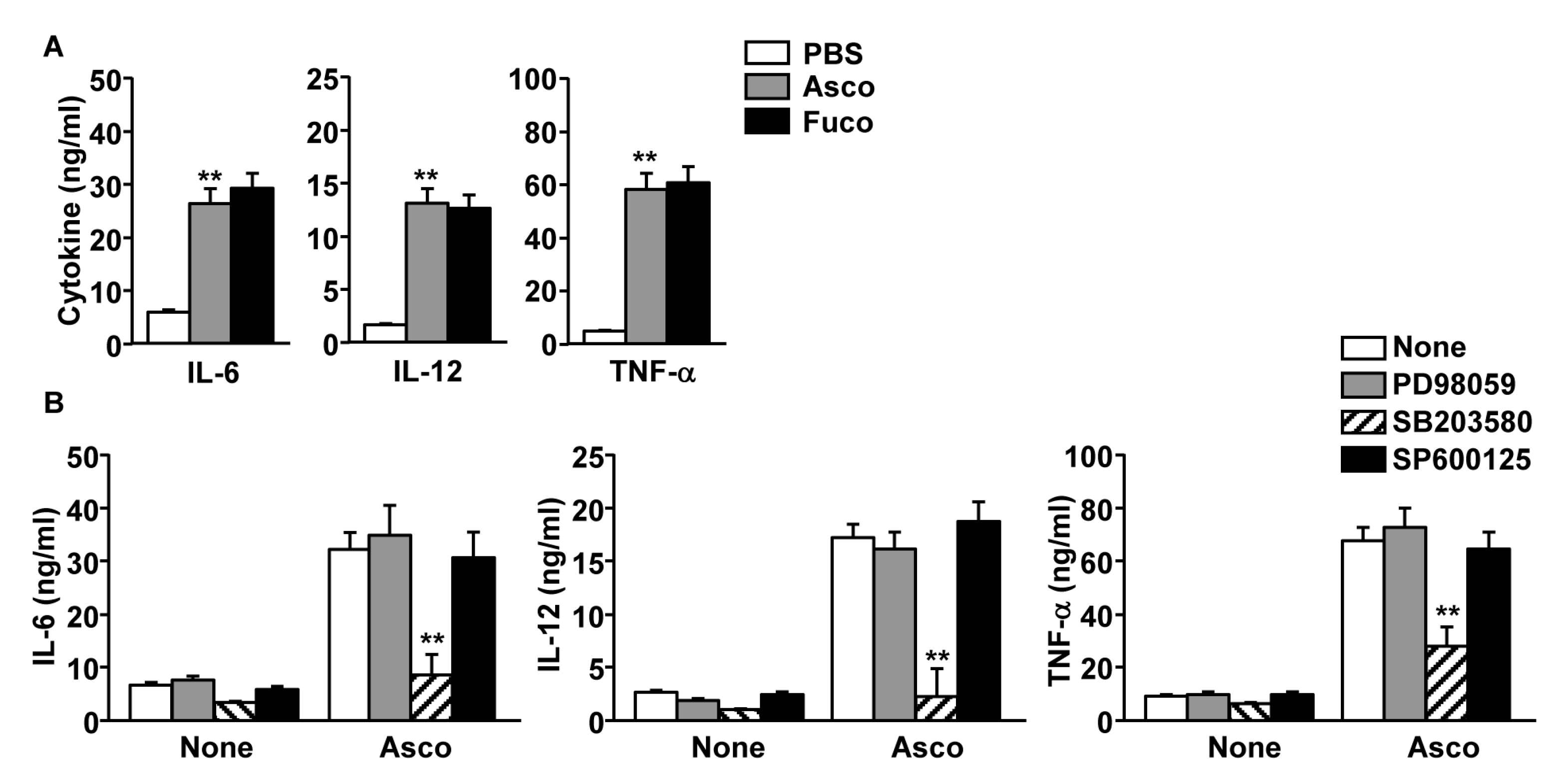
2.3. Production of Pro-Inflammatory Cytokines by MDDCs Was Increased Following Ascophyllan Treatment
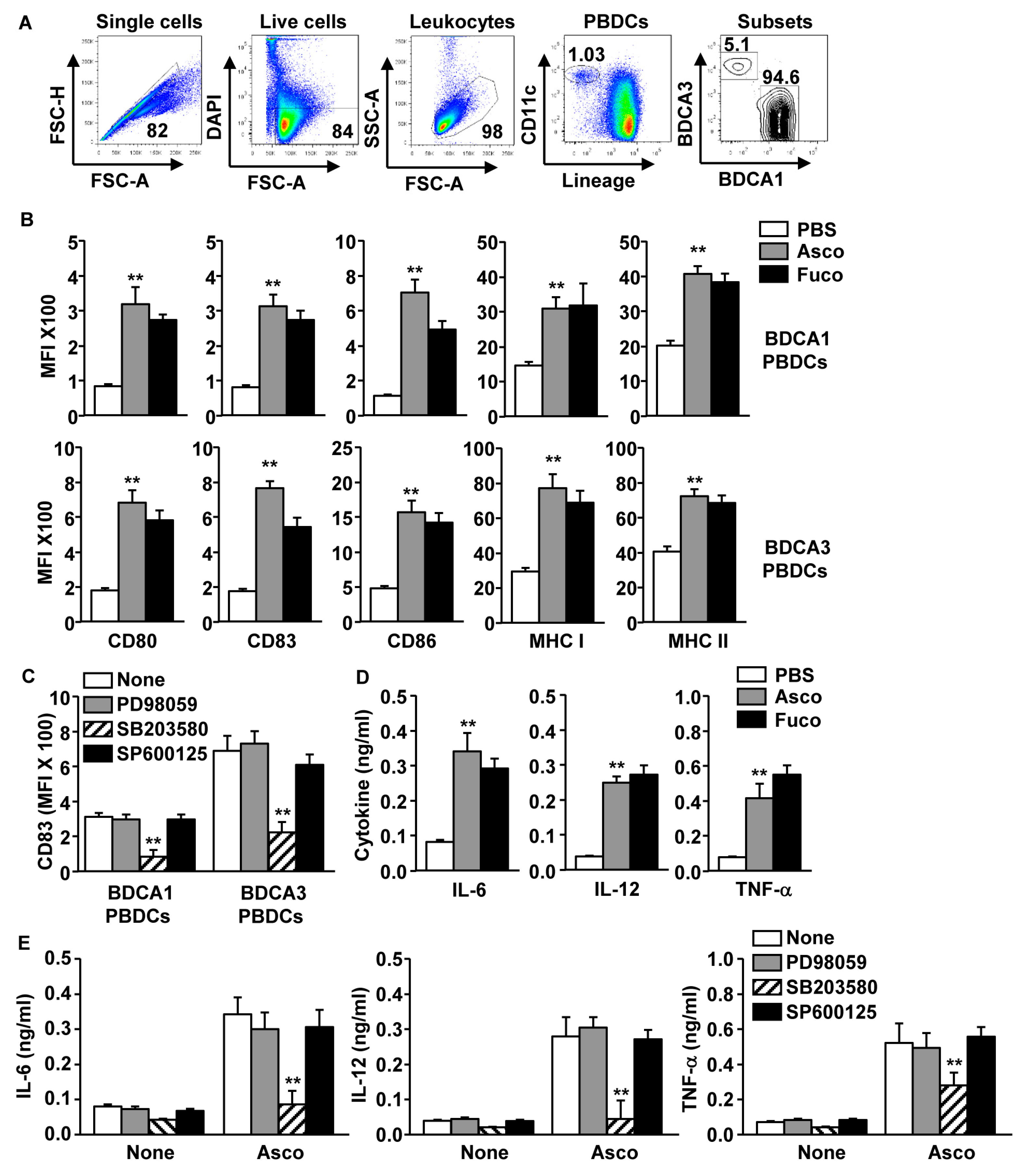
2.4. Ascophyllan-Induced Activation of PBDC Subsets Was Dependent on p38 Signaling
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Chemicals and Cytokines
4.3. Antibodies
4.4. Flow Cytometry
4.5. Differentiation of MDDC and Treatment of Ascophyllan
4.6. Western Blot
4.7. Analysis of PBDCs
4.8. ELISA
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gismondi, A.; Di Marco, G.; Canini, A. Detection of plant microRNAs in honey. PLoS ONE 2017, 12, e0172981. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Kulshreshtha, D.K. Bioactive polysaccharides from plants. Phytochemistry 1989, 28, 2877–2883. [Google Scholar] [CrossRef]
- Luo, M.; Shao, B.; Nie, W.; Wei, X.W.; Li, Y.L.; Wang, B.L.; He, Z.Y.; Liang, X.; Ye, T.H.; Wei, Y.Q. Antitumor and Adjuvant Activity of lambda-carrageenan by Stimulating Immune Response in Cancer Immunotherapy. Sci. Rep. 2015, 5, 11062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, J.Y.; Jiang, Z.; Okimura, T.; Oda, T.; Yu, Q.; Jin, J.O. Ascophyllan purified from Ascophyllum nodosum induces Th1 and Tc1 immune responses by promoting dendritic cell maturation. Mar. Drugs 2014, 12, 4148–4164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.O. Fucoidan from Macrocystis pyrifera has powerful immune-modulatory effects compared to three other fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef]
- Nakano, K.; Kim, D.; Jiang, Z.; Ueno, M.; Okimura, T.; Yamaguchi, K.; Oda, T. Immunostimulatory activities of the sulfated polysaccharide ascophyllan from Ascophyllum nodosum in in vivo and in vitro systems. Biosci. Biotechnol. Biochem. 2012, 76, 1573–1576. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Okimura, T.; Yamaguchi, K.; Oda, T. The potent activity of sulfated polysaccharide, ascophyllan, isolated from Ascophyllum nodosum to induce nitric oxide and cytokine production from mouse macrophage RAW264.7 cells: Comparison between ascophyllan and fucoidan. Nitric Oxide 2011, 25, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayasu, S.; Soegima, R.; Yamaguchi, K.; Oda, T. Biological activities of fucose-containing polysaccharide ascophyllan isolated from the brown alga Ascophyllum nodosum. Biosci. Biotechnol. Biochem. 