Protective Effect of Low Molecular Weight Seleno-Aminopolysaccharide on the Intestinal Mucosal Oxidative Damage

Abstract
:1. Introduction
2. Results
2.1. Effects of LSA on Growth Performance
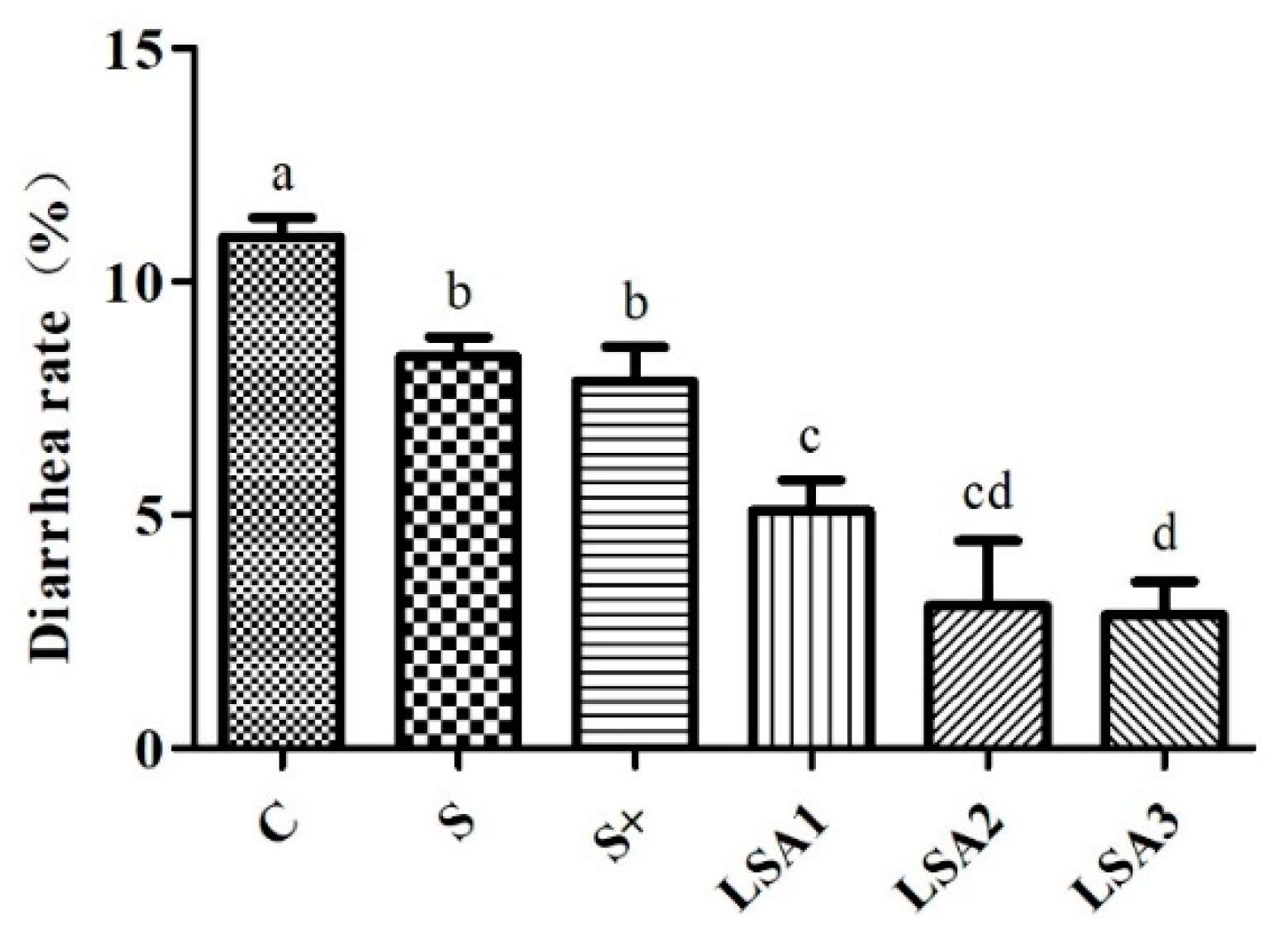
2.2. Effects of LSA on the Diarrhea Rate
2.3. Effects of LSA on Intestinal Morphology
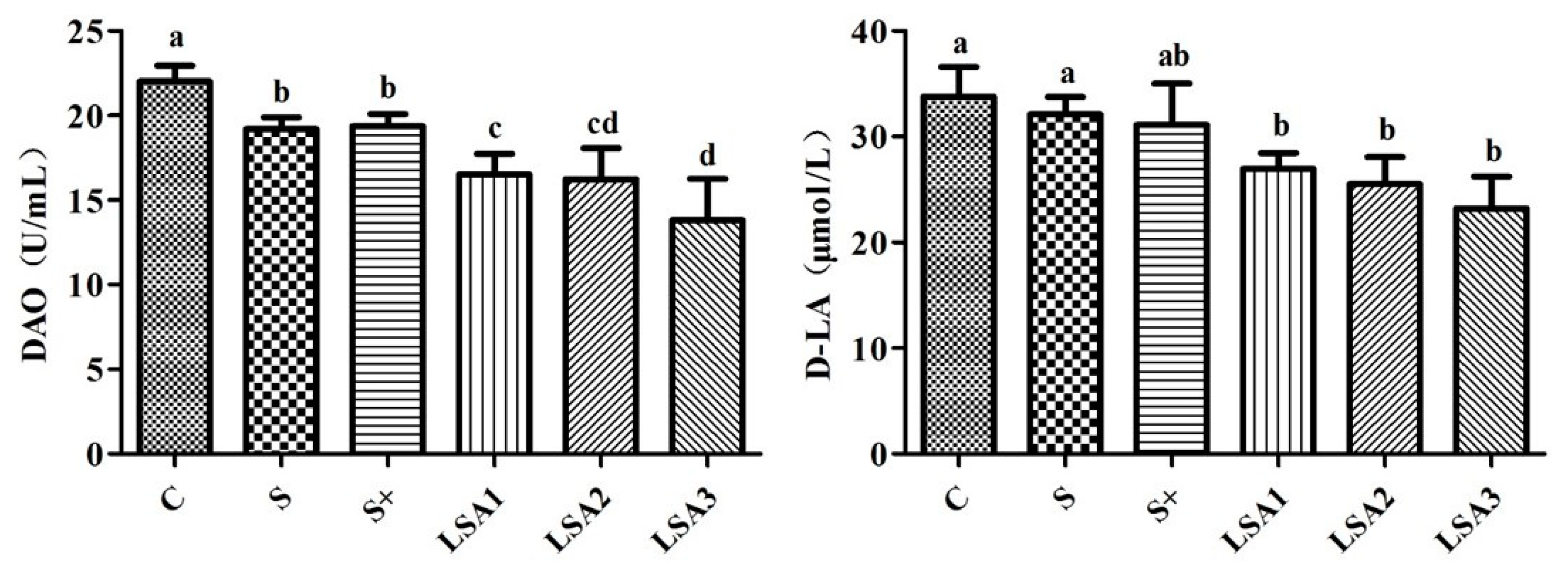
2.4. Effects of LSA on the Level of DAO, d-LA in Serum
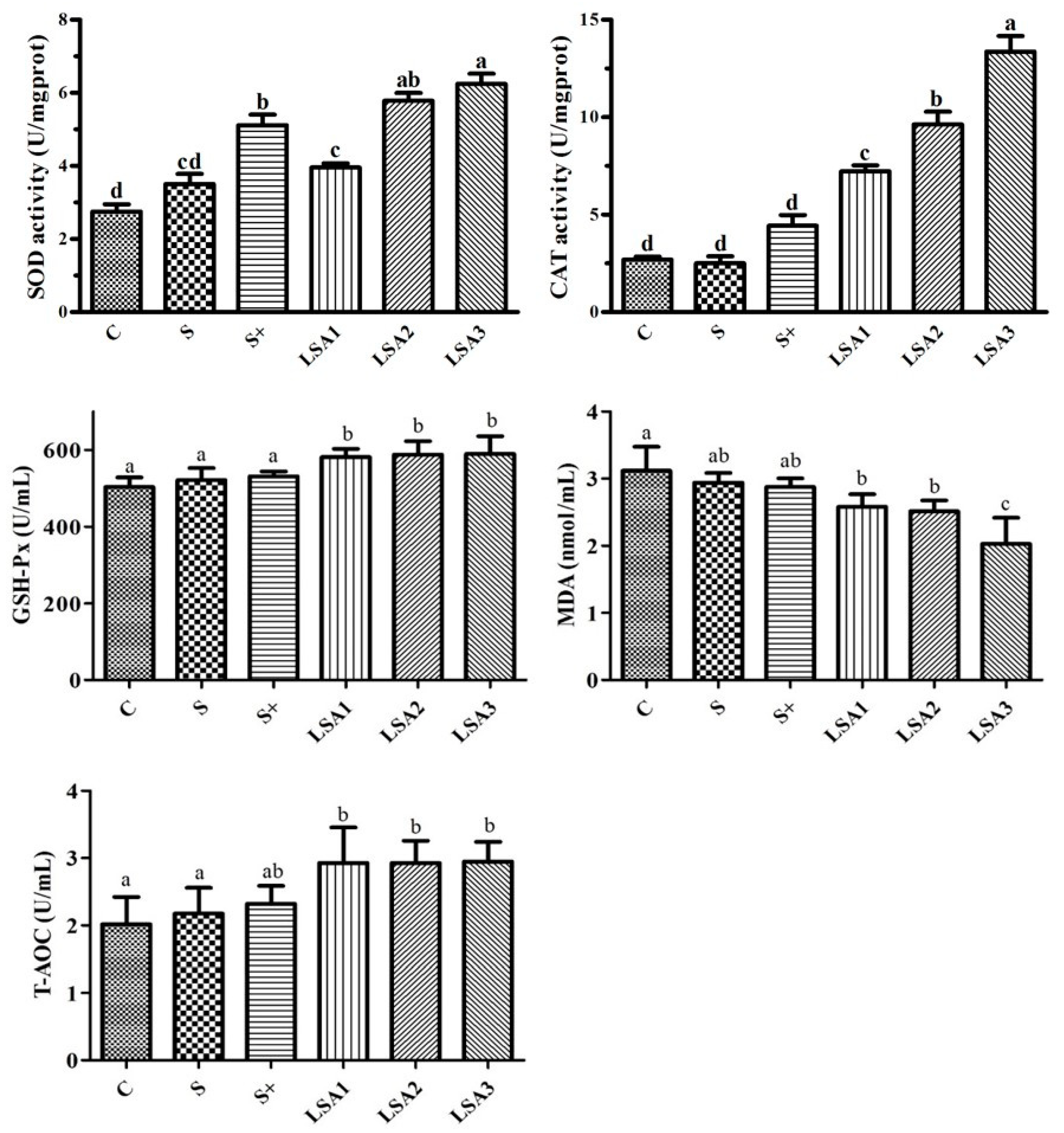
2.5. Effects of LSA on the Antioxidant Levels in Serum
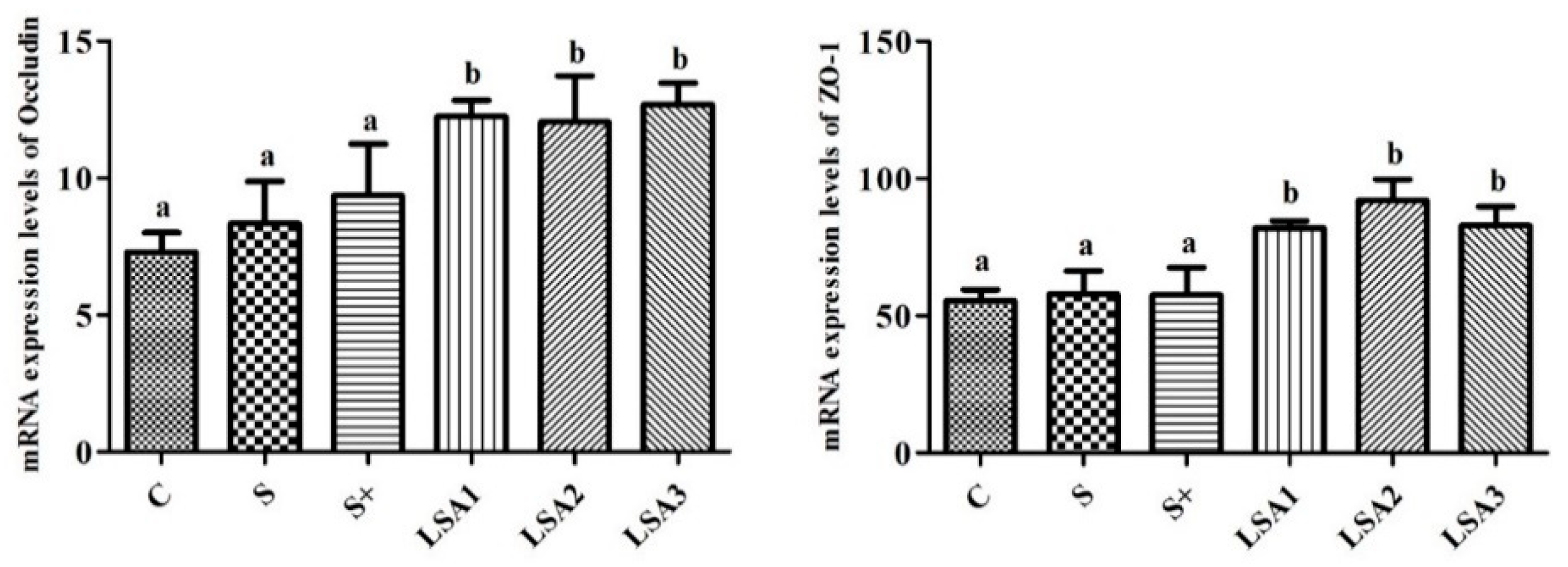
2.6. Effect of LSA on the Gene Expression Levels of Tight Junction Proteins in the Ileum
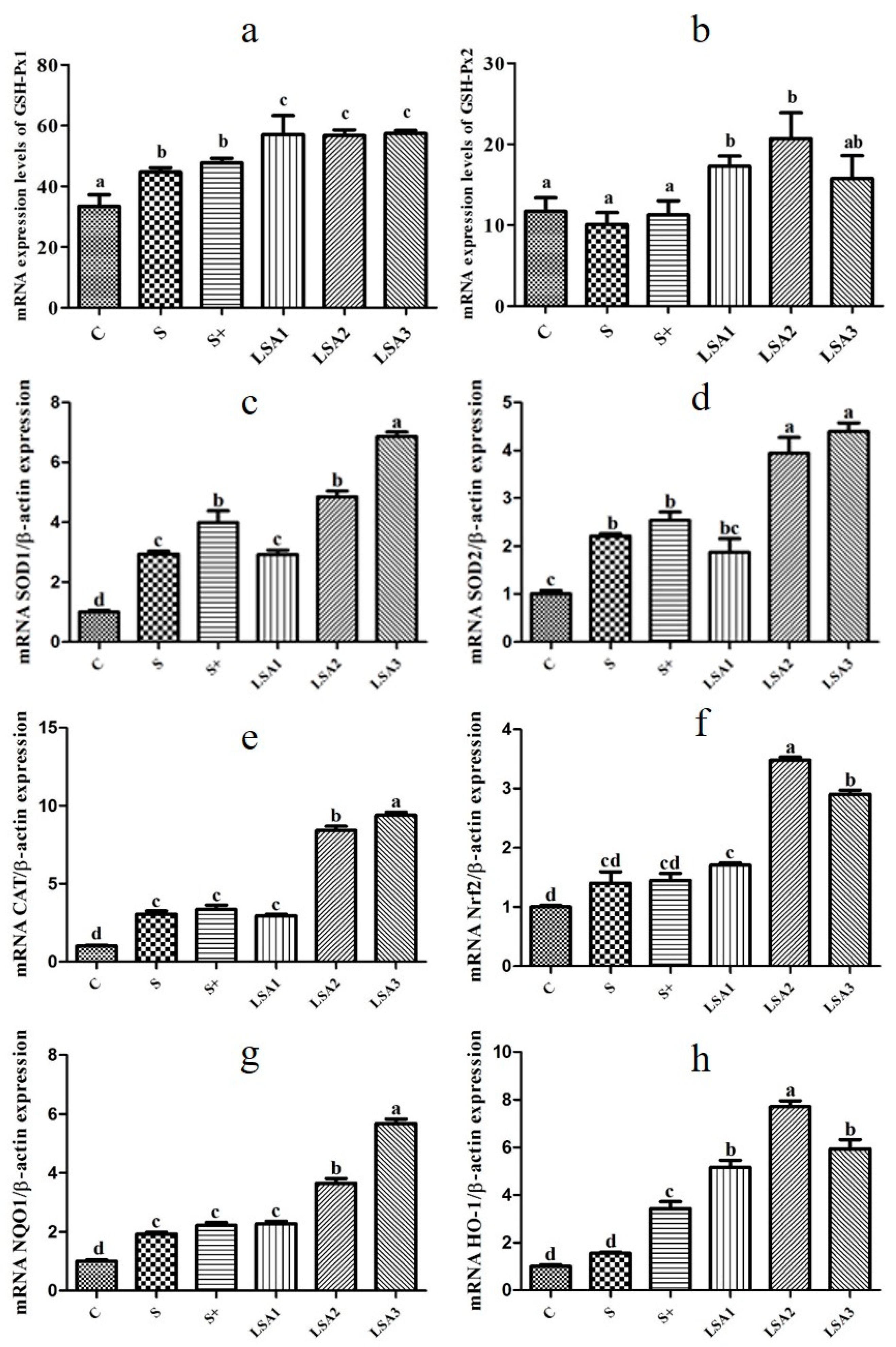
2.7. Effects of LSA on the Expression of Antioxidant Genes in the Ileum
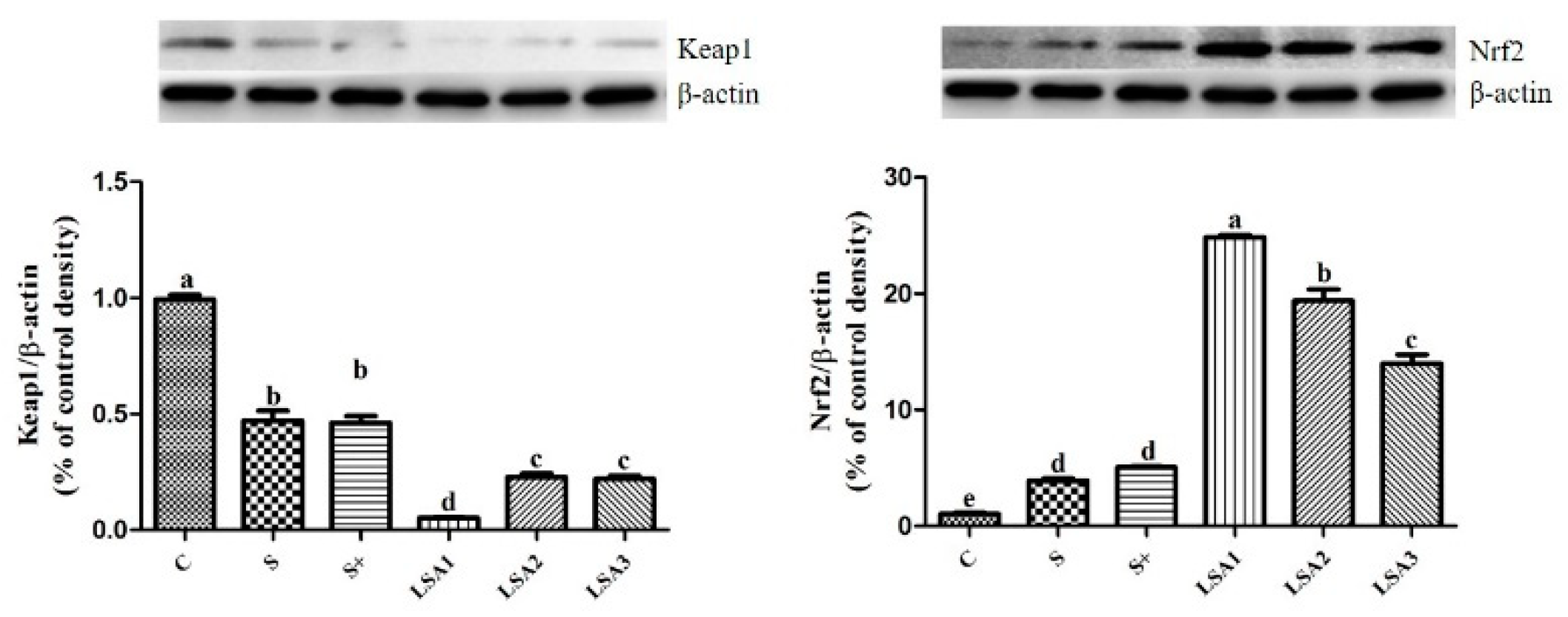
2.8. Effects of LSA on Expression Level of Kelch-Like ECH-Associated Protein 1 (Keap1) and Nrf2
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Animals
4.3. Sample Collection
4.4. Determination of Growth Performance and Diarrhea Rate
4.5. Histopathological Observation
4.6. Determination of DAO and d-LA in Serum
4.7. Determination of Antioxidant Indices in Serum
4.8. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR)
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McLamb, B.L.; Gibson, A.J.; Overman, E.L.; Stahl, C.; Moeser, A.J. Early weaning stress in pigs impairs innate mucosal immune responses to enterotoxigenic E. coli challenge and exacerbates intestinal injury and clinical disease. PLoS ONE 2013, 8, e59838. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.L.; Blikslager, A.T. Impaired intestinal barrier function and relapsing digestive disease: Lessons from a porcine model of early life stress. Neurogastroenterol. Motil. 