

HSPB6 Is Depleted in Colon Cancer Patients and Its Expression Is Induced by 5-aza-2′-Deoxycytidine In Vitro
 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis
2.2. Ethical Approval
2.3. Sample Collections
2.4. Induction of Colon Cancer in Wistar Rats
2.5. Cell Culture and 5-Aza-2′-Deoxycytidine Induction
2.6. Genomic DNA Extraction, Bisulfite Treatment, and Methylation-Specific PCR (MSP)
2.7. RNA Extraction, cDNA Synthesis, and qRT-PCR
2.8. Statistical Analysis
3. Results
3.1. Clinical Data of the Enrolled Patients
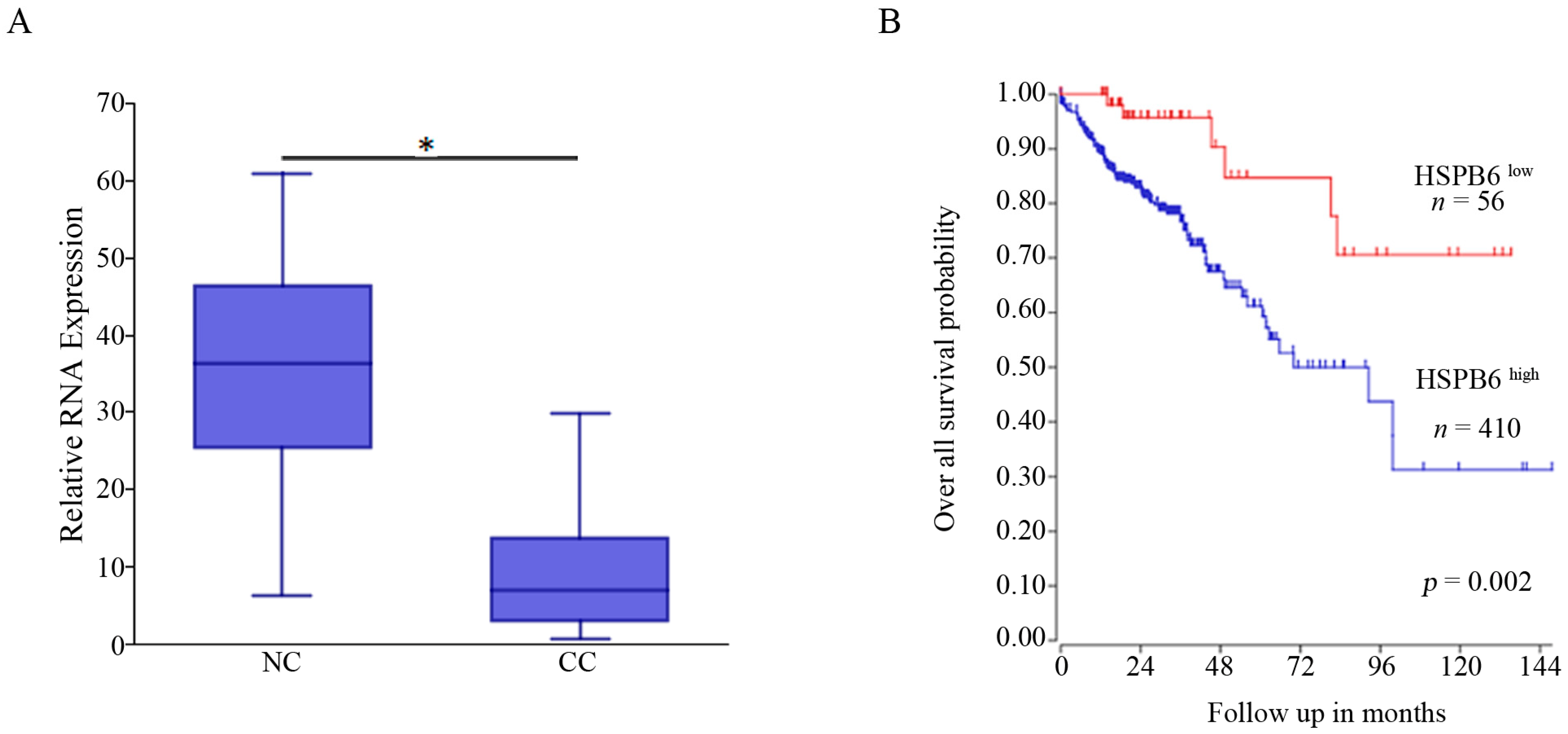
3.2. HSPB6 Is Downregulated in CC and Associated with Poor Outcomes
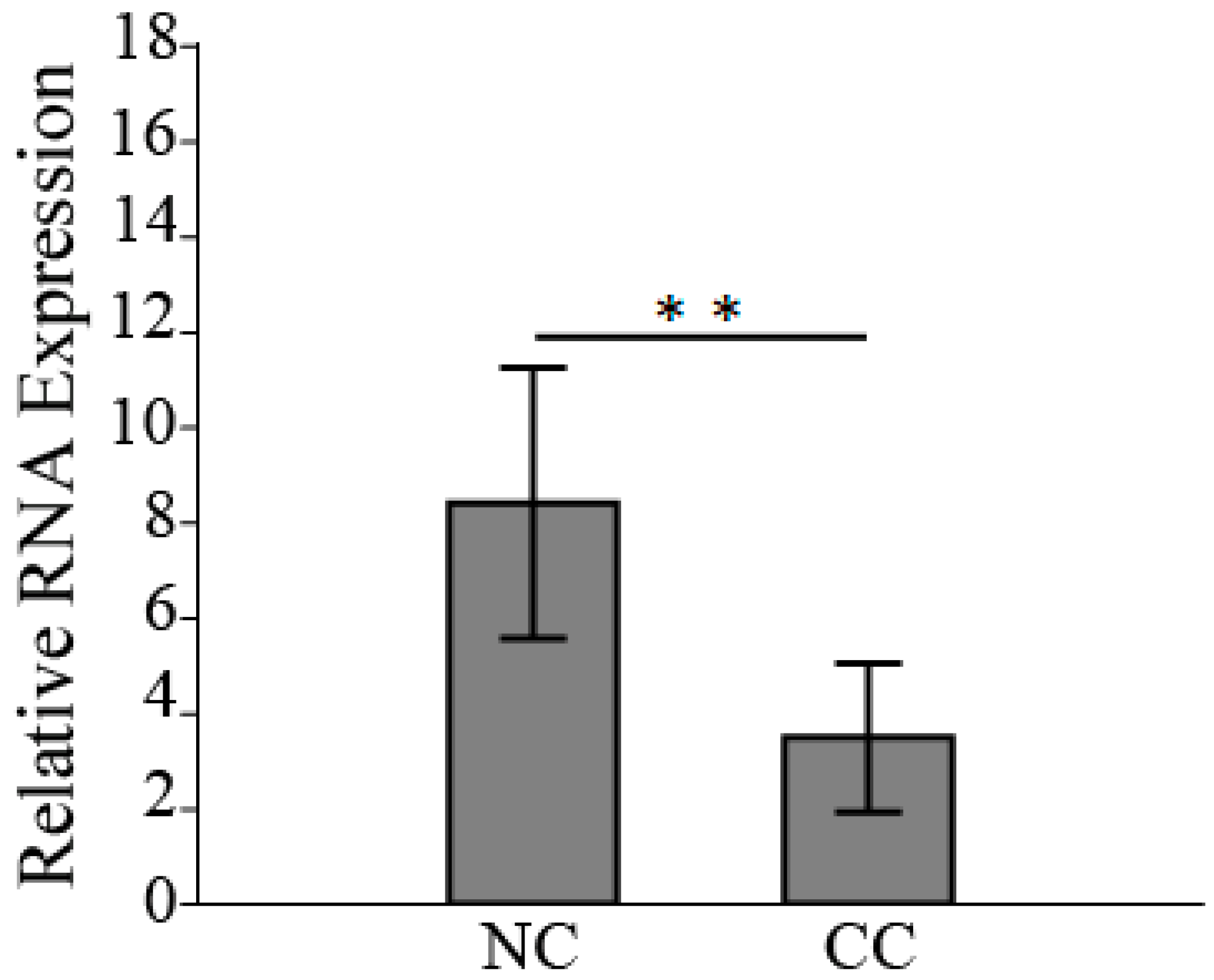
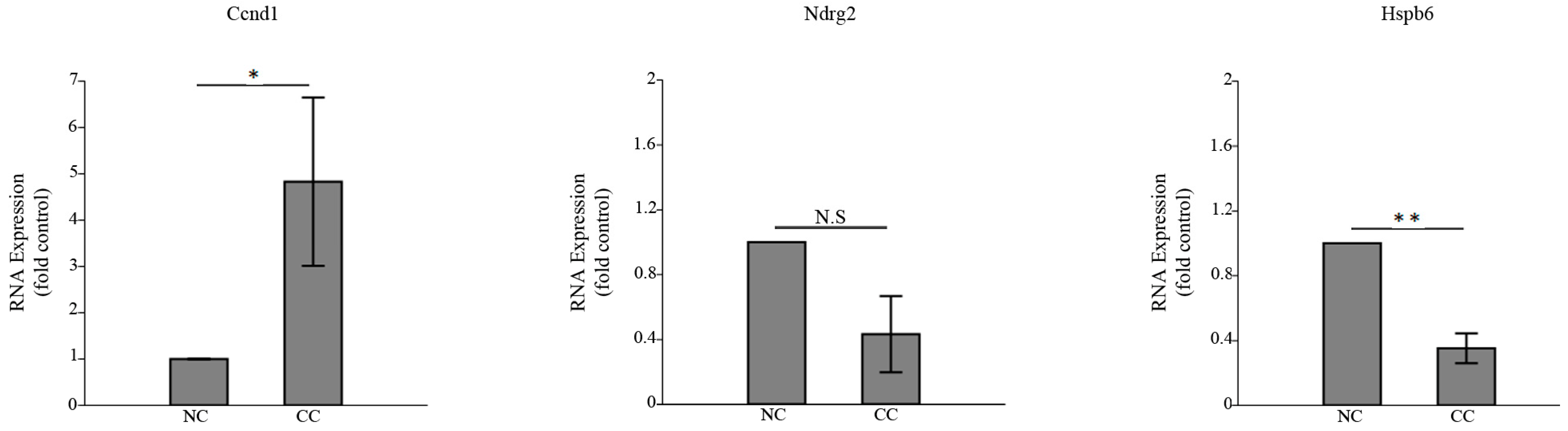
3.3. HSPB6 Depletion Is Observed in Colon Tumorigenesis
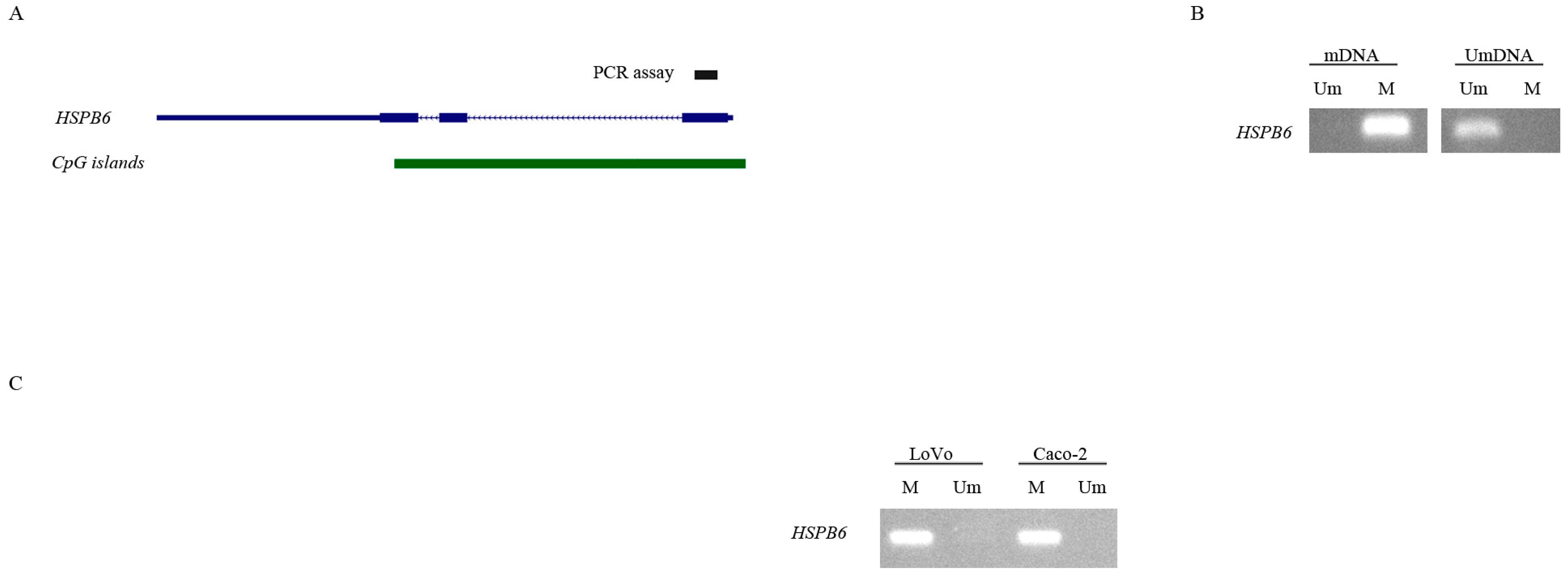
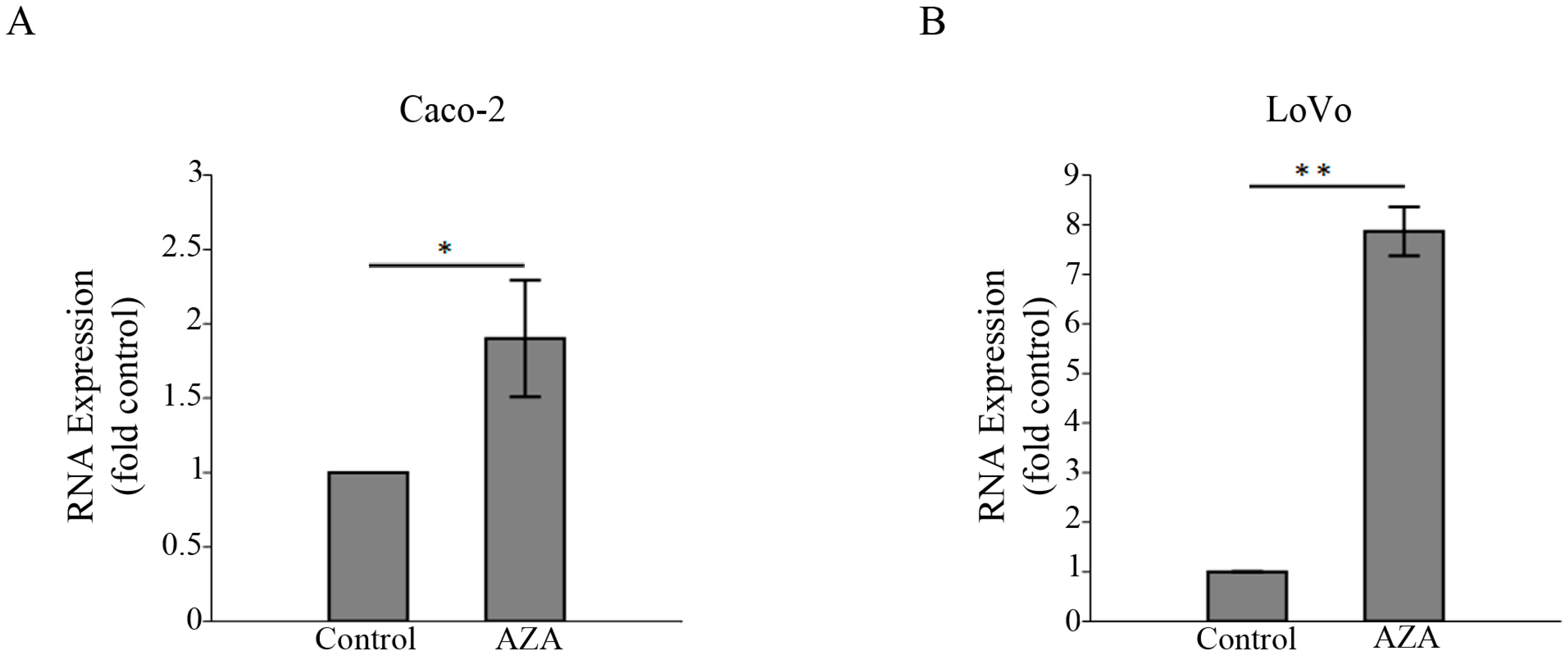
3.4. 5-aza-2′-Deoxycytidine Induces the Expression of HSPB6
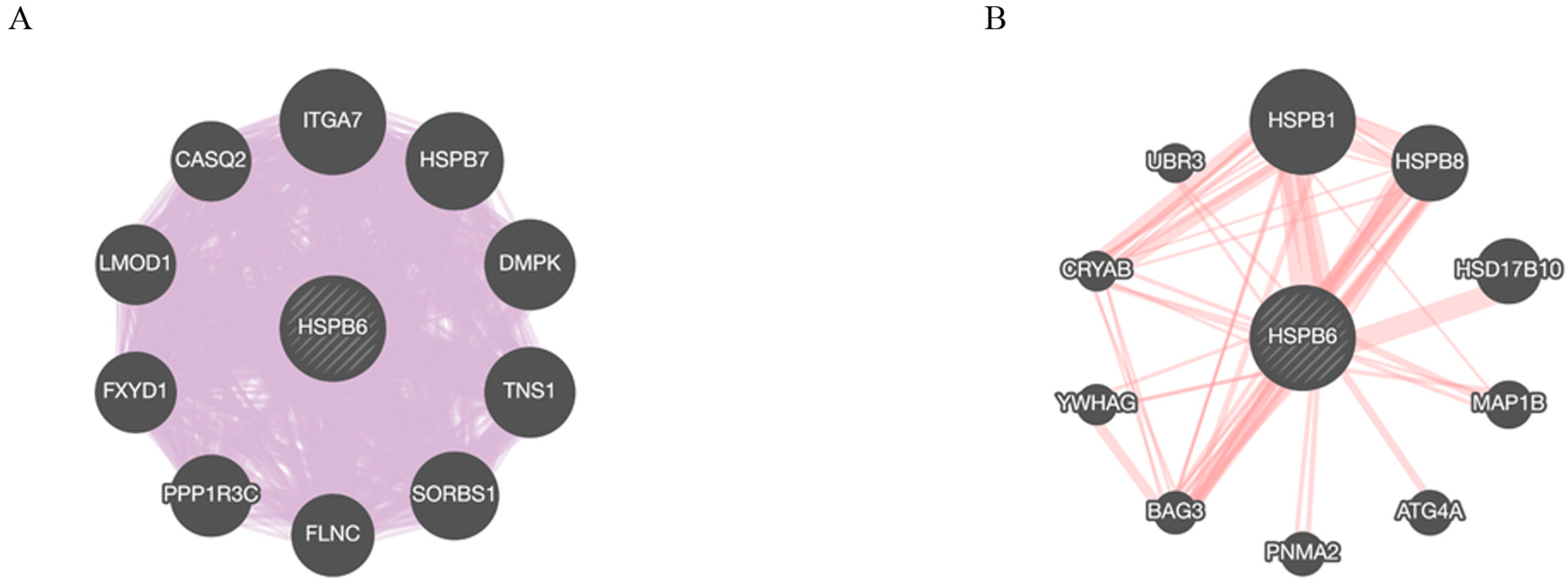
3.5. HSPB6 Gene and Protein Interactions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, E.A.; Castillo, B.M.; García, E.C.; Tascón, J.; Morales, A.I. Colon Cancer Pharmacogenetics: A Narrative Review. Pharmacy 2022, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, W.S.; Almufareh, N.A.; Domiaty, D.M.; Albasher, G.; Alduwish, M.A.; Alkhalaf, H.; Almuzzaini, B.; AL-marshidy, S.S.; Alfraihi, R.; Elasbali, A.M.; et al. Epidemiology of cancer in Saudi Arabia thru 2010–2019: A systematic review with constrained meta-analysis. AIMS Public Health 2020, 7, 679–696. [Google Scholar] [PubMed]
- Almatroudi, A. The Incidence Rate of Colorectal Cancer in Saudi Arabia: An Observational Descriptive Epidemiological Analysis. Int. J. Gen. Med. 2020, 13, 977–990. [Google Scholar] [CrossRef]
- Al-Zalabani, A. Preventability of Colorectal Cancer in Saudi Arabia: Fraction of Cases Attributable to Modifiable Risk Factors in 2015–2040. Int. J. Environ. Res. Public Health 2020, 17, 320. [Google Scholar] [CrossRef]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef]
- Cui, S.; Chang, P.-Y. Current understanding concerning intestinal stem cells. World J. Gastroenterol. 2016, 22, 7099–7110. [Google Scholar] [CrossRef]
- Jahanafrooz, Z.; Mosafer, J.; Akbari, M.; Hashemzaei, M.; Mokhtarzadeh, A.; Baradaran, B. Colon cancer therapy by focusing on colon cancer stem cells and their tumor microenvironment. J. Cell. Physiol. 2019, 235, 4153–4166. [Google Scholar] [CrossRef]
