
Higher Expression Levels of SSX1 and SSX2 in Patients with Colon Cancer: Regulated In Vitro by the Inhibition of Methylation and Histone Deacetylation
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval and Sample Collection
2.2. Sources and Cultures of Human CC Cell Lines and Their Treatments with Epigenetic Drugs (5-aza-CdR or TSA)
2.3. RNA Isolation from NC, CC, BC, CLL, NB, and Cultured Cells
2.4. Synthesis of cDNA
2.5. Design of RT-PCR Primers, RT-PCR Conditions, and Agarose Gel Electrophoresis of RT-PCR Products
2.6. Design of qRT-PCR Primers and qRT-PCR Setups
2.7. Statistical Analysis
2.8. In Silico Analysis
2.9. The Cancer Genome Atlas (TCGA) Database Analysis
3. Results
3.1. Clinical Parameters of the Study Participants
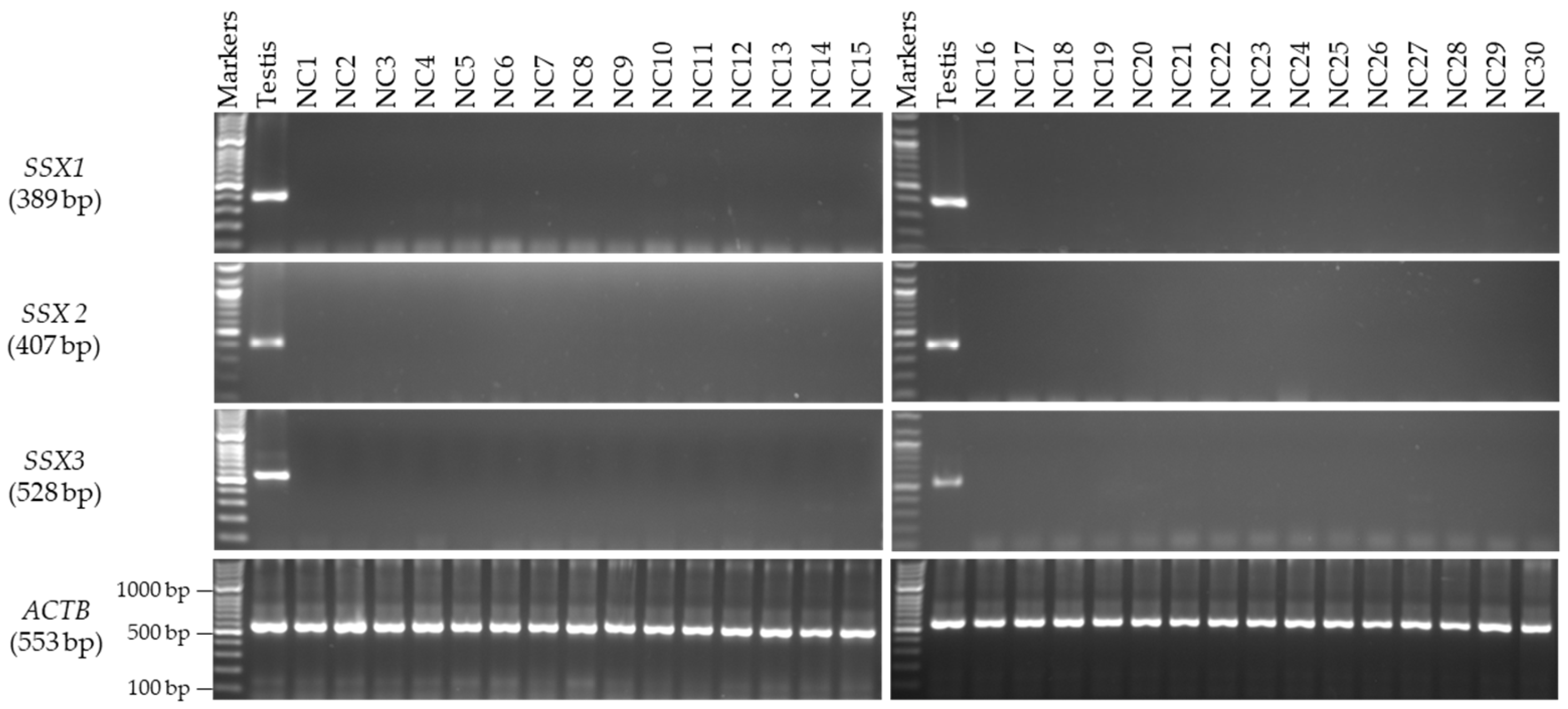
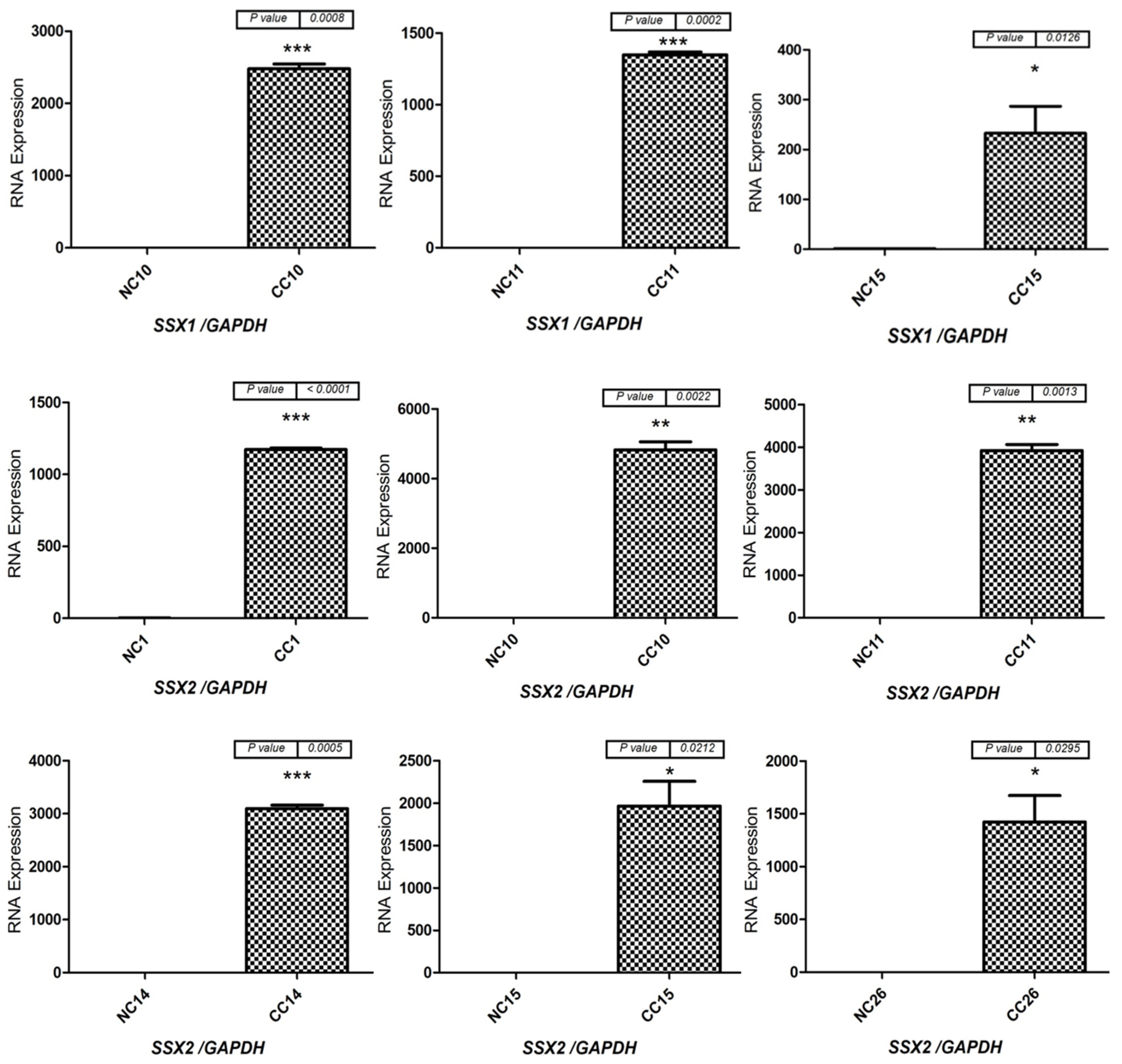
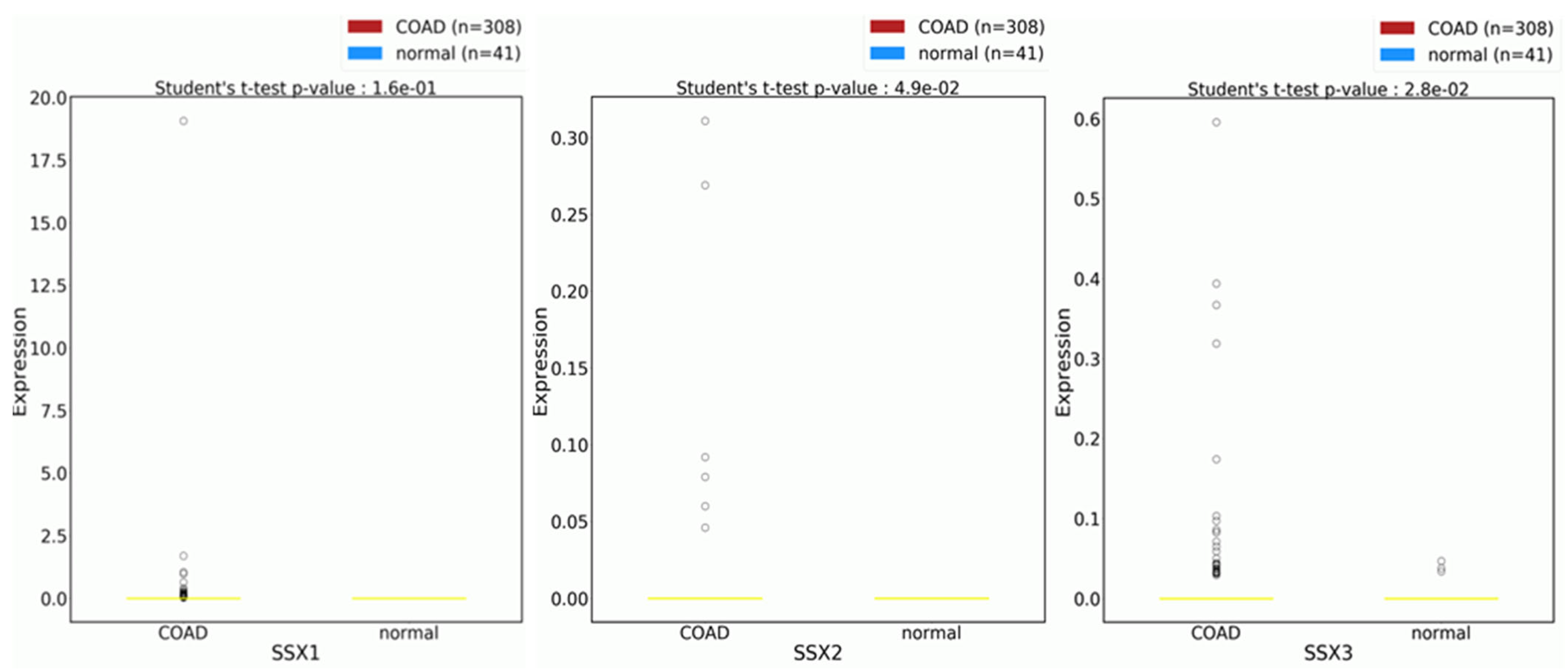
3.2. Expression Profiles of the SSX1, SSX2, and SSX3 Genes in the Matched CC and NC Tissues from the Male and Female Patients
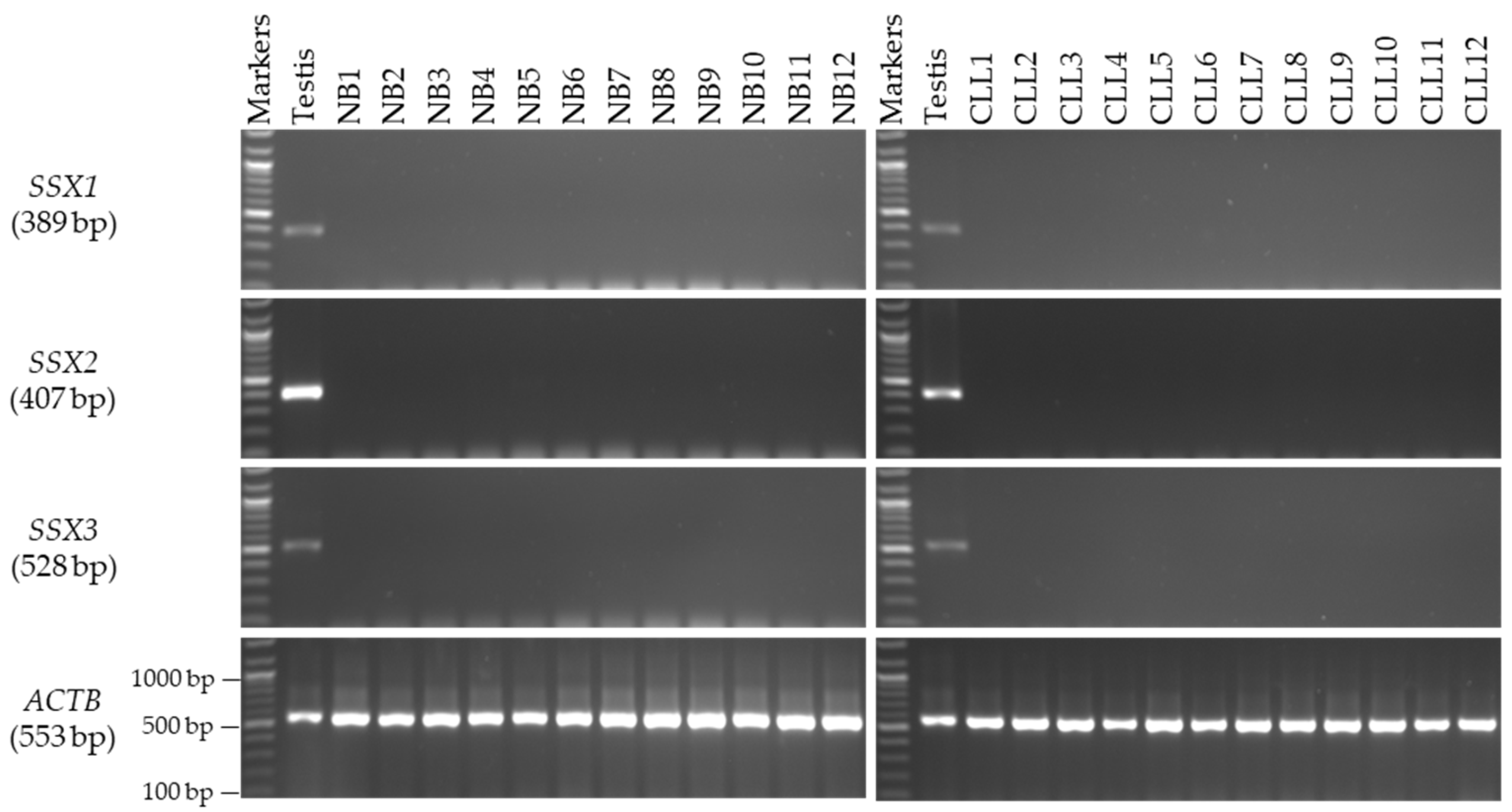
3.3. Screening of the SSX Genes in CLL and BC Tissue Samples
3.4. Effects of 5-aza-2′-CdR and TSA on SSX Gene Expressions in CC Cell Lines
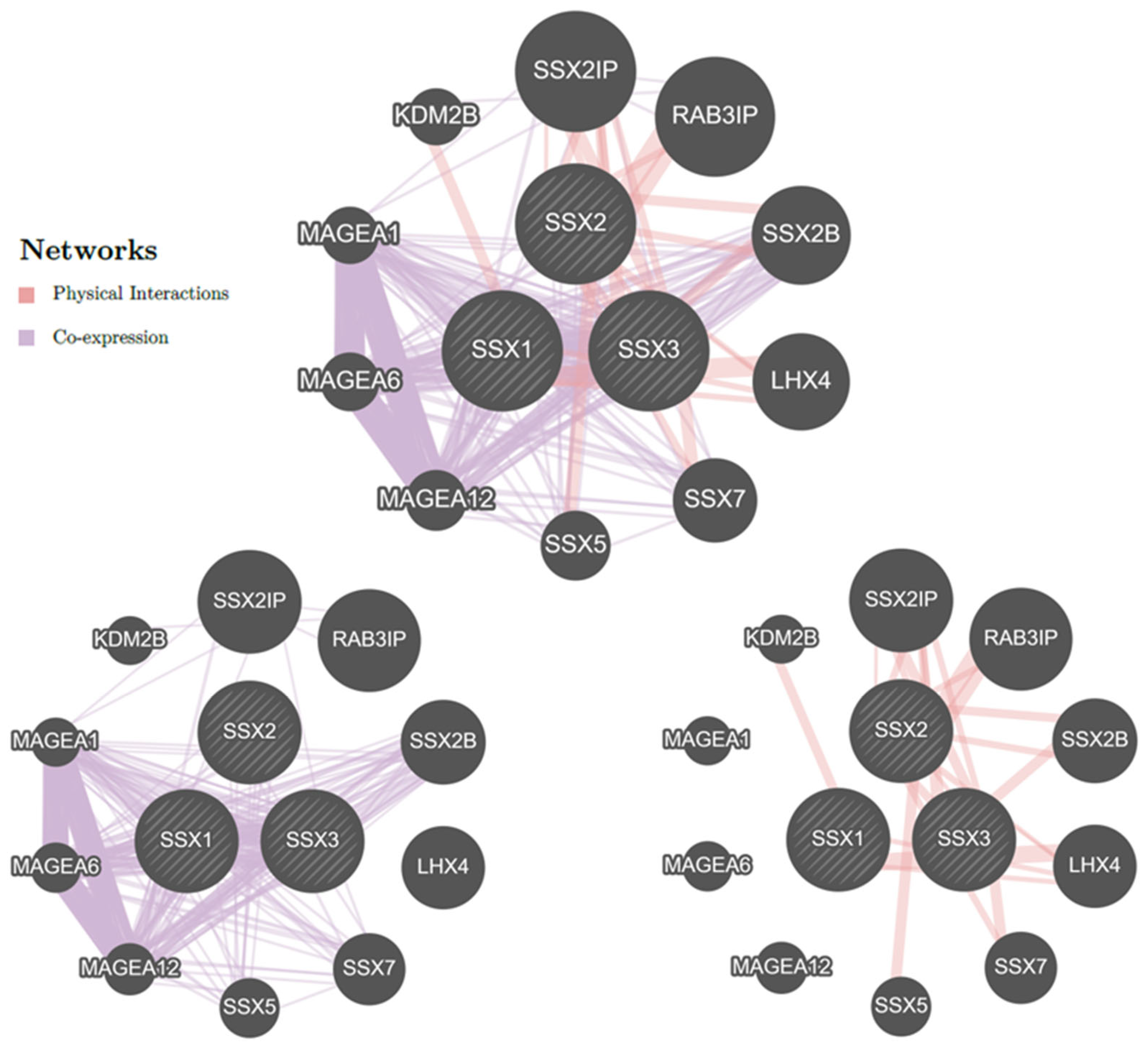
3.5. Gene–Gene Interaction Network
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Araghi, M.; Soerjomataram, I.; Jenkins, M.; Brierley, J.; Morris, E.; Bray, F.; Arnold, M. Global trends in colorectal cancer mortality: Projections to the year 2035. Int. J. Cancer 2019, 144, 2992–3000. [Google Scholar] [CrossRef]
- Almutairi, M.H.; Alotaibi, M.M.; Alonaizan, R.; Almutairi, B.O. Expression Profile of MAGE-B1 Gene and Its Hypomethylation Activation in Colon Cancer. Biomed. Res. Int. 2022, 2022, 6066567. [Google Scholar] [CrossRef]
- Alsanea, N.; Abduljabbar, A.S.; Alhomoud, S.; Ashari, L.H.; Hibbert, D.; Bazarbashi, S. Colorectal cancer in Saudi Arabia: Incidence, survival, demographics and implications for national policies. Ann. Saudi. Med. 2015, 35, 196–202. [Google Scholar] [CrossRef]