2017, 29, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.; Gonzalez, L.; Blikslager, A. Large animal models: The key to translational discovery in digestive disease research. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.M.; Moeser, A.J.; Blikslager, A.T. Porcine models of digestive disease: The future of large animal translational research. Transl. Res. 2015, 166, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.S.; Ma, L.; Xiang, X.W.; Tang, Z.; Guan, R.F.; Qu, Y.L. Protective effect of low molecular-weight seleno-aminopolysaccharides against H2O2-induecd oxidative stress in intestinal epithelial cells. Int. J. Biol. Macromol. 2018, 112, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Jia, Z.; Misra, H.; Li, Y.R. Oxidative stress and redox signaling mechanisms of alcoholic liver disease: Updated experimental and clinical evidence. J. Dig. Dis. 2012, 13, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Ruane, D.T.; Lavelle, E.C. The role of CD103+ dendritic cells in the intestinal mucosal immune system. Front. Immunol. 2011, 2, 25. [Google Scholar] [CrossRef]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of oxidative stress in inflammatory bowel disease and potential antioxidant therapies. Oxid. Med. Cell. Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Joh, T. Oxidative stress and ischemia-reperfusion injury in gastrointestinal tract and antioxidant, protective agents. J. Clin. Biochem. Nutr. 2007, 40, 1–12. [Google Scholar] [CrossRef]
- Barrachina, M.D.; Panés, J.; Esplugues, J.V. Role of nitric oxide in gastrointestinal inflammatory and ulcerative diseases: Perspective for drugs development. Curr. Pharm. Des. 2001, 7, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Zhou, Z.; Chen, C.; Li, M.; Pei, L.; Chu, F.; Yang, J.; Wang, Y.; Li, L.; Liu, C.; et al. Lycopene protects against trimethyltin-induced neurotoxicity