- Xu, J.; Chen, J.; Cheng, C.; Li, H.; Gao, P.; Zheng, J.; Wang, J. Screening Gene Expression-Related Alternative Splicing Event Signature for Colon Cancer Prognostic Prediction. J. Oncol. 2022, 2022, 9952438. [Google Scholar] [CrossRef]
- Sammut, S.J.; Feichtinger, J.; Stuart, N.; Wakeman, J.A.; Larcombe, L.; McFarlane, R.J. A novel cohort of cancer-testis biomarker genes revealed through meta-analysis of clinical data sets. Oncoscience 2014, 1, 349–359. [Google Scholar] [CrossRef]
- Almutairi, M.H.; Alrubie, T.M.; Alamri, A.M.; Almutairi, B.O.; Alrefaei, A.F.; Arafah, M.M.; Alanazi, M.; Semlali, A. Cancer-Testis Gene Biomarkers Discovered in Colon Cancer Patients. Genes 2022, 13, 807. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.D.; Xiong, W.; Bunker, A.M.; Vaughn, C.P.; Furtado, L.V.; Roberts, W.L.; Fang, J.C.; Samowitz, W.S.; A Heichman, K. Septin 9 methylated DNA is a sensitive and specific blood test for colorectal cancer. BMC Med. 2011, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.-C.; Kim, J.-Y.; Park, J.W.; Kim, K.-B.; Oh, H.; Lee, K.-H.; Seo, S.-B. FOXO1 degradation via G9a-mediated methylation promotes cell proliferation in colon cancer. Nucleic Acids Res. 2018, 47, 1692–1705. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, M.H.; Alotaibi, M.M.; Alonaizan, R.; Almutairi, B.O. Expression Profile of MAGE-B1 Gene and Its Hypomethylation Activation in Colon Cancer. BioMed Res. Int. 2022, 2022, 6066567. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Liu, X.; Li, M.; Zhao, Y.; Li, J.; Wen, Z.; Liu, M.; Yang, M.; Fu, B.; Wei, M. Analysis of 5-Methylcytosine Regulators and DNA Methylation-Driven Genes in Colon Cancer. Front. Cell Dev. Biol. 2022, 9, 657092. [Google Scholar] [CrossRef]
- Koch, A.; Joosten, S.C.; Feng, Z.; De Ruijter, T.C.; Draht, M.X.; Melotte, V.; Smits, K.M.; Veeck, J.; Herman, J.G.; Van Neste, L.; et al. Analysis of DNA methylation in cancer: Location revisited. Nat. Rev. Clin. Oncol. 2018, 15, 459–466. [Google Scholar] [CrossRef]
- nbsp; Rappa, F.; Farina, F.; Zummo, G.; David, S.; Campanella, C.; Carini, F.; Tomasello, G.; Damiani, P.; Cappello, F.; et al. HSP-molecular chaperones in cancer biogenesis and tumor therapy: An overview. Anticancer. Res. 2012, 32, 5139–5150. [Google Scholar]