2009, 73, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Okimura, T.; Xu, L.; Zhang, L.; Oda, T.; Kwak, M.; Yu, Q.; Jin, J.O. Ascophyllan functions as an adjuvant to promote anti-cancer effect by dendritic cell activation. Oncotarget 2016, 7, 19284–19298. [Google Scholar] [CrossRef] [Green Version]
- Cohn, L.; Delamarre, L. Dendritic cell-targeted vaccines. Front. Immunol. 2014, 5, 255. [Google Scholar] [CrossRef]
- Ramberg, J.E.; Nelson, E.D.; Sinnott, R.A. Immunomodulatory dietary polysaccharides: A systematic review of the literature. Nutr. J. 2010, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Dubensky, T.W., Jr.; Reed, S.G. Adjuvants for cancer vaccines. Semin. Immunol. 2010, 22, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Palucka, A.K. Dendritic cells as therapeutic vaccines against cancer. Nat. Rev. Immunol. 2005, 5, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.; Murphy, K.M. Dendritic cell regulation of TH1-TH2 development. Nat. Immunol. 2000, 1, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Millard, A.L.; Mertes, P.M.; Ittelet, D.; Villard, F.; Jeannesson, P.; Bernard, J. Butyrate affects differentiation, maturation and function of human monocyte-derived dendritic cells and macrophages. Clin. Exp. Immunol. 2002, 130, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piemonti, L.; Monti, P.; Sironi, M.; Fraticelli, P.; Leone, B.E.; Dal Cin, E.; Allavena, P.; Di Carlo, V. Vitamin D3 affects differentiation, maturation, and function of human monocyte-derived dendritic cells. J. Immunol. 2000, 164, 4443–4451. [Google Scholar] [CrossRef]
- Tel, J.; Smits, E.L.; Anguille, S.; Joshi, R.N.; Figdor, C.G.; de Vries, I.J. Human plasmacytoid dendritic cells are equipped with antigen-presenting and tumoricidal capacities. Blood 2012, 120, 3936–3944. [Google Scholar] [CrossRef] [Green Version]
- Van der Aa, E.; van Montfoort, N.; Woltman, A.M. BDCA3(+)CLEC9A(+) human dendritic cell function and development. Semin. Cell Dev. Biol. 2015, 41, 39–48. [Google Scholar] [CrossRef]