in primary cultured rat hippocampal neurons by inhibiting the mitochondrial apoptotic pathway. Neurochem. Int. 2011, 59, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Grisham, M.B. Oxidants and free radicals in inflammatory bowel disease. Lancet 1994, 344, 859–861. [Google Scholar] [CrossRef]
- Gao, Y.; Yuan, Y.; Zhou, X.; Wang, F.; Zhang, J. Preparation and characterization of polysaccharide selenate of tamarind seeds. Chin. J. Pharm. Anal. 2012, 32, 1222–1226. [Google Scholar]
- Tie, M.; Li, C.; Fei, J.; Zhao, S.; Shi, X.; Li, H.; Zang, S. Study on the Purification and IR spectroscopy of Se-polysaccharides in selenium-enriched flammulina velutipes. J. Instr. Anal. 2008, 27, 158–161. [Google Scholar]
- Zaharoff, D.A.; Rogers, C.J.; Hance, K.W.; Schlom, J.; Greiner, J.W. Chitosan solution enhances both humoral and cell-mediated immune responses to subcutaneous vaccination. Vaccine 2007, 25, 2085–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.X.; Wen, Z.S.; Xiang, X.W.; Ma, L.; Wang, X.B.; Ma, J.Y.; Qu, Y.L. Immunomodulatory effect of low molecular-weight seleno-aminopolysaccharides in intestinal epithelial cells. Int. J. Biol. Macromol. 2017, 99, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Wen, Z.S.; Zhang, P.; Qu, Y.L.; Xu, Z.R. Acute toxicity of low molecular seleno-aminopolysaccharide to mice. Chin. J. Vet. Sci. 2017, 37, 1790–1796. [Google Scholar]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Macpherson A J Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Ridnour, L.A.; Isenberg, J.S.; Espey, M.G.; Thomas, D.D.; Roberts, D.D.; Wink, D.A. Nitric oxide regulates angiogenesis through a functional switch involving thrombospondin-1. Proc. Natl. Acad. Sci. USA 2005, 102, 13147–13152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Morris, H.; Mazúr, M.; Rapta, P.; Bilton, R.F. Oxygen free radical generating mechanisms in the colon: Do the semiquinones of vitamin K play a role in the aetiology of colon cancer? Biochim. Biophys. Acta. 2001, 1527, 161–166. [Google Scholar] [CrossRef]
