
Comprehensive Investigation Illustrates the Role of M2 Macrophages and Its Related Genes in Pancreatic Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
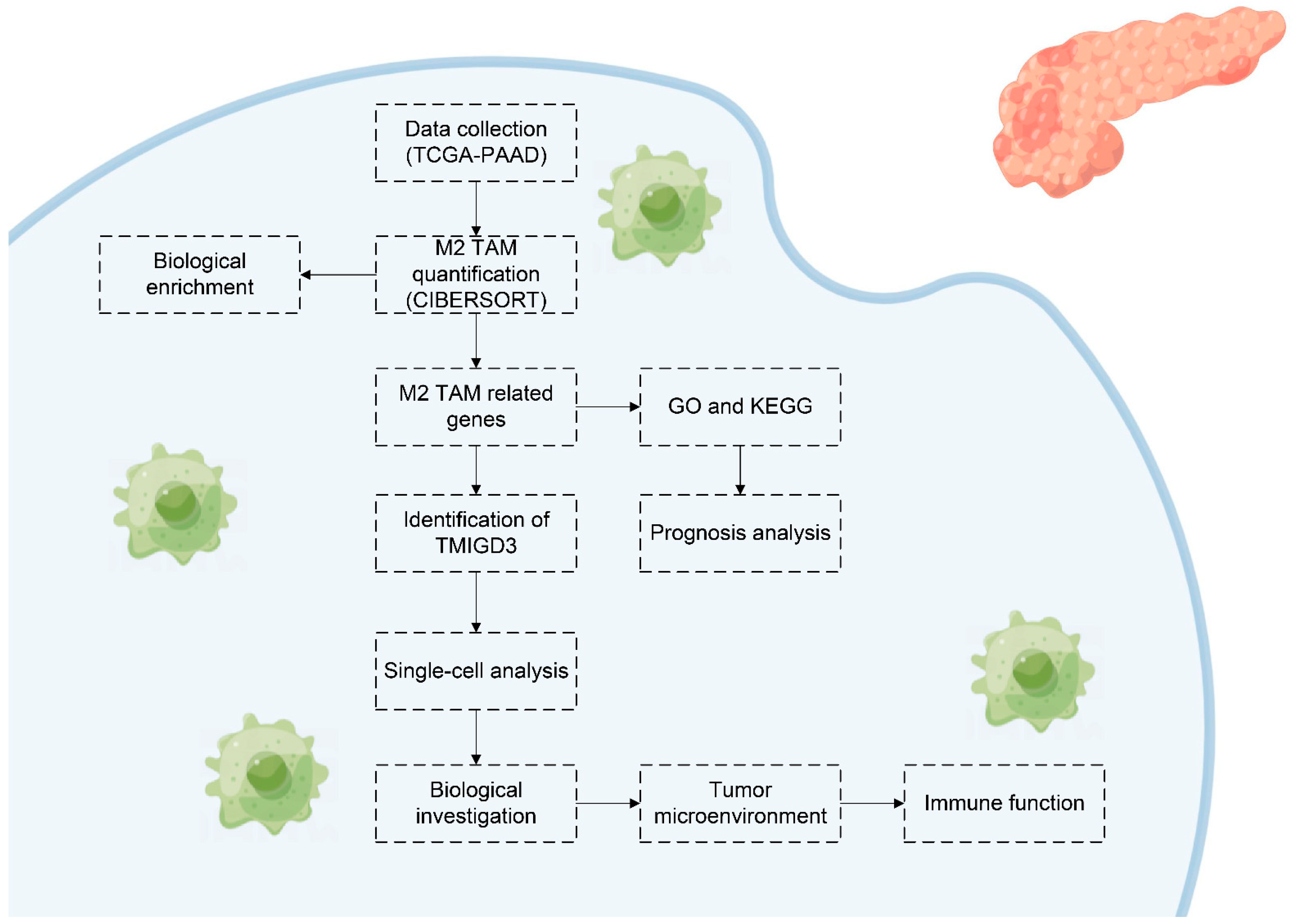
2. Methods
2.1. Acquisition and Pre-Processing of Open-Access Data
2.2. Assessment of M2 Macrophages in the PC Microenvironment
2.3. Correlation Analysis
2.4. Biological Investigation
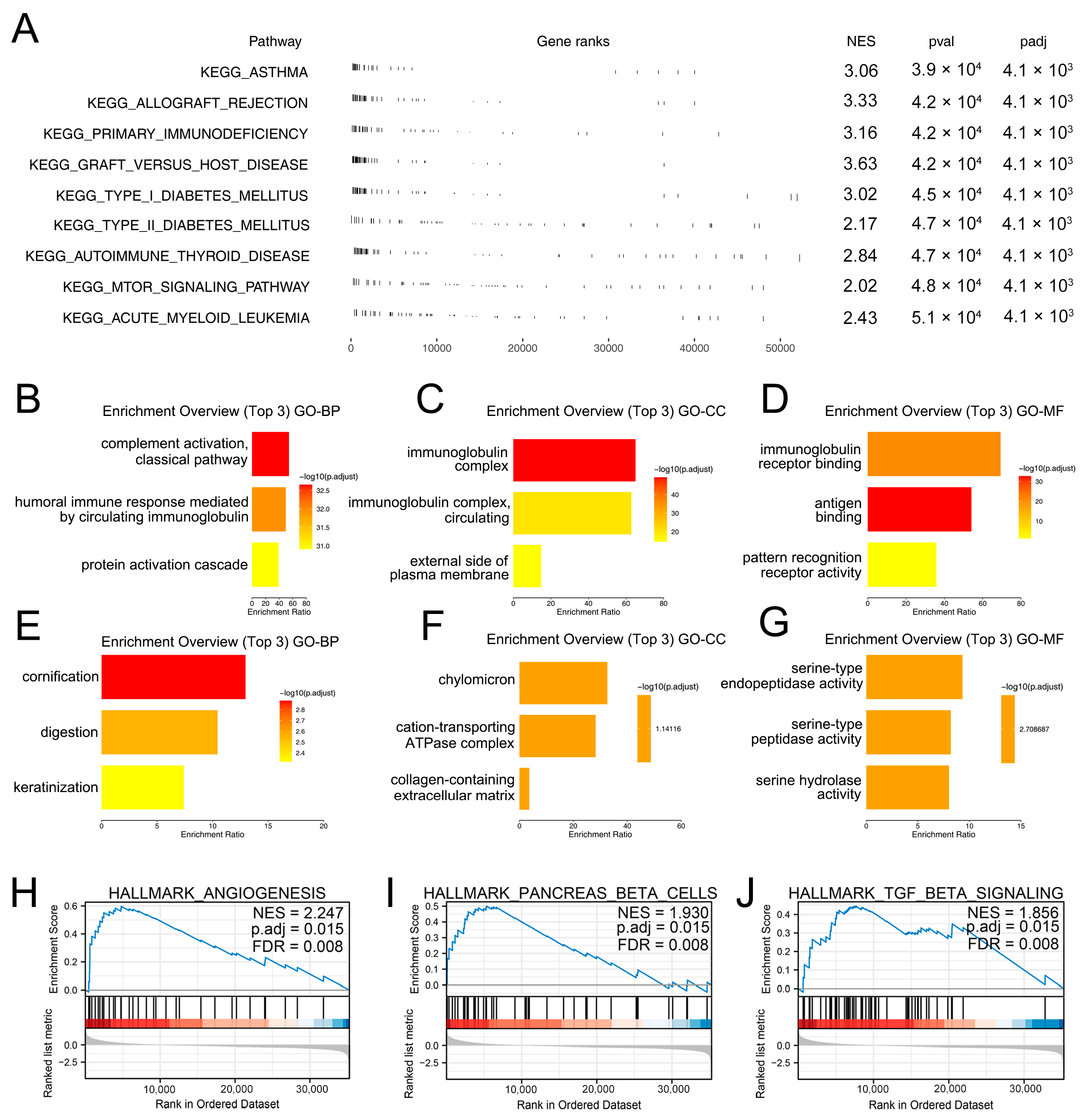
2.5. Gene Set Enrichment Analysis (GSEA)
2.6. Single-Cell Analysis
2.7. Tumor Microenvironment
2.8. Statistical Analysis
3. Results
3.1. The Biological Effect of M2 Macrophages in PC
3.2. Identification of the M2 Macrophages-Related Genes
3.3. Expression Pattern of TMIGD3 in PC
3.4. Biological Role of TMIGD3 in PC
3.5. Effect of TMIGD3 on PC Microenvironment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klein, A.P. Pancreatic Cancer Epidemiology: Understanding the Role of Lifestyle and Inherited Risk Factors. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Park, W.; Chawla, A.; O’Reilly, E.M. Pancreatic Cancer: A Review. JAMA 2021, 326, 851–862. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Mehla, K.; Singh, P.K. Metabolic Regulation of Macrophage Polarization in Cancer. Trends Cancer 2019, 5, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, S.; Wang, Q.; Zhang, X. Tumor-Recruited M2 Macrophages Promote Gastric and Breast Cancer Metastasis Via M2 Macrophage-Secreted Chi3l1 Protein. J. Hematol. Oncol. 2017, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Yin, L.; Ouyang, X.; Zeng, K.; Xiao, Y.; Li, Y. M2 Macrophages Promote Hcc Cells Invasion and Migration Via Mir-149-5p/Mmp9 Signaling. J. Cancer 2020, 11, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Chen, Z.; Zhong, Q.; Zheng, Z.; Chen, Y.; Shangguan, W.; Zhang, Y.; Yang, J.; Zhu, D.; Xie, W. M2 Macrophages Secrete Cxcl13 to Promote Renal Cell Carcinoma Migration, Invasion, and Emt. Cancer Cell Int. 2021, 21, 677. [Google Scholar] [CrossRef]
- Vidyarthi, A.; Agnihotri, T.; Khan, N.; Singh, S.; Tewari, M.K.; Radotra, B.D.; Chatterjee, D.; Agrewala, J.N. Predominance of M2 Macrophages in Gliomas Leads to the Suppression of Local and Systemic Immunity. Cancer Immunol. Immunother. 2019, 68, 1995–2004. [Google Scholar] [CrossRef]