- Hoter, A.; Naim, H.Y. Heat Shock Proteins and Ovarian Cancer: Important Roles and Therapeutic Opportunities. Cancers 2019, 11, 1389. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Large Potentials of Small Heat Shock Proteins. Physiol. Rev. 2011, 91, 1123–1159. [Google Scholar] [CrossRef]
- Qiao, N.; Zhu, Y.; Li, H.; Qu, Z.; Xiao, Z. Expression of heat shock protein 20 inversely correlated with tumor progression in patients with ovarian cancer. Eur. J. Gynaecol. Oncol. 2014, 35, 576–579. [Google Scholar]
- Ju, Y.-T.; Kwag, S.-J.; Park, H.J.; Jung, E.-J.; Jeong, C.-Y.; Jeong, S.-H.; Lee, Y.-J.; Choi, S.-K.; Kang, K.R.; Hah, Y.-S.; et al. Decreased Expression of Heat Shock Protein 20 in Colorectal Cancer and Its Implication in Tumorigenesis. J. Cell. Biochem. 2014, 116, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Duan, Y.; Meng, Q.-H.; Gong, R.; Guo, C.; Zhao, Y.; Zhang, Y. Integrated analysis of DNA methylation profiling and gene expression profiling identifies novel markers in lung cancer in Xuanwei, China. PLoS ONE 2018, 13, e0203155. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhuang, L.; Szatmary, P.; Wen, L.; Sun, H.; Lu, Y.; Xu, Q.; Chen, X. Upregulation of Heat Shock Proteins (HSPA12A, HSP90B1, HSPA4, HSPA5 and HSPA6) in Tumour Tissues Is Associated with Poor Outcomes from HBV-Related Early-Stage Hepatocellular Carcinoma. Int. J. Med. Sci. 2015, 12, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Matsushima-Nishiwaki, R.; Toyoda, H.; Matsuura, J.; Kumada, T.; Kozawa, O. Heat shock protein 20 (HSPB6) regulates apoptosis in human hepatocellular carcinoma cells: Direct association with Bax. Oncol. Rep. 2014, 32, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, T.; Xie, F.; Wang, L.; Liang, Z.; Li, D.; Liang, Y.; Zhao, K.; Qi, X.; Yang, X.; et al. Evaluating the biological functions of the prognostic genes identified by the Pathology Atlas in bladder cancer. Oncol. Rep. 2020, 45, 191–201. [Google Scholar] [CrossRef]
- Uhlén, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, 2507. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, Q.; Su, Y.; Pan, L. Network-Based Differential Analysis to Identify Molecular Features of Tumorigenesis for Esophageal Squamous Carcinoma. Molecules 2018, 23, 88. [Google Scholar] [CrossRef]