- Rasool, M.; Natesan Pushparaj, P.; Karim, S. Overexpression of CXCL8 gene in Saudi colon cancer patients. Saudi. J. Biol. Sci. 2021, 28, 6045–6049. [Google Scholar] [CrossRef]
- Kanojia, D.; Garg, M.; Gupta, S.; Gupta, A.; Suri, A. Sperm-associated antigen 9 is a novel biomarker for colorectal cancer and is involved in tumor growth and tumorigenicity. Am. J. Pathol. 2011, 178, 1009–1020. [Google Scholar] [CrossRef]
- Alyabsi, M.; Alhumaid, A.; Allah-Bakhsh, H.; Alkelya, M.; Aziz, M.A. Colorectal cancer in Saudi Arabia as the proof-of-principle model for implementing strategies of predictive, preventive, and personalized medicine in healthcare. EPMA J. 2020, 11, 119–131. [Google Scholar] [CrossRef]
- Almutairi, M.H.; Alrubie, T.M.; Almutairi, B.O.; Alamri, A.M.; Alrefaei, A.F.; Arafah, M.M.; Alanazi, M.; Semlali, A. The Expression Patterns of Human Cancer-Testis Genes Are Induced through Epigenetic Drugs in Colon Cancer Cells. Pharmaceuticals 2022, 15, 1319. [Google Scholar] [CrossRef]
- Hofmann, O.; Caballero, O.L.; Stevenson, B.J.; Chen, Y.T.; Cohen, T.; Chua, R.; Maher, C.A.; Panji, S.; Schaefer, U.; Kruger, A.; et al. Genome-wide analysis of cancer/testis gene expression. Proc. Natl. Acad. Sci. USA 2008, 105, 20422–20427. [Google Scholar] [CrossRef]
- Almutairi, M.H.; Alrubie, T.M.; Alamri, A.M.; Almutairi, B.O.; Alrefaei, A.F.; Arafah, M.M.; Alanazi, M.; Semlali, A. Cancer-Testis Gene Biomarkers Discovered in Colon Cancer Patients. Genes 2022, 13, 807. [Google Scholar] [CrossRef]
- Babatunde, K.A.; Najafi, A.; Salehipour, P.; Modarressi, M.H.; Mobasheri, M.B. Cancer/Testis genes in relation to sperm biology and function. Iran. J. Basic Med. Sci. 2017, 20, 967–974. [Google Scholar] [CrossRef]