- An, Y.; Yang, Q. Mir-21 Modulates the Polarization of Macrophages and Increases the Effects of M2 Macrophages on Promoting the Chemoresistance of Ovarian Cancer. Life Sci. 2020, 242, 117162. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for Rna-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Khodadoust, M.S.; Liu, C.L.; Newman, A.M.; Alizadeh, A.A. Profiling Tumor Infiltrating Immune Cells with Cibersort. Methods Mol. Biol. 2018, 1711, 243–259. [Google Scholar] [PubMed]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An R Package for Comparing Biological Themes among Gene Clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Wang, J.; Han, Y.; Dong, X.; Ge, J.; Zheng, R.; Shi, X.; Wang, B.; Li, Z.; Ren, P.; et al. Tisch: A Comprehensive Web Resource Enabling Interactive Single-Cell Transcriptome Visualization of Tumor Microenvironment. Nucleic Acids Res. 2021, 49, D1420–D1430. [Google Scholar] [CrossRef]
- Plattner, C.; Finotello, F.; Rieder, D. Deconvoluting Tumor-Infiltrating Immune Cells from Rna-Seq Data Using Quantiseq. Methods Enzymol. 2020, 636, 261–285. [Google Scholar]
- Aran, D.; Hu, Z.; Butte, A.J. Xcell: Digitally Portraying the Tissue Cellular Heterogeneity Landscape. Genome Biol. 2017, 18, 220. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. Timer: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Racle, J.; Gfeller, D. Epic: A Tool to Estimate the Proportions of Different Cell Types from Bulk Gene Expression Data. Methods Mol. Biol. 2020, 2120, 233–248. [Google Scholar]
- Hänzelmann, S.; Castelo, R.; Guinney, J. Gsva: Gene Set Variation Analysis for Microarray and Rna-Seq Data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Pourshams, A.; Sepanlou, S.G.; Ikuta, K.S.; Bisignano, C.; Safiri, S.; Roshandel, G.; Sharif, M.; Khatibian, M.; Fitzmaurice, C.; Nixon, M.R.; et al. The Global, Regional, and National Burden of Pancreatic Cancer and Its Attributable Risk Factors in 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2019, 4, 934–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viale, P.H. The American Cancer Society’s Facts & Figures: 2020 Edition. J. Adv. Pract. Oncol. 2020, 11, 135–136. [Google Scholar]
- Bailey, P.; Chang, D.K.; Nones, K.; Johns, A.L.; Patch, A.M.; Gingras, M.C.; Miller, D.K.; Christ, A.N.; Bruxner, T.J.; Quinn, M.C.; et al. Genomic Analyses Identify Molecular Subtypes of Pancreatic Cancer. Nature 2016, 531, 47–52. [Google Scholar] [CrossRef]