- Wyciszkiewicz, A.; Lach, M.S.; Wróblewska, J.P.; Michalak, M.; Suchorska, W.M.; Kalinowska, A.; Michalak, S. The involvement of small heat shock protein in chemoresistance in ovarian cancer—In vitro study. EXCLI J. 2021, 20, 935–947. [Google Scholar] [CrossRef]
- Koga, Y.; Pelizzola, M.; Cheng, E.; Krauthammer, M.; Sznol, M.; Ariyan, S.; Narayan, D.; Molinaro, A.M.; Halaban, R.; Weissman, S.M. Genome-wide screen of promoter methylation identifies novel markers in melanoma. Genome Res. 2009, 19, 1462–1470. [Google Scholar] [CrossRef]
- Franz, M.; Rodriguez, H.; Lopes, C.; Zuberi, K.; Montojo, J.; Bader, G.D.; Morris, Q. GeneMANIA update 2018. Nucleic Acids Res. 2018, 46, W60–W64. [Google Scholar] [CrossRef]
- Alghamdi, H.A.; Al-Zharani, M.; Aljarba, N.H.; Alghamdi, A.A.; Alghamdi, A.A.; Aldahmash, B.A.; Elnagar, D.M.; Alkahtani, S. Efficacy of ivermectin against colon cancer induced by dimethylhydrazine in male wistar rats. Saudi Pharm. J. 2022, 30, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Chari, K.Y.; Polu, P.R.; Shenoy, R.R. An Appraisal of Pumpkin Seed Extract in 1, 2-Dimethylhydrazine Induced Colon Cancer in Wistar Rats. J. Toxicol. 2018, 2018, 6086490. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, B.; Charlet, J.; Dallosso, A.R.; Szemes, M.; Etchevers, H.C.; Malik, K.T.A.; Brown, K.W. Epigenetic deregulation of GATA3 in neuroblastoma is associated with increased GATA3 protein ex-pression and with poor outcomes. Sci. Rep. 2019, 9, 18934. [Google Scholar] [CrossRef] [PubMed]
- Charlet, J.; Tomari, A.; Dallosso, A.R.; Szemes, M.; Kaselova, M.; Curry, T.J.; Almutairi, B.; Etchevers, H.C.; McConville, C.; Malik, K.T.A.; et al. Genome-wide DNA methylation analysis identifies MEGF10 as a novel epigenetically repressed candidate tumor suppressor gene in neuroblastoma. Mol. Carcinog. 2016, 56, 1290–1301. [Google Scholar] [CrossRef]
- Li, L.-C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef]
- Almutairi, B.O.; Almutairi, M.H.; Alrefaei, A.F.; Ali, D.; Alkahtani, S.; Alarifi, S. Cigarette Smoke Regulates the Expression of EYA4 via Alternation of DNA Methylation Status. BioMed Res. Int. 2022, 2022, 5032172. [Google Scholar] [CrossRef]