- Almatrafi, A.; Feichtinger, J.; Vernon, E.G.; Escobar, N.G.; Wakeman, J.A.; Larcombe, L.D.; McFarlane, R.J. Identification of a class of human cancer germline genes with transcriptional silencing refractory to the hypomethylating drug 5-aza-2′-deoxycytidine. Oncoscience 2014, 1, 745–750. [Google Scholar] [CrossRef]
- Almeida, L.G.; Sakabe, N.J.; deOliveira, A.R.; Silva, M.C.; Mundstein, A.S.; Cohen, T.; Chen, Y.T.; Chua, R.; Gurung, S.; Gnjatic, S.; et al. CTdatabase: A knowledge-base of high-throughput and curated data on cancer-testis antigens. Nucleic Acids Res. 2009, 37, D816–D819. [Google Scholar] [CrossRef]
- Gure, A.O.; Wei, I.J.; Old, L.J.; Chen, Y.T. The SSX gene family: Characterization of 9 complete genes. Int. J. Cancer 2002, 101, 448–453. [Google Scholar] [CrossRef]
- Smith, H.A.; McNeel, D.G. The SSX family of cancer-testis antigens as target proteins for tumor therapy. Clin. Dev. Immunol. 2010, 2010, 150591. [Google Scholar] [CrossRef]
- Gure, A.O.; Tureci, O.; Sahin, U.; Tsang, S.; Scanlan, M.J.; Jager, E.; Knuth, A.; Pfreundschuh, M.; Old, L.J.; Chen, Y.T. SSX: A multigene family with several members transcribed in normal testis and human cancer. Int. J. Cancer 1997, 72, 965–971. [Google Scholar]
- Tureci, O.; Chen, Y.T.; Sahin, U.; Gure, A.O.; Zwick, C.; Villena, C.; Tsang, S.; Seitz, G.; Old, L.J.; Pfreundschuh, M. Expression of SSX genes in human tumors. Int. J. Cancer 1998, 77, 19–23. [Google Scholar] [CrossRef]
- Choi, J.; Chang, H. The expression of MAGE and SSX, and correlation of COX2, VEGF, and survivin in colorectal cancer. Anticancer Res. 2012, 32, 559–564. [Google Scholar]
- Jakobsen, M.K.; Traynor, S.; Staehr, M.; Duijf, P.G.; Nielsen, A.Y.; Terp, M.G.; Pedersen, C.B.; Guldberg, P.; Ditzel, H.J.; Gjerstorff, M.F. The Cancer/Testis Antigen Gene VCX2 Is Rarely Expressed in Malignancies but Can Be Epigenetically Activated Using DNA Methyltransferase and Histone Deacetylase Inhibitors. Front. Oncol. 2020, 10, 584024. [Google Scholar] [CrossRef]
