
Astaxanthin Attenuates Early Acute Kidney Injury Following Severe Burns in Rats by Ameliorating Oxidative Stress and Mitochondrial-Related Apoptosis

Abstract
:1. Introduction
2. Results
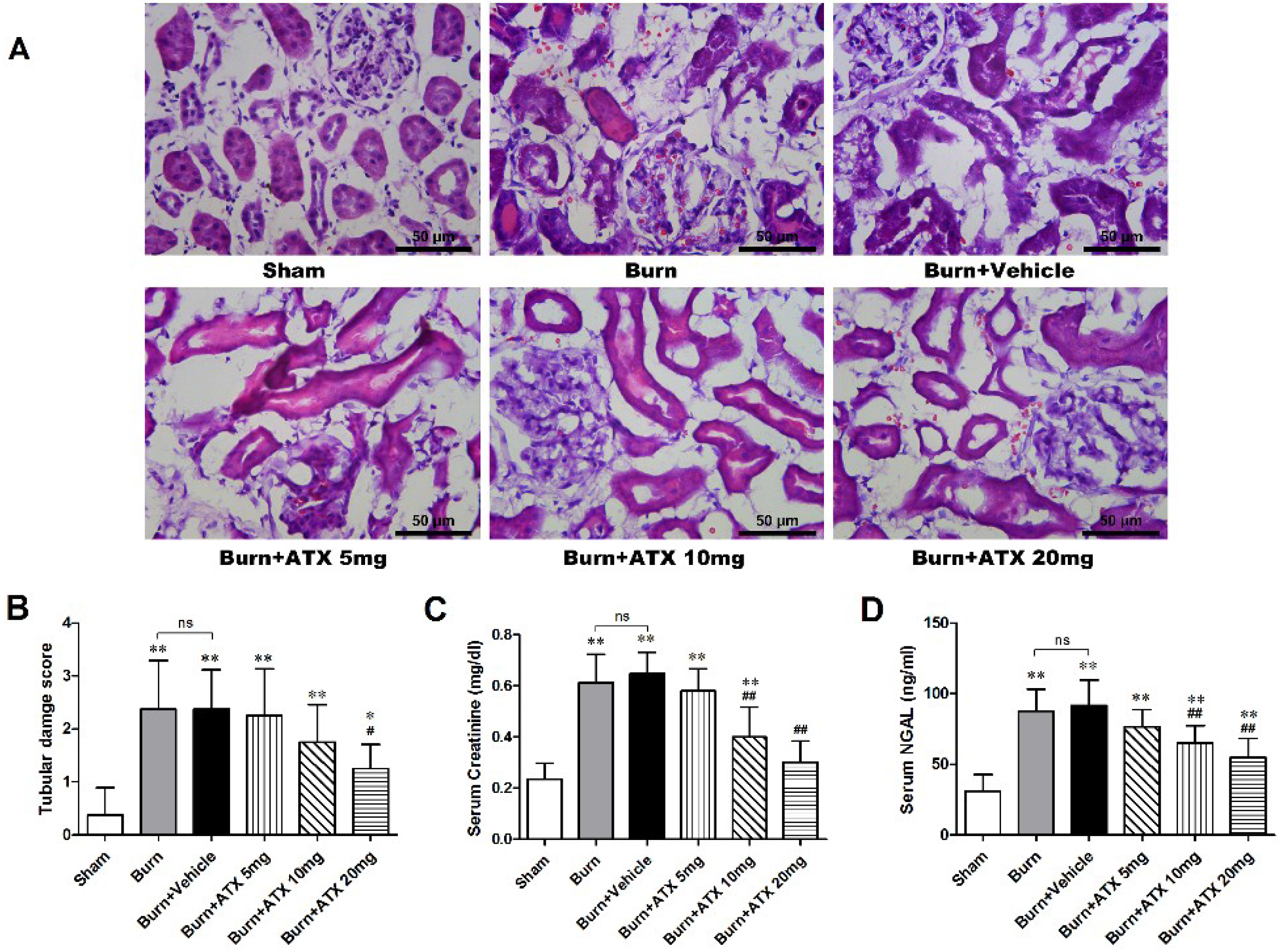
2.1. ATX Attenuates the Histological and Functional Deterioration of Severely Burned rat Kidneys

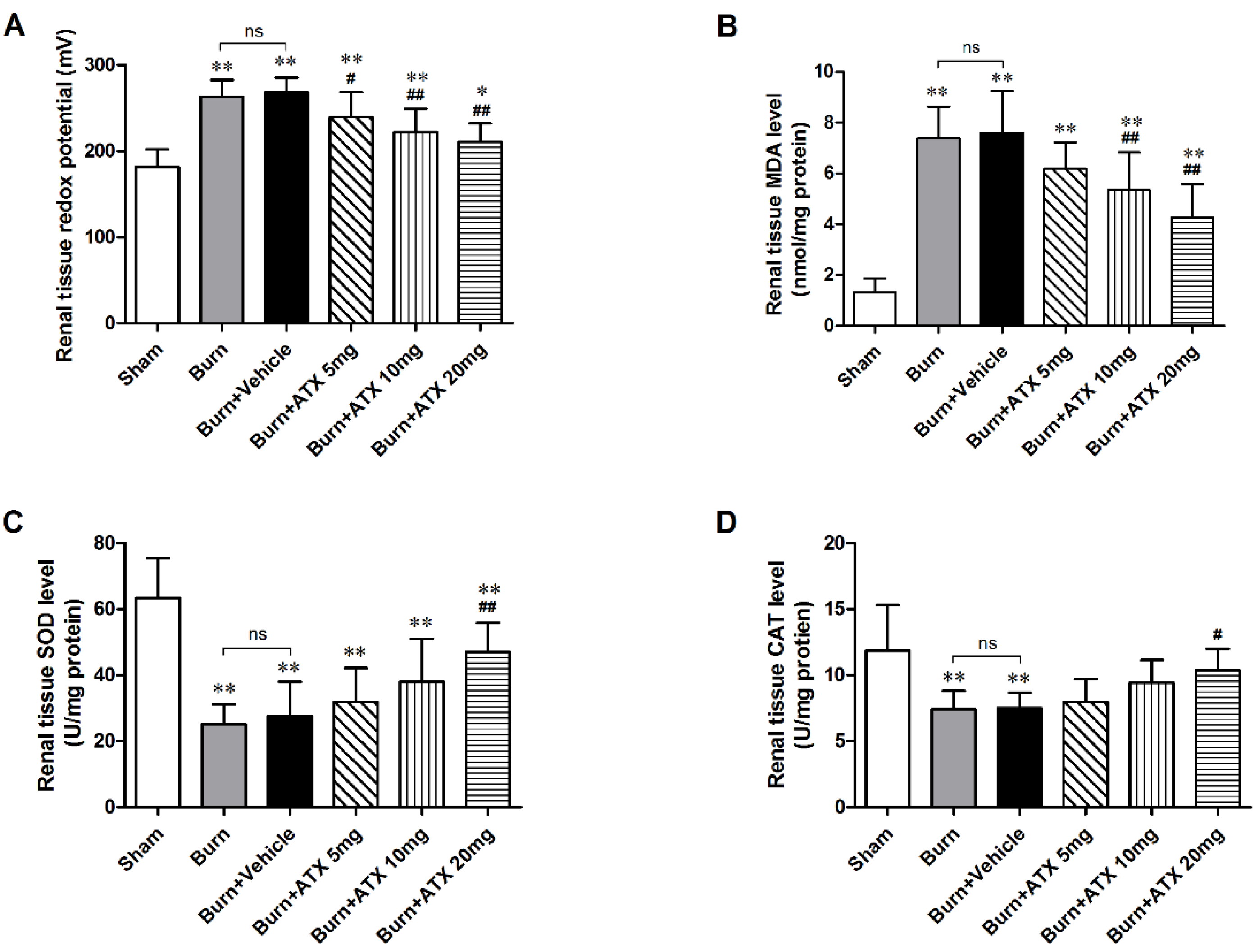
2.2. ATX Ameliorates Severe Burn-Induced Oxidative Stress in Rat Renal Tissue

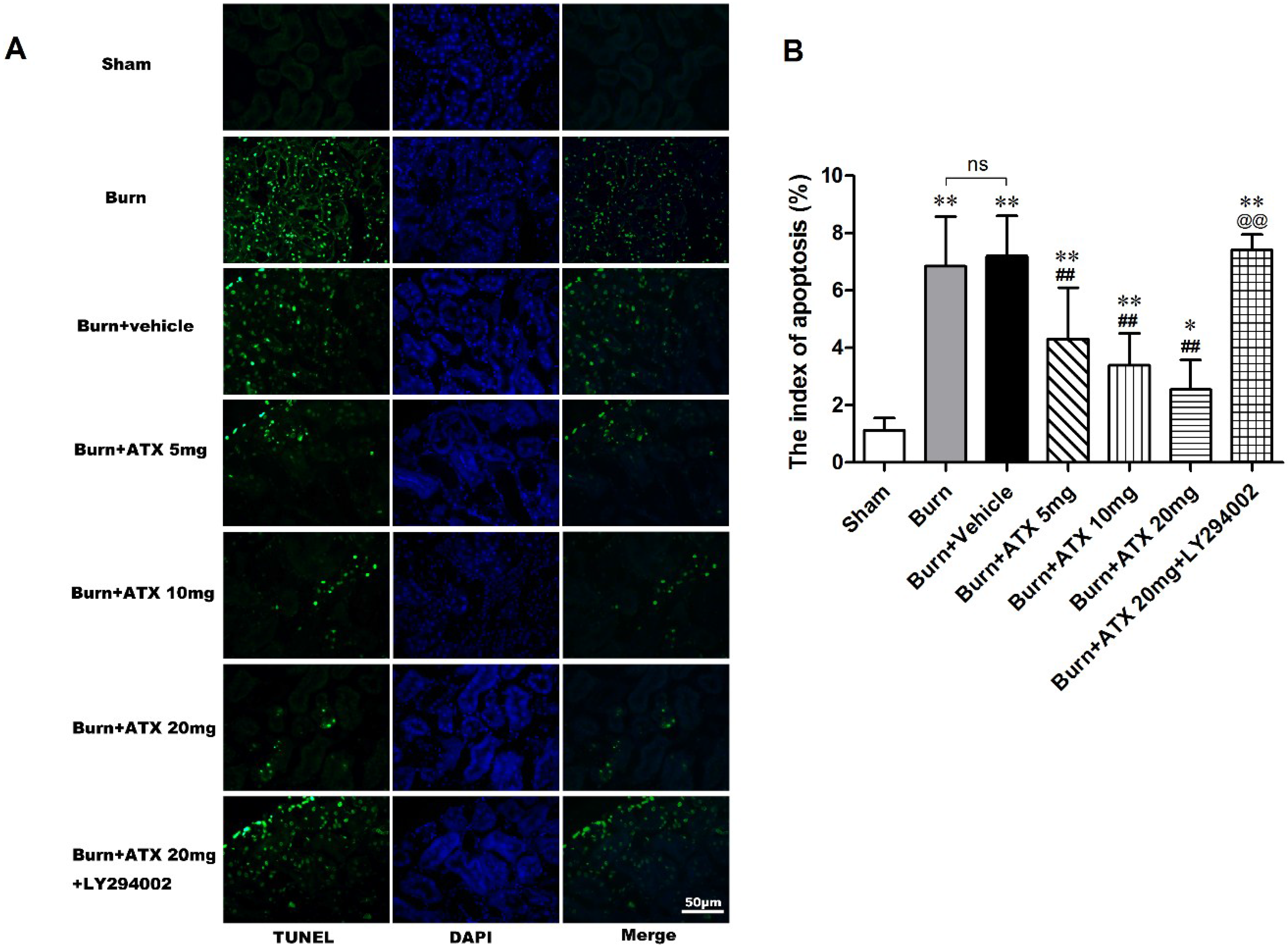
2.3. ATX Relieves Burn-Induced Tubular Apoptosis in Rats

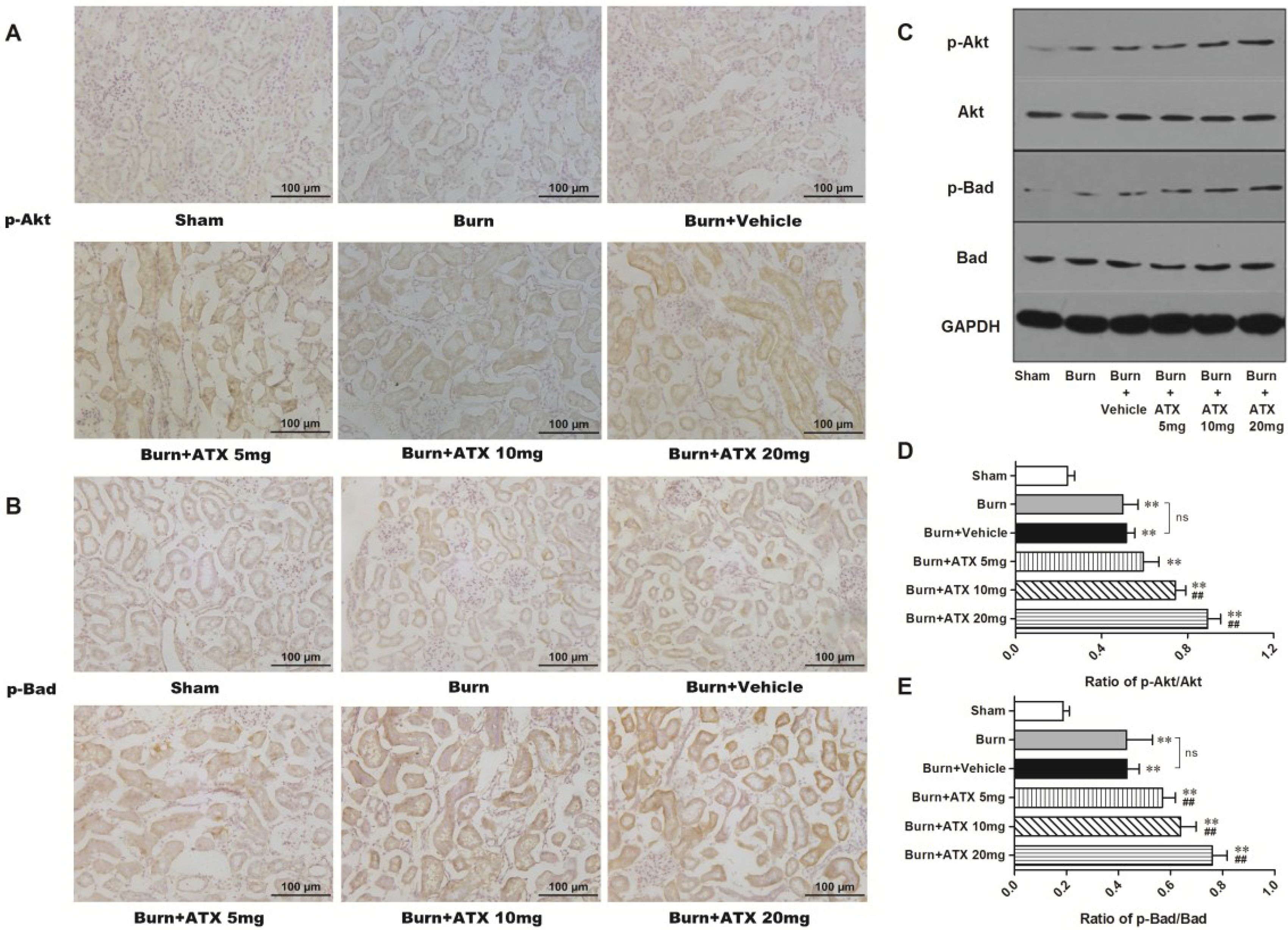
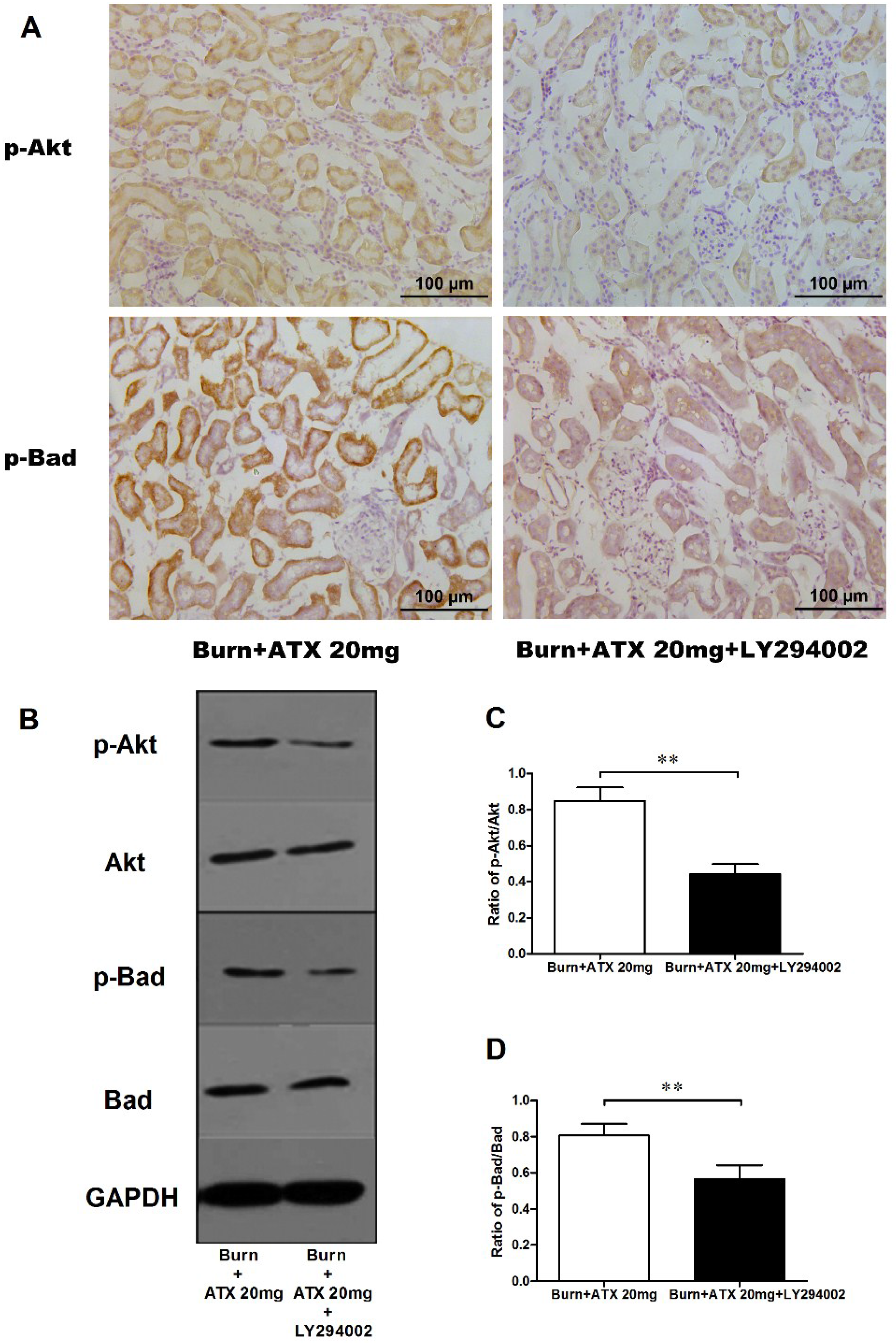
2.4. ATX Dose-Dependently Increases the Distribution and Protein Expression of p-Akt and p-Bad in Rats with Burns

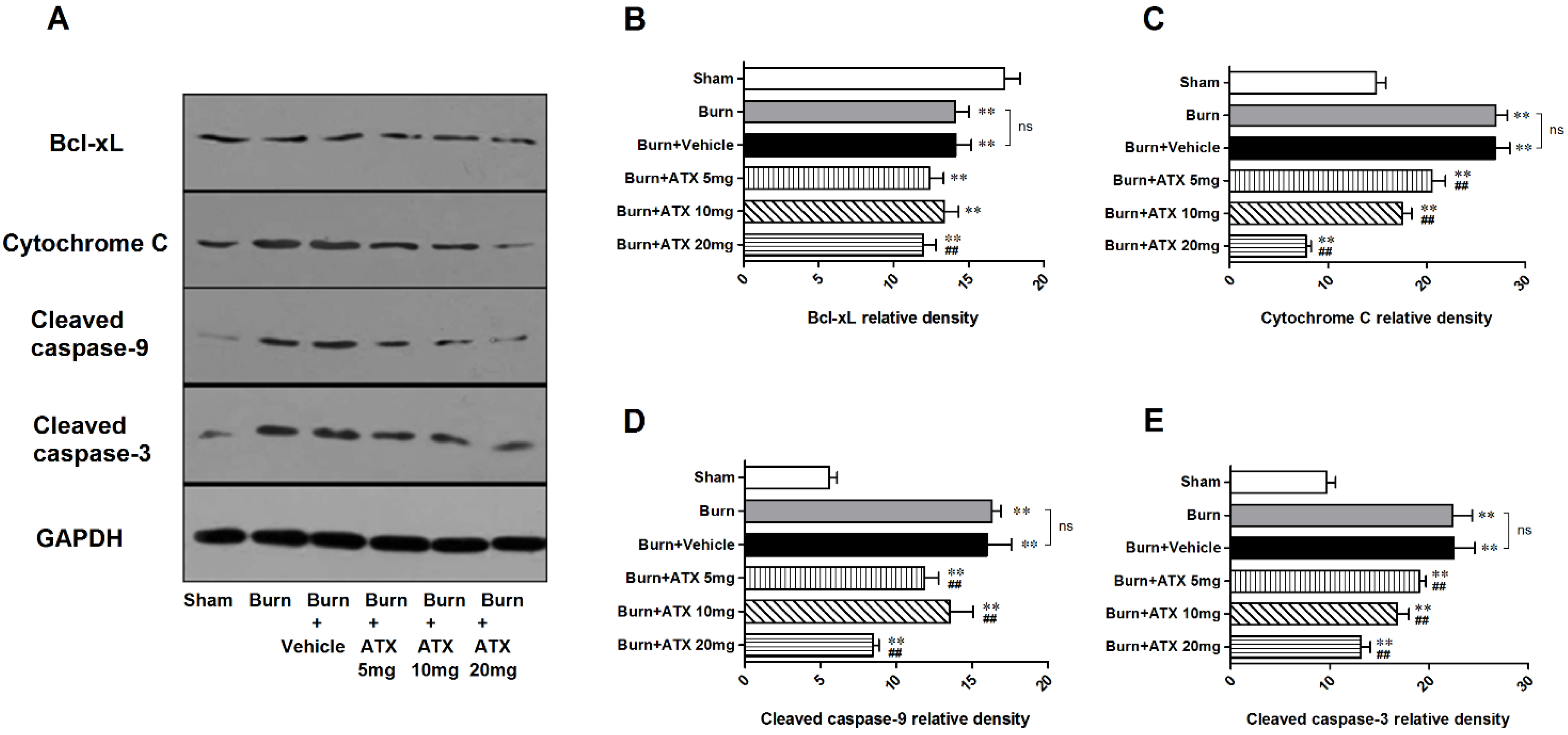
2.5. Effects of ATX on Pro-Apoptotic Proteins of Mitochondrial Pathway

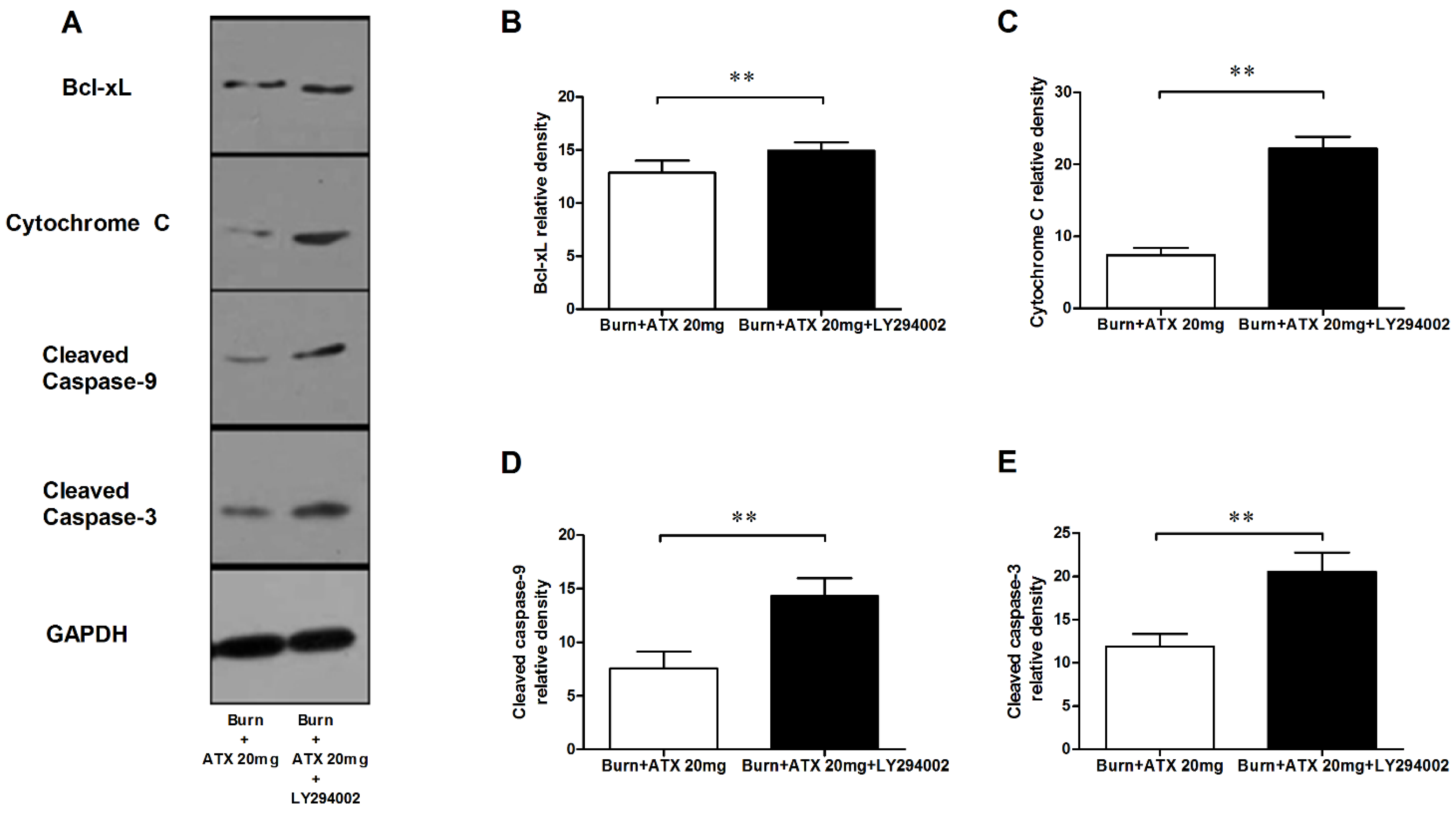
2.6. LY294002 Reverses the Effects of ATX on Renal Mitochondria Related Apoptosis after Burn


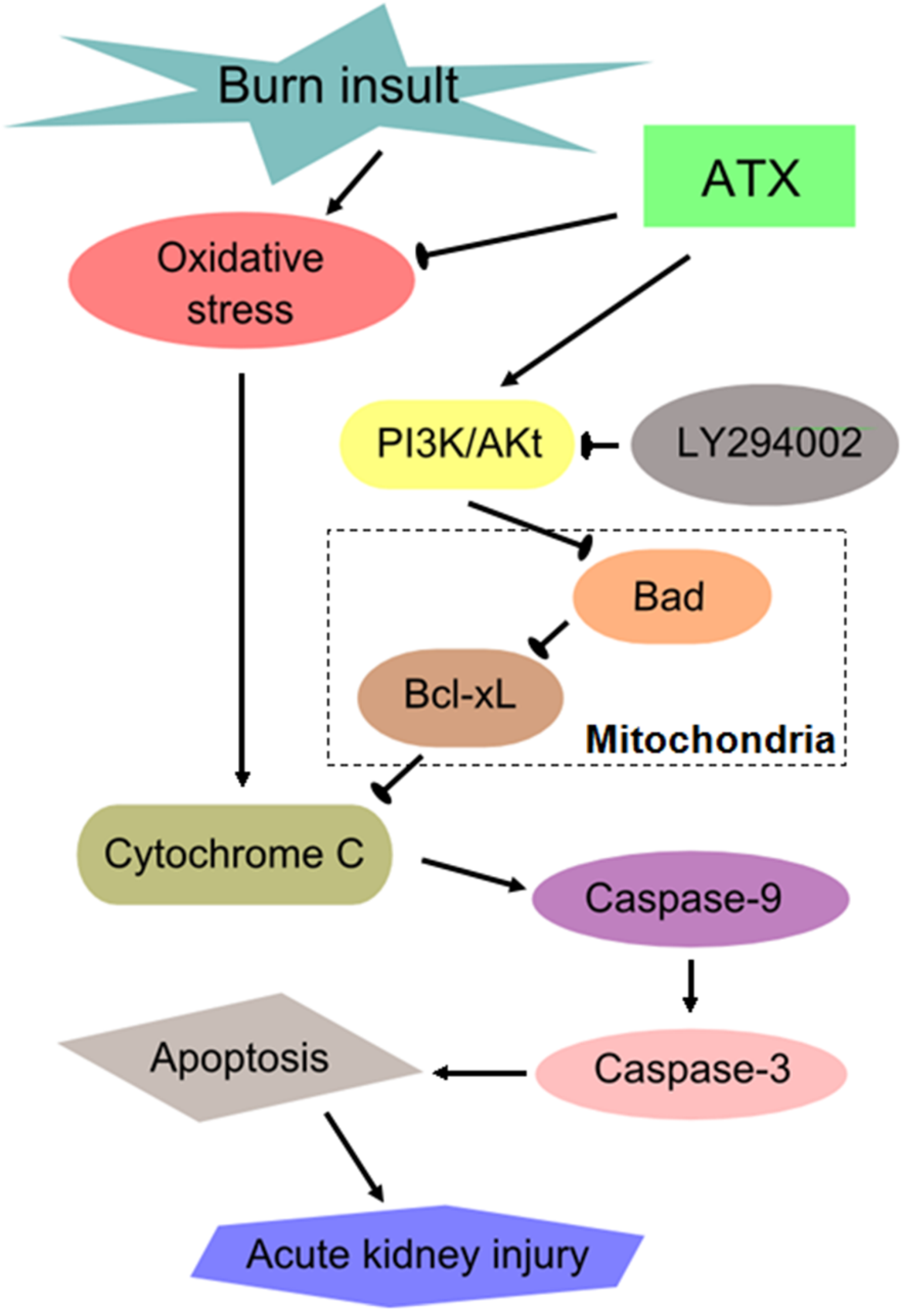
3. Discussion

4. Materials and Methods
4.1. Animals
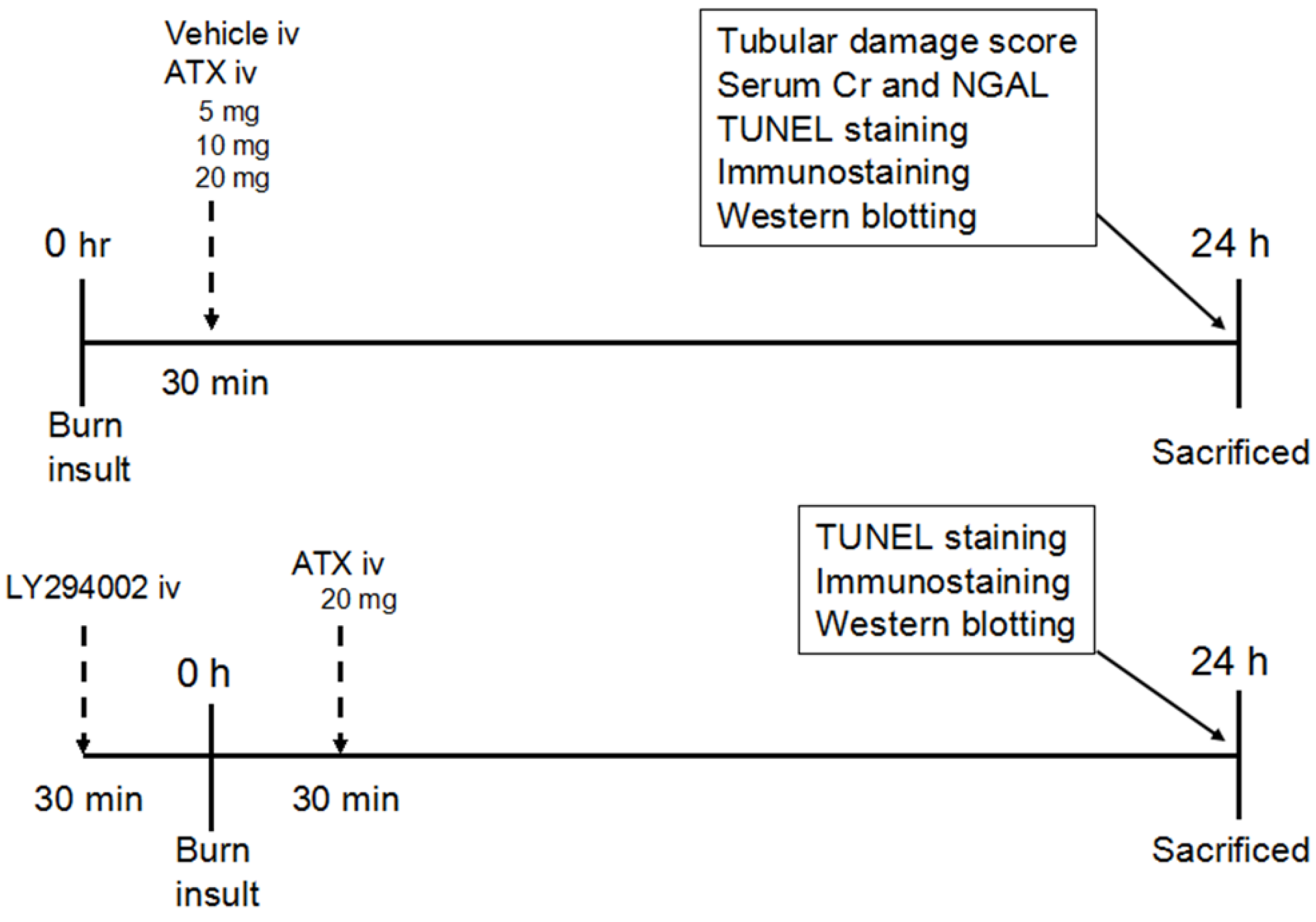
4.2. Severe Burn Model and Experiment Groups

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assessment Score | 0 | 1 | 2 | 3 |
|---|---|---|---|---|
| Attitude and posture | Alert and not hunched | Not alert or hunched | Not alert and hunched | Not responsive to stimuli |
| Gait and movement | Active | Somewhat inactive | Completely inactive | Lying on side |
| Burn wound site | Clean and not swollen | Exudates or swelling | Exudates and swelling | Increased wound depth in examination |
| Appetite | Eating and drinking | Reduced eating or drinking | Not eating or drinking | Not eating or drinking for >3 days |
| Elimination | Normal | Softer than normal | Diarrhea | Diarrhea >3 days |

4.3. Histological and Renal Function Evaluation
4.4. Assessment of Oxidative Stress
4.5. TUNEL Staining for Apoptosis
4.6. Immunohistochemistry (IHC) Staining
4.7. Western Blotting Analysis