- Inokuma, T.; Haraguchi, M.; Fujita, F.; Tajima, Y.; Kanematsu, T. Oxidative stress and tumor progression in colorectal cancer. Hepatogastroenterology 2009, 56, 343–347. [Google Scholar] [PubMed]
- Liang, X.L.; Dang, W.Q.; Yang, J.Q.; Min, H.E.; Zhang, J. Effects of yeast selenium on production performance and diarrhea incidence of weaned piglets. Anim. Husb. Feed Sci. 2017, 38, 37–39. [Google Scholar]
- Salles, M.S.V.; Zanetti, M.A.; Junior, L.C.R.; Salles, F.A.; Azzolini, A.E.C.S.; Soares, E.M.; Faccioli, L.H.; Valim, Y.M.L. Performance and immune response of suckling calves fed organic selenium. Anim. Feed Sci. Technol. 2014, 188, 28–35. [Google Scholar] [CrossRef]
- Payne, R.L. The Effects of Inorganic and Organic Selenium Sources on Growth Performance, Carcass Traits, Tissue Mineral Concentrations, and Enzyme Activity in Poultry. Ph.D. Thesis, Louisiana State University, Baton Rouge, LA, USA, August 2004. [Google Scholar]
- Segovia, J.; Arzola, C.; Ruiz, O.; Salinas-Chavira, J.; Rodriguez-Muela, C.; Jimenez, J.; González-Garcia, H.; Castillo-Castillo, Y. Effect of organic and inorganic selenium supplementation on weight performance of ewes and lambs. J. Anim. Vet. Adv. 2012, 7, 1555–1558. [Google Scholar]
- Yang, Y.R.; Meng, F.C.; Wang, P.; Jiang, Y.B.; Yin, Q.Q.; Chang, J.; Zuo, R.Y.; Zheng, Q.H.; Liu, J.X. Effect of organic and inorganic selenium supplementation on growth performance, meat quality and antioxidant property of broilers. Afr. J. Biotechnol. 2012, 11, 3031–3036. [Google Scholar]