- Caponigro, F.; Basile, M.; de Rosa, V.; Normanno, N. New drugs in cancer therapy, National Tumor Institute, Naples, 17-18 June 2004. Anticancer Drugs 2005, 16, 211–221. [Google Scholar] [CrossRef]
- Feichtinger, J.; Aldeailej, I.; Anderson, R.; Almutairi, M.; Almatrafi, A.; Alsiwiehri, N.; Griffiths, K.; Stuart, N.; Wakeman, J.A.; Larcombe, L.; et al. Meta-analysis of clinical data using human meiotic genes identifies a novel cohort of highly restricted cancer-specific marker genes. Oncotarget 2012, 3, 843–853. [Google Scholar] [CrossRef]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Bruggeman, J.W.; Koster, J.; van Pelt, A.M.M.; Speijer, D.; Hamer, G. How germline genes promote malignancy in cancer cells. Bioessays 2022, 45, 2200112. [Google Scholar] [CrossRef]
- Gure, A.O.; Chua, R.; Williamson, B.; Gonen, M.; Ferrera, C.A.; Gnjatic, S.; Ritter, G.; Simpson, A.J.; Chen, Y.T.; Old, L.J.; et al. Cancer-testis genes are coordinately expressed and are markers of poor outcome in non-small cell lung cancer. Clin. Cancer Res. 2005, 11, 8055–8062. [Google Scholar] [CrossRef]
- Naka, N.; Joyama, S.; Tsukamoto, Y.; Yoshioka, K.; Hashimoto, N.; Ujiiye, T.; Hayashi, T.; Kawase, M.; Mano, M.; Ishiguro, S.; et al. Quantification of SSX mRNA expression in human bone and soft tissue tumors using nucleic acid sequence-based amplification. J. Mol. Diagn 2005, 7, 187–197. [Google Scholar] [CrossRef]
- Taylor, B.J.; Reiman, T.; Pittman, J.A.; Keats, J.J.; de Bruijn, D.R.; Mant, M.J.; Belch, A.R.; Pilarski, L.M. SSX cancer testis antigens are expressed in most multiple myeloma patients: Co-expression of SSX1, 2, 4, and 5 correlates with adverse prognosis and high frequencies of SSX-positive PCs. J. Immunother. 2005, 28, 564–575. [Google Scholar] [CrossRef]