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mosier, M.J.; Pham, T.N.; Klein, M.B.; Gibran, N.S.; Arnoldo, B.D.; Gamelli, R.L.; Tompkins, R.G.; Herndon, D.N. Early acute kidney injury predicts progressive renal dysfunction and higher mortality in severely burned adults. J. Burn Care Res. 2010, 31, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, T.; Lavrentieva, A.; Greenhalgh, D.G. Acute kidney injury in critically ill burn patients. Risk factors, progression and impact on mortality. Burns 2010, 36, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Steinvall, I.; Bak, Z.; Sjoberg, F. Acute kidney injury is common, parallels organ dysfunction or failure, and carries appreciable mortality in patients with major burns: A prospective exploratory cohort study. Crit. Care 2008, 12, R124. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, M.; Thibeault, Y.; Querin, S. Continuous haemofiltration and haemodiafiltration for acute renal failure in severely burned patients. Burns 1997, 23, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Coca, S.G.; Bauling, P.; Schifftner, T.; Howard, C.S.; Teitelbaum, I.; Parikh, C.R. Contribution of acute kidney injury toward morbidity and mortality in burns: A contemporary analysis. Am. J. Kidney Dis. 2007, 49, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Colpaert, K.; Hoste, E.A. Acute kidney injury in burns: A story of volume and inflammation. Crit. Care 2008, 12, 192. [Google Scholar] [CrossRef] [PubMed]
- Mustonen, K.M.; Vuola, J. Acute renal failure in intensive care burn patients (arf in burn patients). J. Burn Care Res. 2008, 29, 227–237. [Google Scholar] [PubMed]
- Yavuz, S.; Anarat, A.; Acarturk, S.; Dalay, A.C.; Kesiktas, E.; Yavuz, M.; Acarturk, T.O. Neutrophil gelatinase associated lipocalin as an indicator of acute kidney injury and inflammation in burned children. Burns 2014, 40, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Hoetzenecker, W.; Echtenacher, B.; Guenova, E.; Hoetzenecker, K.; Woelbing, F.; Bruck, J.; Teske, A.; Valtcheva, N.; Fuchs, K.; Kneilling, M.; et al. Ros-induced atf3 causes susceptibility to secondary infections during sepsis-associated immunosuppression. Nat. Med. 2012, 18, 128–134. [Google Scholar] [CrossRef]