- Catalioto, R.M.A.; Maggi, C.; Giuliani, S. Intestinal Epithelial Barrier Dysfunction in Disease and Possible Therapeutical Interventions. Curr. Med. Chem. 2011, 18, 398–426. [Google Scholar] [CrossRef] [PubMed]
- Salvo, R.E.; Alonso, C.C.; Pardo, C.C.; Casado, B.M.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef]
- Martinez, C.; Gonzalez-Castro, A.; Vicario, M.; Santos, J. Cellular and molecular basis of intestinal barrier dysfunction in the irritable bowel syndrome. Gut Liver 2012, 6, 305–315. [Google Scholar] [CrossRef]
- Van Itallie, C.M.; Anderson, J.M. Architecture of tight junctions and principles of molecular composition. Semin. Cell Dev. Biol. 2014, 36, 157–165. [Google Scholar] [CrossRef]
- Smith, F.; Clark, J.E.; Overman, B.L. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moeser, A.J.; Borst, L.B.; Overman, B.L. Defects in small intestinal epithelial barrier function and morphology associated with peri-weaning failure to thrive syndrome (PFTS) in swine. Res. Vet. Sci. 2012, 93, 975–982. [Google Scholar] [CrossRef]
- Castillo, M.; Martín-Orúe, S.M.; Nofrarías, M. Changes in caecal microbiota and mucosal morphology of weaned pigs. Vet. Microbiol. 2007, 124, 239–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Péron, V.; Le Huërou-Luron, I.; Lallès, J.P.; Sève, B. Weaning induces both transient and long-lasting modifications of absorptive, secretory, and barrier properties of piglet intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.H.; Zhao, K.L.; Chen, X.L.; Xu, J.X. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2012, 90, 2581–2589. [Google Scholar] [CrossRef] [PubMed]