- Smith, H.A.; Cronk, R.J.; Lang, J.M.; McNeel, D.G. Expression and immunotherapeutic targeting of the SSX family of cancer-testis antigens in prostate cancer. Cancer Res. 2011, 71, 6785–6795. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, L.Q.; Lu, Z.H.; Wang, X.J.; Yang, J.Y. Expression of SSX-1 and NY-ESO-1 mRNA in tumor tissues and its corresponding peripheral blood expression in patients with hepatocellular carcinoma. Chin. Med. J. 2007, 120, 1042–1046. [Google Scholar]
- Peng, J.R.; Chen, H.S.; Mou, D.C.; Cao, J.; Cong, X.; Qin, L.L.; Wei, L.; Leng, X.S.; Wang, Y.; Chen, W.F. Expression of cancer/testis (CT) antigens in Chinese hepatocellular carcinoma and its correlation with clinical parameters. Cancer Lett. 2005, 219, 223–232. [Google Scholar] [CrossRef]
- Lungulescu, C.; Ghimpau, V.; Gheonea, D.I.; Sur, D.; Lungulescu, C.V. The Role of Neuropilin-2 in the Epithelial to Mesenchymal Transition of Colorectal Cancer: A Systematic Review. Biomedicines 2022, 10, 172. [Google Scholar] [CrossRef]
- Sanchez-Diez, M.; Alegria-Aravena, N.; Lopez-Montes, M.; Quiroz-Troncoso, J.; Gonzalez-Martos, R.; Menendez-Rey, A.; Sanchez-Sanchez, J.L.; Pastor, J.M.; Ramirez-Castillejo, C. Implication of Different Tumor Biomarkers in Drug Resistance and Invasiveness in Primary and Metastatic Colorectal Cancer Cell Lines. Biomedicines 2022, 10, 1083. [Google Scholar] [CrossRef]
- Niemeyer, P.; Tureci, O.; Eberle, T.; Graf, N.; Pfreundschuh, M.; Sahin, U. Expression of serologically identified tumor antigens in acute leukemias. Leuk. Res. 2003, 27, 655–660. [Google Scholar] [CrossRef]
- Caballero, O.L.; Chen, Y.T. Cancer/testis (CT) antigens: Potential targets for immunotherapy. Cancer Sci. 2009, 100, 2014–2021. [Google Scholar] [CrossRef]
- He, L.; Ji, J.N.; Liu, S.Q.; Xue, E.; Liang, Q.; Ma, Z. Expression of cancer-testis antigen in multiple myeloma. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 181–185. [Google Scholar] [CrossRef]