- Combes, J.-D.; Grelier, G.; Laversanne, M.; Voirin, N.; Chabaud, S.; Ecochard, R.; Lasset, C.; Moyret-Lalle, C. Contribution of cell culture, RNA extraction, and reverse transcription to the measurement error in quantitative reverse transcription polymerase chain reaction-based gene expression quantification. Anal. Biochem. 2009, 393, 29–35. [Google Scholar] [CrossRef]
- Chen, H.-P.; Pan, M.-H.; Chou, Y.-Y.; Sung, C.; Lee, K.-H.; Leung, C.-M.; Hsu, P.-C. Effects of di(2-ethylhexyl)phthalate exposure on 1,2-dimethyhydrazine-induced colon tumor promotion in rats. Food Chem. Toxicol. 2017, 103, 157–167. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wu, L.; Zhang, Z.; Han, T.; Guo, H.; Jiang, N.; Tao, K.; Ti, Z.; Liu, X.; et al. NDRG2 in rat liver regeneration: Role in proliferation and apoptosis. Wound Repair Regen. 2010, 18, 524–531. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Riedl, M.; Peters, C.; Weinkauf, S.; Haslbeck, M.; Buchner, J. Regulation of small heat-shock proteins by hetero-oligomer formation. J. Biol. Chem. 2020, 295, 158–169. [Google Scholar] [CrossRef]
- Boland, C.R.; Goel, A. Microsatellite instability in colorectal cancer. Gastroenterology 2010, 138, 2073–2087.e3. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, Y.; Isomoto, H.; Oka, M.; Yano, Y.; Soda, H.; Shikuwa, S.; Takeshima, F.; Omagari, K.; Mizuta, Y.; Murase, K.; et al. Expression of Heat Shock Protein (Hsp) 70 and Hsp 40 in Colorectal Cancer. Med. Oncol. 2003, 20, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Hwang, T.S.; Han, H.S.; Choi, H.K.; Lee, Y.J.; Kim, Y.-J.; Han, M.-Y.; Park, Y.-M. Differential, stage-dependent expression of Hsp70, Hsp110 and Bcl-2 in colorectal cancer. J. Gastroenterol. Hepatol. 2003, 18, 690–700. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, X.; Cai, R.; Liu, M.; Wang, R. Identification of a tumor microenvironment-associated prognostic gene signature in bladder cancer by inte-grated bioinformatic analysis. Int. J. Clin. Exp. Pathol. 2021, 14, 551–566. [Google Scholar]
- Chen, S.; Huang, H.; Yao, J.; Pan, L.; Ma, H. Heat shock protein B6 potently increases non-small cell lung cancer growth. Mol. Med. Rep. 2014, 10, 677–682. [Google Scholar] [CrossRef]
- Christman, J.K. 5-Azacytidine and 5-aza-2′-deoxycytidine as inhibitors of DNA methylation: Mechanistic studies and their implications for cancer therapy. Oncogene 2002, 21, 5483–5495. [Google Scholar] [CrossRef]