- Dalgaard, T.S.; Briens, M.; Engberg, R.M.; Lauridsen, C. The influence of selenium and selenoproteins on immune responses of poultry and pigs. Anim. Feed Sci. Technol. 2018, 238, 73–83. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Habeos, I.G.; Samuelson, A.V.; Bohmann, D. The role of the antioxidant and longevity-promoting Nrf2 pathway in metabolic regulation. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Motohashi, H.; Yamamoto, M. Toward clinical application of the Keap1-Nrf2 pathway. Trends Pharmacol. Sci. 2013, 34, 340–346. [Google Scholar] [CrossRef]
- Vachharajani, T.J.; Work, J.; Issekutz, A.C.; Granger, D.N. Heme oxygenase modulates selectin expression in different regional vascular beds. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H1613–H1617. [Google Scholar] [CrossRef]
- Kwak, M.K.; Wakabayashi, N.; Kensler, T.W. Chemoprevention through the Keap1-Nrf2 signaling pathway by phase 2 enzyme inducers. Mutat. Res-Fundam. Mol. 2004, M555, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Reszka, E.; Wieczorek, E.; Jablonska, E.; Janasik, B.; Fendler, W.; Wasowicz, W. Association between plasma selenium level and Nrf2 target genes expression in humans. J. Trace Elem. Med. Biol. 2014, 30, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Fontagnédicharry, S.; Godin, S.; Liu, H.; Antony, J.P.P.; Bouyssière, B.; Bueno, M.; Tacon, P.; Médale, F.; Kaushik, S.J. Influence of the forms and levels of dietary selenium on antioxidant status and oxidative stress-related parameters in rainbow trout (Oncorhynchus mykiss) fry. Br. J. Nutr. 2015, 113, 1876–1887. [Google Scholar] [CrossRef]
- Lian, F.; Wang, X.D. Enzymatic metabolites of lycopene induce Nrf2-mediated expression of phase II detoxifying/antioxidant enzymes in human bronchial epithelial cells. Int. J. Cancer 2008, 123, 1262–1268. [Google Scholar] [CrossRef] [PubMed]