- Cannuyer, J.; Loriot, A.; Parvizi, G.K.; De Smet, C. Epigenetic hierarchy within the MAGEA1 cancer-germline gene: Promoter DNA methylation dictates local histone modifications. PLoS ONE 2013, 8, e58743. [Google Scholar] [CrossRef]
- Kim, R.; Kulkarni, P.; Hannenhalli, S. Derepression of Cancer/testis antigens in cancer is associated with distinct patterns of DNA hypomethylation. BMC Cancer 2013, 13, 144. [Google Scholar] [CrossRef]
- Tamura, G.; Yin, J.; Wang, S.; Fleisher, A.S.; Zou, T.; Abraham, J.M.; Kong, D.; Smolinski, K.N.; Wilson, K.T.; James, S.P.; et al. E-Cadherin gene promoter hypermethylation in primary human gastric carcinomas. J. Natl. Cancer Inst. 2000, 92, 569–573. [Google Scholar] [CrossRef]
- Lu, W.; Kang, Y. Epithelial-Mesenchymal Plasticity in Cancer Progression and Metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef]
- Vatolin, S.; Abdullaev, Z.; Pack, S.D.; Flanagan, P.T.; Custer, M.; Loukinov, D.I.; Pugacheva, E.; Hong, J.A.; Morse, H., 3rd; Schrump, D.S.; et al. Conditional expression of the CTCF-paralogous transcriptional factor BORIS in normal cells results in demethylation and derepression of MAGE-A1 and reactivation of other cancer-testis genes. Cancer Res. 2005, 65, 7751–7762. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, J.; Fang, M.; Wang, X. Investigation of fusion gene expression in HCT116 cells. Oncol. Lett. 2017, 14, 6962–6968. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Liang, Q.; Xu, Y.; Sang, J. MAGEA1 inhibits the expression of BORIS via increased promoter methylation. J. Cell Sci. 2019, 132, jcs218628. [Google Scholar] [CrossRef]
- Chen, J.; Wu, L.; Xu, H.; Cheng, S. 5-Aza-Cdr regulates RASSF1a by inhibiting DNMT1 to affect colon cancer cell proliferation, migration and apoptosis. Cancer Manag. Res. 2019, 11, 9517. [Google Scholar] [CrossRef]