- Parihar, A.; Parihar, M.S.; Milner, S.; Bhat, S. Oxidative stress and anti-oxidative mobilization in burn injury. Burns 2008, 34, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Sedeek, M.; Callera, G.; Montezano, A.; Gutsol, A.; Heitz, F.; Szyndralewiez, C.; Page, P.; Kennedy, C.R.; Burns, K.D.; Touyz, R.M.; et al. Critical role of nox4-based nadph oxidase in glucose-induced oxidative stress in the kidney: Implications in type 2 diabetic nephropathy. Am. J. Physiol. Ren. Physiol. 2010, 299, F1348–F1358. [Google Scholar] [CrossRef]
- Zhang, X.S.; Zhang, X.; Zhou, M.L.; Zhou, X.M.; Li, N.; Li, W.; Cong, Z.X.; Sun, Q.; Zhuang, Z.; Wang, C.X.; et al. Amelioration of oxidative stress and protection against early brain injury by astaxanthin after experimental subarachnoid hemorrhage. J. Neurosurg. 2014, 121, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.W. Free radicals and lipid peroxidation mediated injury in burn trauma: The role of antioxidant therapy. Toxicology 2003, 189, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Gravante, G.; Delogu, D.; Sconocchia, G. “Systemic apoptotic response” after thermal burns. Apoptosis 2007, 12, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Quoilin, C.; Mouithys-Mickalad, A.; Lecart, S.; Fontaine-Aupart, M.P.; Hoebeke, M. Evidence of oxidative stress and mitochondrial respiratory chain dysfunction in an in vitro model of sepsis-induced kidney injury. Biochim. Biophys. Acta 2014, 1837, 1790–1800. [Google Scholar] [CrossRef] [PubMed]
- Sahu, B.D.; Rentam, K.K.; Putcha, U.K.; Kuncha, M.; Vegi, G.M.; Sistla, R. Carnosic acid attenuates renal injury in an experimental model of rat cisplatin-induced nephrotoxicity. Food Chem. Toxicol. 2011, 49, 3090–3097. [Google Scholar] [CrossRef] [PubMed]
- Wagener, F.A.; Dekker, D.; Berden, J.H.; Scharstuhl, A.; van der Vlag, J. The role of reactive oxygen species in apoptosis of the diabetic kidney. Apoptosis 2009, 14, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Liu, Y.; Wang, L.; Cai, X.; Wang, D.; Wu, K.; Chen, H.; Li, J.; Lei, W. Sustained oxidative stress causes late acute renal failure via duplex regulation on p38 mapk and akt phosphorylation in severely burned rats. PLoS ONE 2013, 8, e54593. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Luo, M.; Liu, D.; Chen, B.; Zhang, W.; Mai, L.; Zeng, J.; Huang, N.; Huang, Y.; Mo, X.; et al. Bad overexpression inhibits cell growth and induces apoptosis via mitochondrial-dependent pathway in non-small cell lung cancer. Cancer Cell Int. 2013, 13, 53. [Google Scholar] [CrossRef]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of bad couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Stoyanoff, T.R.; Todaro, J.S.; Aguirre, M.V.; Zimmermann, M.C.; Brandan, N.C. Amelioration of lipopolysaccharide-induced acute kidney injury by erythropoietin: Involvement of mitochondria-regulated apoptosis. Toxicology 2014, 318, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zeng, M.; Chen, W.; Liu, C.; Wang, F.; Han, X.; Zuo, Z.; Peng, S. Dexmedetomidine reduces isoflurane-induced neuroapoptosis partly by preserving pi3k/akt pathway in the hippocampus of neonatal rats. PLoS ONE 2014, 9, e93639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Zhang, X.; Wu, Q.; Li, W.; Zhang, Q.R.; Wang, C.X.; Zhou, X.M.; Li, H.; Shi, J.X.; Zhou, M.L. Astaxanthin alleviates early brain injury following subarachnoid hemorrhage in rats: Possible involvement of akt/bad signaling. Mar. Drugs 2014, 12, 4291–4310. [Google Scholar] [CrossRef] [PubMed]
- Noshita, N.; Lewen, A.; Sugawara, T.; Chan, P.H. Evidence of phosphorylation of akt and neuronal survival after transient focal cerebral ischemia in mice. J. Cereb. Blood Flow Metab. 2001, 21, 1442–1450. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.D.; Kang, H.E.; Yang, S.H.; Lee, M.G.; Shin, W.G. Pharmacokinetics and first-pass metabolism of astaxanthin in rats. Br. J. Nutr. 2011, 105, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Kuo, C.C.; Chou, J.; Delvolve, A.; Jackson, S.N.; Post, J.; Woods, A.S.; Hoffer, B.J.; Wang, Y.; Harvey, B.K. Astaxanthin reduces ischemic brain injury in adult rats. FASEB J. 2009, 23, 1958–1968. [Google Scholar] [CrossRef] [PubMed]