- Martinez-Cingolani, C.; Grandclaudon, M.; Jeanmougin, M.; Jouve, M.; Zollinger, R.; Soumelis, V. Human blood BDCA-1 dendritic cells differentiate into Langerhans-like cells with thymic stromal lymphopoietin and TGF-beta. Blood 2014, 124, 2411–2420. [Google Scholar] [CrossRef]
- Jin, J.O.; Zhang, W.; Du, J.Y.; Yu, Q. BDCA1-positive dendritic cells (DCs) represent a unique human myeloid DC subset that induces innate and adaptive immune responses to Staphylococcus aureus Infection. Infect. Immun. 2014, 82, 4466–4476. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Park, H.Y.; Xu, Q.; Park, J.I.; Zvyagintseva, T.; Stonik, V.A.; Kwak, J.Y. Ligand of scavenger receptor class A indirectly induces maturation of human blood dendritic cells via production of tumor necrosis factor-alpha. Blood 2009, 113, 5839–5847. [Google Scholar] [CrossRef] [PubMed]
- Langenkamp, A.; Messi, M.; Lanzavecchia, A.; Sallusto, F. Kinetics of dendritic cell activation: Impact on priming of TH1, TH2 and nonpolarized T cells. Nat. Immunol. 2000, 1, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.Y. Fucoidan as a marine anticancer agent in preclinical development. Mar. Drugs 2014, 12, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Zhang, W.; Du, J.Y.; Wong, K.W.; Oda, T.; Yu, Q. Fucoidan can function as an adjuvant in vivo to enhance dendritic cell maturation and function and promote antigen-specific T cell immune responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, L.; Cho, S.Y.; Min, K.J.; Oda, T.; Zhang, L.; Yu, Q.; Jin, J.O. Ginseng Berry Extract Attenuates Dextran Sodium Sulfate-Induced Acute and Chronic Colitis. Nutrients 2016, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cho, S.Y.; Xiang, G.; Min, K.J.; Yu, Q.; Jin, J.O. Ginseng Berry Extract Promotes Maturation of Mouse Dendritic Cells. PLoS ONE 2015, 10, e0130926. [Google Scholar] [CrossRef]
- Handley, M.E.; Rasaiyaah, J.; Barnett, J.; Thakker, M.; Pollara, G.; Katz, D.R.; Chain, B.M. Expression and function of mixed lineage kinases in dendritic cells. Int. Immunol. 2007, 19, 923–933. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Xu, Q.; Kwon, M.J.; Matta, R.; Liu, Y.; Hong, S.C.; Chang, C.H. ERK and p38 MAPK signaling pathways negatively regulate CIITA gene expression in dendritic cells and macrophages. J. Immunol. 2006, 177, 70–76. [Google Scholar] [CrossRef]