- Yu, J.; Qin, B.; Moyer, A.M.; Nowsheen, S.; Liu, T.; Qin, S.; Zhuang, Y.; Liu, D.; Lu, S.W.; Kalari, K.R.; et al. DNA methyltransferase expression in triple-negative breast cancer predicts sensitivity to decitabine. J. Clin. Investig. 2018, 128, 2376–2388. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Accession Number | Category | Sequence (5′→3′) | Ta * | Product Size (bp) |
|---|---|---|---|---|---|
| ACTB | NM_001101.5 | Forward Reverse | AGAAAATCTGGCACCACACC AGGAAGGAAGGCTGGAAGAG | 58 °C | 553 |
| SSX1 | NM_001278691.2 | Forward Reverse | CCAGGGATGATGCTAAAGCA GTGCAGTTGTTTCCCATCGT | 58 °C | 389 |
| SSX2 | NM_175698.4 | Forward Reverse | CAGAGAAGATCCAAAAGGCC CTCGTGAATCTTCTCAGAGG | 58 °C | 407 |
| SSX3 | NM_021014.4 | Forward Reverse | CACGGTTGGTGCTCAAATAC TCATCTTCCTCAGGATCGCT | 58 °C | 528 |
| Primer | Category | Sequence (5′→3′) | Ta * | Product Size (bp) |
|---|---|---|---|---|
| GAPDH | Forward Reverse | GGGAAGCTTGTCATCAATGG GAGATGATGACCCTTTTGGC | 58 °C | 173 |
| SSX1 | Forward Reverse | ACCATAACCGCAGGATTCAG GTGCAGTTGTTTCCCATCGT | 58 °C | 161 |
| SSX2 | Forward Reverse | ACGTCCTCAGATGACTTTCG CTCGTGAATCTTCTCAGAGG | 58 °C | 175 |
| SSX3 | Forward Reverse | AACGATGGGAAACAGCTGTG TCATCTTCCTCAGGATCGCT | 58 °C | 161 |
| Variables | CC (N%) | NC (N%) | BC (N%) | CLL (N%) | NB (N%) |
|---|---|---|---|---|---|
| Participants | 35 (100%) | 35 (100%) | 15 (100%) | 12 (100%) | 12 (100%) |
| Sex | |||||
| Males | 30 (86%) | 30 (86%) | ----- | 12 (100%) | 12 (100%) |
| Females | 5 (14%) | 5 (14%) | 15 (100%) | ----- | ----- |
| Mean age (min–max) | 61 (24–96) | 61 (24–96) | 52 (32–74) | 49 (39–64) | 43 (33–52) |
| Below 61 | 15 (43%) | 15 (43%) | ----- | ----- | ----- |
| Above 61 | 20 (57%) | 20 (57%) | ----- | ----- | ----- |
| Below 52 | ----- | ----- | 9 (60%) | ----- | ----- |
| Above 52 | ----- | ----- | 6 (40%) | ----- | ----- |
| Below 49 | ----- | ----- | ----- | 6 (50%) | ----- |
| Above 49 | ----- | ----- | ----- | 6 (50%) | ----- |
| Below 43 | ----- | ----- | ----- | ----- | 8 (67%) |
| Above 43 | ----- | ----- | ----- | ----- | 4 (33%) |