- Naguib, Y.M. Antioxidant activities of astaxanthin and related carotenoids. J. Agric. Food Chem. 2000, 48, 1150–1154. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, Y.J.; Kwon, K.H. Neuroprotective effects of astaxanthin in oxygen-glucose deprivation in sh-sy5y cells and global cerebral ischemia in rat. J. Clin. Biochem. Nutr. 2010, 47, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, Y.A.; Yokozawa, T. Protection against oxidative stress, inflammation, and apoptosis of high-glucose-exposed proximal tubular epithelial cells by astaxanthin. J. Agric. Food Chem. 2009, 57, 8793–8797. [Google Scholar] [CrossRef] [PubMed]
- Sila, A.; Ghlissi, Z.; Kamoun, Z.; Makni, M.; Nasri, M.; Bougatef, A.; Sahnoun, Z. Astaxanthin from shrimp by-products ameliorates nephropathy in diabetic rats. Eur. J. Nutr. 2015, 54, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Fu, K.; Zhao, X.; Zhang, Y.; Yuan, Y.; Zhang, S.; Gu, X.; Guo, H. Protective effects of astaxanthin against ischemia/reperfusion induced renal injury in mice. J. Transl. Med. 2015, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Gal, A.F.; Andrei, S.; Cernea, C.; Taulescu, M.; Catoi, C. Effects of astaxanthin supplementation on chemically induced tumorigenesis in wistar rats. Acta Vet. Scand. 2012, 54, 50. [Google Scholar] [CrossRef] [PubMed]
- Lauver, D.A.; Lockwood, S.F.; Lucchesi, B.R. Disodium disuccinate astaxanthin (cardax) attenuates complement activation and reduces myocardial injury following ischemia/reperfusion. J. Pharmacol. Exp. Ther. 2005, 314, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Nagendraprabhu, P.; Sudhandiran, G. Astaxanthin inhibits tumor invasion by decreasing extracellular matrix production and induces apoptosis in experimental rat colon carcinogenesis by modulating the expressions of erk-2, nfkb and cox-2. Investig. New Drugs 2011, 29, 207–224. [Google Scholar] [CrossRef]
- Arunkumar, E.; Bhuvaneswari, S.; Anuradha, C.V. An intervention study in obese mice with astaxanthin, a marine carotenoid—Effects on insulin signaling and pro-inflammatory cytokines. Food Funct. 2012, 3, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Fassett, R.G.; Coombes, J.S. Astaxanthin, oxidative stress, inflammation and cardiovascular disease. Future Cardiol. 