- Nakahara, T.; Uchi, H.; Urabe, K.; Chen, Q.; Furue, M.; Moroi, Y. Role of c-Jun N-terminal kinase on lipopolysaccharide induced maturation of human monocyte-derived dendritic cells. Int. Immunol. 2004, 16, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Cannon, M.J.; Goyne, H.E.; Stone, P.J.; Macdonald, L.J.; James, L.E.; Cobos, E.; Chiriva-Internati, M. Modulation of p38 MAPK signaling enhances dendritic cell activation of human CD4+ Th17 responses to ovarian tumor antigen. Cancer Immunol. Immunother. 2013, 62, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Re, F.; Strominger, J.L. Toll-like receptor 2 (TLR2) and TLR4 differentially activate human dendritic cells. J. Biol. Chem. 2001, 276, 37692–37699. [Google Scholar] [CrossRef] [PubMed]
- Colletti, N.J.; Liu, H.; Gower, A.C.; Alekseyev, Y.O.; Arendt, C.W.; Shaw, M.H. TLR3 Signaling Promotes the Induction of Unique Human BDCA-3 Dendritic Cell Populations. Front. Immunol. 2016, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4(+)T cells: Differentiation and functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kwak, M.; Lee, P.C.; Jin, J.O. Rehmannia glutinosa polysaccharide promoted activation of human dendritic cells. Int. J. Biol. Macromol. 2018, 116, 232–238. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Kwak, M.; Park, H.-B.; Okimura, T.; Oda, T.; Lee, P.C.-W.; Jin, J.-O. Activation of Human Dendritic Cells by Ascophyllan Purified from Ascophyllum nodosum. Mar. Drugs 2019, 17, 66. https://doi.org/10.3390/md17010066
Zhang W, Kwak M, Park H-B, Okimura T, Oda T, Lee PC-W, Jin J-O. Activation of Human Dendritic Cells by Ascophyllan Purified from Ascophyllum nodosum. Marine Drugs. 2019; 17(1):66. https://doi.org/10.3390/md17010066
Chicago/Turabian StyleZhang, Wei, Minseok Kwak, Hae-Bin Park, Takasi Okimura, Tatsuya Oda, Peter Chang-Whan Lee, and Jun-O Jin. 2019. "Activation of Human Dendritic Cells by Ascophyllan Purified from Ascophyllum nodosum" Marine Drugs 17, no. 1: 66. https://doi.org/10.3390/md17010066