- Pierdomenico, M.; Cesi, V.; Cucchiara, S.; Vitali, R.; Prete, E.; Costanzo, M.; Aloi, M.; Oliva, S.; Stronati, L. NOD2 Is Regulated By Mir-320 in Physiological Conditions but this Control Is Altered in Inflamed Tissues of Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016, 22, 11. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Shi, E.; Yan, L.; Li, J.; Jiang, X. Inhibition of micro-ribonucleic acid-320 attenuates neurologic injuries after spinal cord ischemia. J. Thorac. Cardiovasc. Surg. 2015, 150, 398–406. [Google Scholar] [CrossRef]
- Matsushima-Nishiwaki, R.; Adachi, S.; Yoshioka, T.; Yasuda, E.; Yamagishi, Y.; Matsuura, J.; Muko, M.; Iwamura, R.; Noda, T.; Toyoda, H.; et al. Suppression by heat shock protein 20 of hepatocellular carcinoma cell proliferation via inhibition of the mitogen-activated protein kinases and AKT pathways. J. Cell. Biochem. 2011, 112, 3430–3439. [Google Scholar] [CrossRef]
- Matsushima-Nishiwaki, R.; Kumada, T.; Nagasawa, T.; Suzuki, M.; Yasuda, E.; Okuda, S.; Maeda, A.; Kaneoka, Y.; Toyoda, H.; Kozawa, O. Direct Association of Heat Shock Protein 20 (HSPB6) with Phosphoinositide 3-kinase (PI3K) in Human Hepatocellular Carcinoma: Regulation of the PI3K Activity. PLoS ONE 2013, 8, e78440. [Google Scholar] [CrossRef]
- Matsushima-Nishiwaki, R.; Takai, S.; Adachi, S.; Minamitani, C.; Yasuda, E.; Noda, T.; Kato, K.; Toyoda, H.; Kaneoka, Y.; Yamaguchi, A.; et al. Phosphorylated Heat Shock Protein 27 Represses Growth of Hepatocellular Carcinoma via Inhibition of Extracellular Signal-regulated Kinase. J. Biol. Chem. 2008, 283, 18852–18860. [Google Scholar] [CrossRef] [PubMed]
- Mallon, B.S.; Chenoweth, J.G.; Johnson, K.R.; Hamilton, R.S.; Tesar, P.J.; Yavatkar, A.S.; Tyson, L.J.; Park, K.; Chen, K.G.; Fann, Y.C.; et al. StemCellDB: The Human Pluripotent Stem Cell Database at the National Institutes of Health. Stem Cell Res. 2013, 10, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Roth, R.B.; Hevezi, P.; Lee, J.; Willhite, D.; Lechner, S.M.; Foster, A.C.; Zlotnik, A. Gene expression analyses reveal molecular relationships among 20 regions of the human CNS. Neurogenetics 2006, 7, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.F.; Huang, X.; McKercher, S.R.; Zaidi, R.; Okamoto, S.