- Kraft, A.D.; Johnson, D.A.; Johnson, J.A. Nuclear factor E2-related factor 2-dependent antioxidant response element activation by tert-butylhydroquinone and sulforaphane occurring preferentially in astrocytes conditions neurons against oxidative insult. J. Neurosci. 2004, 24, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Li, G.; Wang, B. Study on the physical and chemical properties and the molecular structure of seleno-chitosan. Chin. J. Mar. Drugs 2000, 54, 124–126. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | C | S | S+ | LSA1 | LSA2 | LSA3 |
|---|---|---|---|---|---|---|
| Initial weight (kg) | 8.28 ± 0.09 | 8.28 ± 0.12 | 8.35 ± 0.17 | 8.33 ± 0.12 | 8.30 ± 0.15 | 8.31 ± 0.13 |
| Final weight (kg) | 19.36 ± 0.87 b | 19.50 ± 0.81 b | 19.49 ± 0.93 b | 20.29 ± 0.90 a | 20.42 ± 1.07 a | 19.35 ± 0.90 b |
| Average daily gain (g) | 395.89 ± 31.3 b | 400.70 ± 30.10 b | 398.07 ± 35.59 b | 427.03 ± 31.06 a | 432.90 ± 37.51 a | 394.32 ± 33.91 b |
| Average daily feed intake (g) | 662.15 ± 10.28 | 648.57 ± 8.84 | 651.31 ± 16.87 | 650.36 ± 5.71 | 666.79 ± 8.89 | 646.55 ± 8.47 |
| Feed/gain ratio | 1.67 ± 0.05 a | 1.62 ± 0.04 a | 1.64 ± 0.05 a | 1.52 ± 0.03 b | 1.54 ± 0.03 b | 1.64 ± 0.04 a |
| Groups | Feed Composition |
|---|---|
| Control (C) | Basal diet |
| Na2SeO3 (S) | Basal diet + 0.3 mg/kg Se (Na2SeO3) |
| Na2SeO3 + LA (S+) | Basal diet + 0.3 mg/kg Se (Na2SeO3) + 11 mg/kg LA |
| LSA1 | Basal diet + 17.15 mg/kg LSA (0.3 mg/kg Se) |
| LSA2 | Basal diet + 34.30 mg/kg LSA (0.6 mg/kg Se) |
| LSA3 | Basal diet + 51.45 mg/kg LSA (0.9 mg/kg Se) |
| Degree of Diarrhea | Fecal Appearance | Water Volume (%) | Score |
|---|---|---|---|
| No diarrhea | Normal, strip, or long strip | <70 | 1 |
| Mild diarrhea | Soft, basically shaped | 70–75 | 2 |
| Moderate diarrhea | Viscous, not formed, no separation of manure | 75–80 | 3 |
| Severe diarrhea | Liquid, not formed, separation of manure | >80 | 4 |
| Gene | Gene Accession Number | Primer Sequence 5′-3′ | PCR Product Size (bp) | Tm |