| ER status | |||||
| ER+ | ----- | ----- | 6 (40%) | ----- | ----- |
| ER− | ----- | ----- | 9 (60%) | ----- | ----- |
| PR status | |||||
| PR+ | ----- | ----- | 6 (40%) | ----- | ----- |
| PR− | ----- | ----- | 9 (60%) | ----- | ----- |
| The cancer grades of CC patients | |||||
| Grade | I | II | III | ||
| CC patients | Males (4, 21–23, 27–29) | Males (3, 5–10, 16–20, 24–26, 30) Females (1–5) | Males (1–2, 11–15) | ||
| The cancer grades of BC patients | |||||
| Grade | I | II | III | ||
| BC patients | 7, 11, 14 | 1–3, 6, 8, 12–13, 15 | 4–5, 9–10 | ||
| SSX Genes | Positive Expression Number in 30 NC Samples (%) | Positive Expression Number in 30 CC Samples (%) | p Value |
|---|---|---|---|
| SSX1 | 0 (0%) | 3 (10%) | 0.078 |
| SSX2 | 0 (0%) | 6 (20%) | 0.009 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alrubie, T.M.; Alamri, A.M.; Almutairi, B.O.; Alrefaei, A.F.; Arafah, M.M.; Alanazi, M.; Semlali, A.; Almutairi, M.H. Higher Expression Levels of SSX1 and SSX2 in Patients with Colon Cancer: Regulated In Vitro by the Inhibition of Methylation and Histone Deacetylation. Medicina 2023, 59, 988. https://doi.org/10.3390/medicina59050988
Alrubie TM, Alamri AM, Almutairi BO, Alrefaei AF, Arafah MM, Alanazi M, Semlali A, Almutairi MH. Higher Expression Levels of SSX1 and SSX2 in Patients with Colon Cancer: Regulated In Vitro by the Inhibition of Methylation and Histone Deacetylation. Medicina. 2023; 59(5):988. https://doi.org/10.3390/medicina59050988
Chicago/Turabian StyleAlrubie, Turki M., Abdullah M. Alamri, Bader O. Almutairi, Abdulwahed F. Alrefaei, Maha M. Arafah, Mohammad Alanazi, Abdelhabib Semlali, and Mikhlid H. Almutairi. 2023. "Higher Expression Levels of SSX1 and SSX2 in Patients with Colon Cancer: Regulated In Vitro by the Inhibition of Methylation and Histone Deacetylation" Medicina 59, no. 5: 988. https://doi.org/10.3390/medicina59050988