- Vincent, A.; Herman, J.; Schulick, R.; Hruban, R.H.; Goggins, M. Pancreatic Cancer. Lancet 2011, 378, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, D.; Seo, N.; Hayashi, T.; Tahara, Y.; Fujii, K.; Tawara, I.; Miyahara, Y.; Okamori, K.; Yagita, H.; Imoto, S.; et al. Antigen Delivery Targeted to Tumor-Associated Macrophages Overcomes Tumor Immune Resistance. J. Clin. Investig. 2019, 129, 1278–1294. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-Associated Macrophages in Tumor Metastasis: Biological Roles and Clinical Therapeutic Applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- Wang, H.; Yung, M.M.H.; Ngan, H.Y.S.; Chan, K.K.L.; Chan, D.W. The Impact of the Tumor Microenvironment on Macrophage Polarization in Cancer Metastatic Progression. Int. J. Mol. Sci. 2021, 22, 6560. [Google Scholar] [CrossRef] [PubMed]
- Marigo, I.; Trovato, R.; Hofer, F.; Ingangi, V.; Desantis, G.; Leone, K.; De Sanctis, F.; Ugel, S.; Canè, S.; Simonelli, A.; et al. Disabled Homolog 2 Controls Prometastatic Activity of Tumor-Associated Macrophages. Cancer Discov. 2020, 10, 1758–1773. [Google Scholar] [CrossRef]
- Iyer, S.V.; Ranjan, A.; Elias, H.K.; Parrales, A.; Sasaki, H.; Roy, B.C.; Umar, S.; Tawfik, O.W.; Iwakuma, T. Genome-Wide Rnai Screening Identifies Tmigd3 Isoform1 as a Suppressor of Nf-Κb and Osteosarcoma Progression. Nat. Commun. 2016, 7, 13561. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ji, S.; Zhang, B.; Liu, J.; Qin, Y.; Xu, J.; Yu, X. Role of Angiogenesis in Pancreatic Cancer Biology and Therapy. Biomed. Pharmacother. 2018, 108, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Amsterdam, A.; Raanan, C.; Schreiber, L.; Polin, N.; Givol, D. Lgr5 and Nanog Identify Stem Cell Signature of Pancreas Beta Cells Which Initiate Pancreatic Cancer. Biochem. Biophys. Res. Commun. 2013, 433, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Gough, N.R.; Xiang, X.; Mishra, L. Tgf-Β Signaling in Liver, Pancreas, and Gastrointestinal Diseases and Cancer. Gastroenterology 2021, 161, 434–452.e15. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.H.; Wang, X.K. Microenvironment-Related Prognostic Genes in Esophageal Cancer. Transl. Cancer Res. 2020, 9, 7531–7539. [Google Scholar] [CrossRef]
- Juusola, M.; Kuuliala, K.; Kuuliala, A.; Mustonen, H.; Vähä-Koskela, M.; Puolakkainen, P.; Seppänen, H. Pancreatic Cancer Is Associated with Aberrant Monocyte Function and Successive Differentiation into Macrophages with Inferior Anti-Tumour Characteristics. Pancreatology 2021, 21, 397–405. [Google Scholar] [CrossRef]