2009, 5, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Sener, G.; Sehirli, A.O.; Gedik, N.; Dulger, G.A. Rosiglitazone, a ppar-gamma ligand, protects against burn-induced oxidative injury of remote organs. Burns 2007, 33, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Li, P.K.; Burdmann, E.A.; Mehta, R.L. Acute kidney injury: Acute kidney injury—Global health alert. Nat. Rev. Nephrol. 2013, 9, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.Y.; Lee, J.H.; Park, S.O.; Baek, K.J.; Lee, K.R. Plasma neutrophil gelatinase-associated lipocalin as early biomarker for acute kidney injury in burn patients. J. Burn Care Res. 2013, 34, e326–e332. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.N.; Bao, H.G.; Xu, L.; Wang, X.L.; Shen, Y.; Wang, J.S.; Yang, X.B. Dexmedetomidine protects against ischemia/reperfusion injury in rat kidney. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 1843–1851. [Google Scholar] [PubMed]
- Tasanarong, A.; Kongkham, S.; Itharat, A. Antioxidant effect of phyllanthus emblica extract prevents contrast-induced acute kidney injury. BMC Complement. Altern. Med. 2014, 14, 138. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Kaur, T.; Kaur, A.; Singh, A.P. Glycine aggravates ischemia reperfusion-induced acute kidney injury through n-methyl-d-aspartate receptor activation in rats. Mol. Cell. Biochem. 2014, 393, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Billings, F.T.; Yu, C.; Byrne, J.G.; Petracek, M.R.; Pretorius, M. Heme oxygenase-1 and acute kidney injury following cardiac surgery. Cardiorenal Med. 2014, 4, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Fu, X.J.; Gu, C.; Xu, P.; Wang, Y.; Yu, W.R.; Sun, Q.; Sun, X.J.; Yao, M. Hydrogen-rich saline protects against acute lung injury induced by extensive burn in rat model. J. Burn Care Res. 2011, 32, e82–e91. [Google Scholar] [CrossRef] [PubMed]
- Palipoch, S. A review of oxidative stress in acute kidney injury: Protective role of medicinal plants-derived antioxidants. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 88–93. [Google Scholar] [PubMed]
- Elks, C.M.; Reed, S.D.; Mariappan, N.; Shukitt-Hale, B.; Joseph, J.A.; Ingram, D.K.; Francis, J. A blueberry-enriched diet attenuates nephropathy in a rat model of hypertension via reduction in oxidative stress. PLoS ONE 2011, 6, e24028. [Google Scholar] [CrossRef]