|---|---|---|---|---|
| SOD1 | NM_000454 | F: TCCATGTCCATCAGTTTGGA R: CTGCCCAAGTCATCTGGTTT | 250 | 50 |
| SOD2 | NM_001322820 | F: TGGAGGCCACATCAATCATA R: AGCGGTCAACTTCTCCTTGA | 136 | 59 |
| CAT | NM_214301.2 | F: TGTGAACTGTCCCTTCCGTG R: CGTCTGTTCGGGAGCACTAA | 124 | 60 |
| GPx1 | NM_201397.2 | F: TGGGGAGATCCTGAATTG R: GATAAACTTGGGGTCGGT | 184 | 53 |
| GPx2 | NM_001115136.1 | F: TCGTGGCTTCCCTTGCAAC R: CCATTCACGTCACACTTCTG | 138 | 61 |
| Nrf2 | XM_021075133.1 | F: CACCACCTCAGGGTAATA R: GCGGCTTGAATGTTTGTC | 125 | 55 |
| HO-1 | NM_001004027.1 | F: AGCTGTTTCTGAGCCTCCAA R: CAAGACGGAAACACGAGACA | 130 | 59.2 |
| NQO1 | NM_001159613.1 | F: CCAGCAGCCCGGCCAATCTG R: AGGTCCGACACGGCGACCTC | 160 | 66.2 |
| ZO-1 | NM_001355015 | F: CAGCCCGAGGCGTGTTTA R: TGGGAGGATGCTGTTGTCT | 146 | 60 |
| Occludin | NM_214301.2 | F: GCTGGAGGAAGACTGGAT R: ATCCGCAGATCCCTTAAC | 124 | 61 |
| β-actin | XM_003124280 | F: GATGAGATTGGCATGGCTTT R: CACCTTCACCGTTCCAGTTT | 122 | 60 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Z.-S.; Tang, Z.; Ma, L.; Zhu, T.-L.; Wang, Y.-M.; Xiang, X.-W.; Zheng, B. Protective Effect of Low Molecular Weight Seleno-Aminopolysaccharide on the Intestinal Mucosal Oxidative Damage. Mar. Drugs 2019, 17, 64. https://doi.org/10.3390/md17010064
Wen Z-S, Tang Z, Ma L, Zhu T-L, Wang Y-M, Xiang X-W, Zheng B. Protective Effect of Low Molecular Weight Seleno-Aminopolysaccharide on the Intestinal Mucosal Oxidative Damage. Marine Drugs. 2019; 17(1):64. https://doi.org/10.3390/md17010064
Chicago/Turabian StyleWen, Zheng-Shun, Zhen Tang, Li Ma, Tian-Long Zhu, You-Ming Wang, Xing-Wei Xiang, and Bin Zheng. 2019. "Protective Effect of Low Molecular Weight Seleno-Aminopolysaccharide on the Intestinal Mucosal Oxidative Damage" Marine Drugs 17, no. 1: 64. https://doi.org/10.3390/md17010064