- Padoan, A.; Plebani, M.; Basso, D. Inflammation and Pancreatic Cancer: Focus on Metabolism, Cytokines, and Immunity. Int. J. Mol. Sci. 2019, 20, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.; Noel, P.; Borazanci, E.H.; Lee, J.; Amini, A.; Han, I.W.; Heo, J.S.; Jameson, G.S.; Fraser, C.; Steinbach, M.; et al. Single-Cell Transcriptome Analysis of Tumor and Stromal Compartments of Pancreatic Ductal Adenocarcinoma Primary Tumors and Metastatic Lesions. Genome Med. 2020, 12, 80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ren, X.; Zhang, T.; Zhou, X.; Chen, X.; Lu, H.; Zhou, X.; Zhang, X.; Wang, S.; Qin, C. Comprehensive Analysis of the Association between Human Non-Obstructive Azoospermia and Plasticisers Via Single-Cell and Traditional Rna Sequencing Methods. Expo. Health 2022, 14, 829–842. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, X.; Zhang, X.; Ren, X.; Wu, J.; Wang, Z.; Wang, S.; Wang, Z. Gut Microbiota May Contribute to the Postnatal Male Reproductive Abnormalities Induced by Prenatal Dibutyl Phthalate Exposure. Chemosphere 2022, 287 Pt 1, 132046. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, T.; Ren, X.; Chen, X.; Wang, S.; Qin, C. Pyrethroids Toxicity to Male Reproductive System and Offspring as a Function of Oxidative Stress Induction: Rodent Studies. Front. Endocrinol. 2021, 12, 656106. [Google Scholar] [CrossRef]
- Wu, D.; Yin, Z.; Ji, Y.; Li, L.; Li, Y.; Meng, F.; Ren, X.; Xu, M. Identification of Novel Autophagy-Related Lncrnas Associated with a Poor Prognosis of Colon Adenocarcinoma through Bioinformatics Analysis. Sci. Rep. 2021, 11, 8069. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Tang, W.; Shen, X.; Weng, S.; Dong, L. Comprehensive Investigation Illustrates the Role of M2 Macrophages and Its Related Genes in Pancreatic Cancer. Medicina 2023, 59, 717. https://doi.org/10.3390/medicina59040717
Zhang D, Tang W, Shen X, Weng S, Dong L. Comprehensive Investigation Illustrates the Role of M2 Macrophages and Its Related Genes in Pancreatic Cancer. Medicina. 2023; 59(4):717. https://doi.org/10.3390/medicina59040717
Chicago/Turabian StyleZhang, Danying, Wenqing Tang, Xizhong Shen, Shuqiang Weng, and Ling Dong. 2023. "Comprehensive Investigation Illustrates the Role of M2 Macrophages and Its Related Genes in Pancreatic Cancer" Medicina 59, no. 4: 717. https://doi.org/10.3390/medicina59040717