- Shah, S. Role of iron in progressive renal disease. Am. J. Kidney Dis. 2001, 37, S30–S33. [Google Scholar] [CrossRef] [PubMed]
- Rael, L.T.; Bar-Or, R.; Mains, C.W.; Slone, D.S.; Levy, A.S.; Bar-Or, D. Plasma oxidation-reduction potential and protein oxidation in traumatic brain injury. J. Neurotrauma 2009, 26, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Rael, L.T.; Bar-Or, R.; Ambruso, D.R.; Mains, C.W.; Slone, D.S.; Craun, M.L.; Bar-Or, D. The effect of storage on the accumulation of oxidative biomarkers in donated packed red blood cells. J. Trauma 2009, 66, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Guo, S.; Chen, S.; Sun, C.; Zhang, J.; Sun, X. Beneficial effect of hydrogen-rich saline on cerebral vasospasm after experimental subarachnoid hemorrhage in rats. J. Neurosci. Res. 2012, 90, 1670–1680. [Google Scholar] [CrossRef] [PubMed]
- Dagher, P.C.; Mai, E.M.; Hato, T.; Lee, S.Y.; Anderson, M.D.; Karozos, S.C.; Mang, H.E.; Knipe, N.L.; Plotkin, Z.; Sutton, T.A. The p53 inhibitor pifithrin-alpha can stimulate fibrosis in a rat model of ischemic acute kidney injury. Am. J. Physiol. -Ren. 2012, 302, F284–F291. [Google Scholar] [CrossRef]
- Mariano, F.; Cantaluppi, V.; Stella, M.; Romanazzi, G.M.; Assenzio, B.; Cairo, M.; Biancone, L.; Triolo, G.; Ranieri, V.M.; Camussi, G. Circulating plasma factors induce tubular and glomerular alterations in septic burns patients. Crit. Care 2008, 12, R42. [Google Scholar] [CrossRef] [PubMed]
- Rana, A.; Sathyanarayana, P.; Lieberthal, W. Role of apoptosis of renal tubular cells in acute renal failure: Therapeutic implications. Apoptosis 2001, 6, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Wu, M.J.; Sun, H.Y.; Xiong, J.; Zhang, Y.; Liu, C.Y.; Fu, L.L.; Liu, D.M.; Liu, H.Q.; Mei, C.L. Vegf-modified human embryonic mesenchymal stem cell implantation enhances protection against cisplatin-induced acute kidney injury. Am. J. Physiol. Ren. Physiol. 2011, 300, F207–F218. [Google Scholar] [CrossRef]
- Liu, C.M.; Ma, J.Q.; Sun, Y.Z. Puerarin protects rat kidney from lead-induced apoptosis by modulating the pi3k/akt/enos pathway. Toxicol. Appl. Pharmacol. 2012, 258, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Chalah, A.; Khosravi-Far, R. The mitochondrial death pathway. Adv. Exp. Med. Biol. 2008, 615, 25–45. [Google Scholar] [PubMed]