-I.; Nakanishi, N.; Lipton, S.A. Transcriptional profiling of MEF2-regulated genes in human neural progenitor cells derived from embryonic stem cells. Genom. Data 2015, 3, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, D.; Zafari, Y.; Estaki, Z.; Mehrabi, M.; Moghbelinejad, S. Evaluation of plasma circ_0006282 as a novel diagnostic biomarker in colorectal cancer. J. Clin. Lab. Anal. 2021, 36, e24147. [Google Scholar] [CrossRef]
- Yu, J.; Yang, K.; Zheng, J.; Sun, X.; Zhao, W. Establishment of a novel prognostic signature based on an identified expression profile of integrin superfamily to predict overall survival of patients with colorectal adenocarcinoma. Gene 2021, 808, 145990. [Google Scholar] [CrossRef]
- Huttlin, E.L.; Ting, L.; Bruckner, R.J.; Gebreab, F.; Gygi, M.P.; Szpyt, J.; Tam, S.; Zarraga, G.; Colby, G.; Baltier, K.; et al. The BioPlex Network: A Systematic Exploration of the Human Interactome. Cell 2015, 162, 425–440. [Google Scholar] [CrossRef]
- Huttlin, E.L.; Bruckner, R.J.; Paulo, J.A.; Cannon, J.R.; Ting, L.; Baltier, K.; Colby, G.; Gebreab, F.; Gygi, M.P.; Parzen, H.; et al. Architecture of the human interactome defines protein communities and disease networks. Nature 2017, 545, 505–509. [Google Scholar] [CrossRef]
- Arrigo, A.-P.; Gibert, B. Protein interactomes of three stress inducible small heat shock proteins: HspB1, HspB5 and HspB8. Int. J. Hyperth. 2013, 29, 409–422. [Google Scholar] [CrossRef]
- Weeks, S.D.; Muranova, L.K.; Heirbaut, M.; Beelen, S.; Strelkov, S.V.; Gusev, N.B. Characterization of human small heat shock protein HSPB1 α-crystallin domain localized mutants associated with hereditary motor neuron diseases. Sci. Rep. 2018, 8, 688. [Google Scholar] [CrossRef]
- Tedesco, B.; Cristofani, R.; Ferrari, V.; Cozzi, M.; Rusmini, P.; Casarotto, E.; Chierichetti, M.; Mina, F.; Galbiati, M.; Piccolella, M.; et al. Insights on Human Small Heat Shock Proteins and Their Alterations in Diseases. Front. Mol. Biosci. 2022, 9, 842149. [Google Scholar] [CrossRef] [PubMed]
- Matsushima-Nishiwaki, R.; Toyoda, H.; Maeda, A.; Kaneoka, Y.; Kumada, T.; Kozawa, O. Correlation between the complex of small heat shock proteins (HSPBs) and the progression in patients with hepatocellular carcinoma. Arch. Biochem. Biophys. 2022, 732, 109461. [Google Scholar] [CrossRef] [PubMed]