- Rong, Y.; Distelhorst, C.W. Bcl-2 protein family members: Versatile regulators of calcium signaling in cell survival and apoptosis. Ann. Rev. Physiol. 2008, 70, 73–91. [Google Scholar] [CrossRef]
- Small, D.M.; Gobe, G.C. Cytochrome c: Potential as a noninvasive biomarker of drug-induced acute kidney injury. Expert Opin. Drug Metab. Toxicol. 2012, 8, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Huan, J.; Li, W.; Tang, J. Protective effects of ulinastatin on pancreatic and renal damage in rats following early scald injury. Burns 2009, 35, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Abali, A.E.; Cabioglu, T.; Ozdemir, H.; Haberal, M. Interactive effects of acupuncture on pain and distress in major burns: An experiment with rats. Burns 2014, in press. [Google Scholar]
- Yuan, X.; Jing, S.; Wu, L.; Chen, L.; Fang, J. Pharmacological postconditioning with tanshinone iia attenuates myocardial ischemia-reperfusion injury in rats by activating the phosphatidylinositol 3-kinase pathway. Exp. Ther. Med. 2014, 8, 973–977. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, S.-X.; Zhou, H.-L.; Huang, C.-L.; You, C.-G.; Fang, Q.; Wu, P.; Wang, X.-G.; Han, C.-M. Astaxanthin Attenuates Early Acute Kidney Injury Following Severe Burns in Rats by Ameliorating Oxidative Stress and Mitochondrial-Related Apoptosis. Mar. Drugs 2015, 13, 2105-2123. https://doi.org/10.3390/md13042105
Guo S-X, Zhou H-L, Huang C-L, You C-G, Fang Q, Wu P, Wang X-G, Han C-M. Astaxanthin Attenuates Early Acute Kidney Injury Following Severe Burns in Rats by Ameliorating Oxidative Stress and Mitochondrial-Related Apoptosis. Marine Drugs. 2015; 13(4):2105-2123. https://doi.org/10.3390/md13042105
Chicago/Turabian StyleGuo, Song-Xue, Han-Lei Zhou, Chun-Lan Huang, Chuan-Gang You, Quan Fang, Pan Wu, Xin-Gang Wang, and Chun-Mao Han. 2015. "Astaxanthin Attenuates Early Acute Kidney Injury Following Severe Burns in Rats by Ameliorating Oxidative Stress and Mitochondrial-Related Apoptosis" Marine Drugs 13, no. 4: 2105-2123. https://doi.org/10.3390/md13042105