- Heirbaut, M.; Lermyte, F.; Martin, E.M.; Beelen, S.; Sobott, F.; Strelkov, S.V.; Weeks, S.D. Specific sequences in the N-terminal domain of human small heat-shock protein HSPB6 dictate preferential hetero-oligomerization with the orthologue HSPB1. J. Biol. Chem. 2017, 292, 9944–9957. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense Region (5′–3′) | Antisense Region (5′–3′) | Ta 1 | Product Size (bp) |
|---|---|---|---|---|
| HSPB6 | CTTCCAGACCCTACCAGCAC | CCAAGAAAGTGGGTCGGTGG | 58 | 71 |
| GAPDH | GGGAAGCTTGTCATCAATGG | GAGATGATGACCCTTTTGGC | 173 | |
| Ccdn1 | TCCGGAGACCGGTCGTAGA | GTCTTAAGCATGGCTCGCAG | 194 | |
| Ndrg2 | GCTTGTTGTCCAACTTCTCAC | GAATGAGTCTGTCCCTGGTCC | 192 | |
| Hspb6 | ATGAGGAGCGCCCAGATGAA | GGAGACAGTGCAGAGGTCAC | 106 | |
| Gapdh | CAACTCCCTCAAGATTGTCAGC | GGCATGGACTGTGGTCATGA | 180 |
| Parameters | NC | CC | ||
|---|---|---|---|---|
| Number of patients | 10 (100%) | 10 (100%) | ||
| Average of the age (youngest–oldest) | 57 years and 8 months (24–79) | |||
| Above 57 years and 8 months | 6 (60%) | 6 (60%) | ||
| Below 57 years and 8 months | 4 (40%) | 4 (40%) | ||
| The age, gender, TNM, and cancer grade of each patient | ||||
| Number of patients | Age | Gender | TNM staging | Cancer Grade |
| 1 | 79 | Male | T2N1M0 | 2 |
| 2 | 54 | Male | T3N0Mx | 2 |
| 3 | 24 | Male | T3N0Mx | 2 |
| 4 | 69 | Male | T2N2M0 | 2 |
| 5 | 61 | Male | T2N0Mx | 2 |
| 6 | 38 | Male | T3N1Mx | 1 |
| 7 | 73 | Male | T2N0Mx | 2 |
| 8 | 79 | Male | T3N0Mx | 2 |
| 9 | 38 | Male | T3N1Mx | 2 |
| 10 | 63 | Male | T3N0Mx | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almutairi, B.O.; Almutairi, M.H.; Alrefaei, A.F.; Alkahtani, S.; Alarifi, S. HSPB6 Is Depleted in Colon Cancer Patients and Its Expression Is Induced by 5-aza-2′-Deoxycytidine In Vitro. Medicina 2023, 59, 996. https://doi.org/10.3390/medicina59050996
Almutairi BO, Almutairi MH, Alrefaei AF, Alkahtani S, Alarifi S. HSPB6 Is Depleted in Colon Cancer Patients and Its Expression Is Induced by 5-aza-2′-Deoxycytidine In Vitro. Medicina. 2023; 59(5):996. https://doi.org/10.3390/medicina59050996
Chicago/Turabian StyleAlmutairi, Bader O., Mikhlid H. Almutairi, Abdulwahed F. Alrefaei, Saad Alkahtani, and Saud Alarifi. 2023. "HSPB6 Is Depleted in Colon Cancer Patients and Its Expression Is Induced by 5-aza-2′-Deoxycytidine In Vitro" Medicina 59, no. 5: 996. https://doi.org/10.3390/medicina59050996