
Distributions of Platelet-Derived Growth Factor Receptor-α Positive Cells and Interstitial Cells of Cajal in the Colon of Rats with Diabetes Mellitus Type 2

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
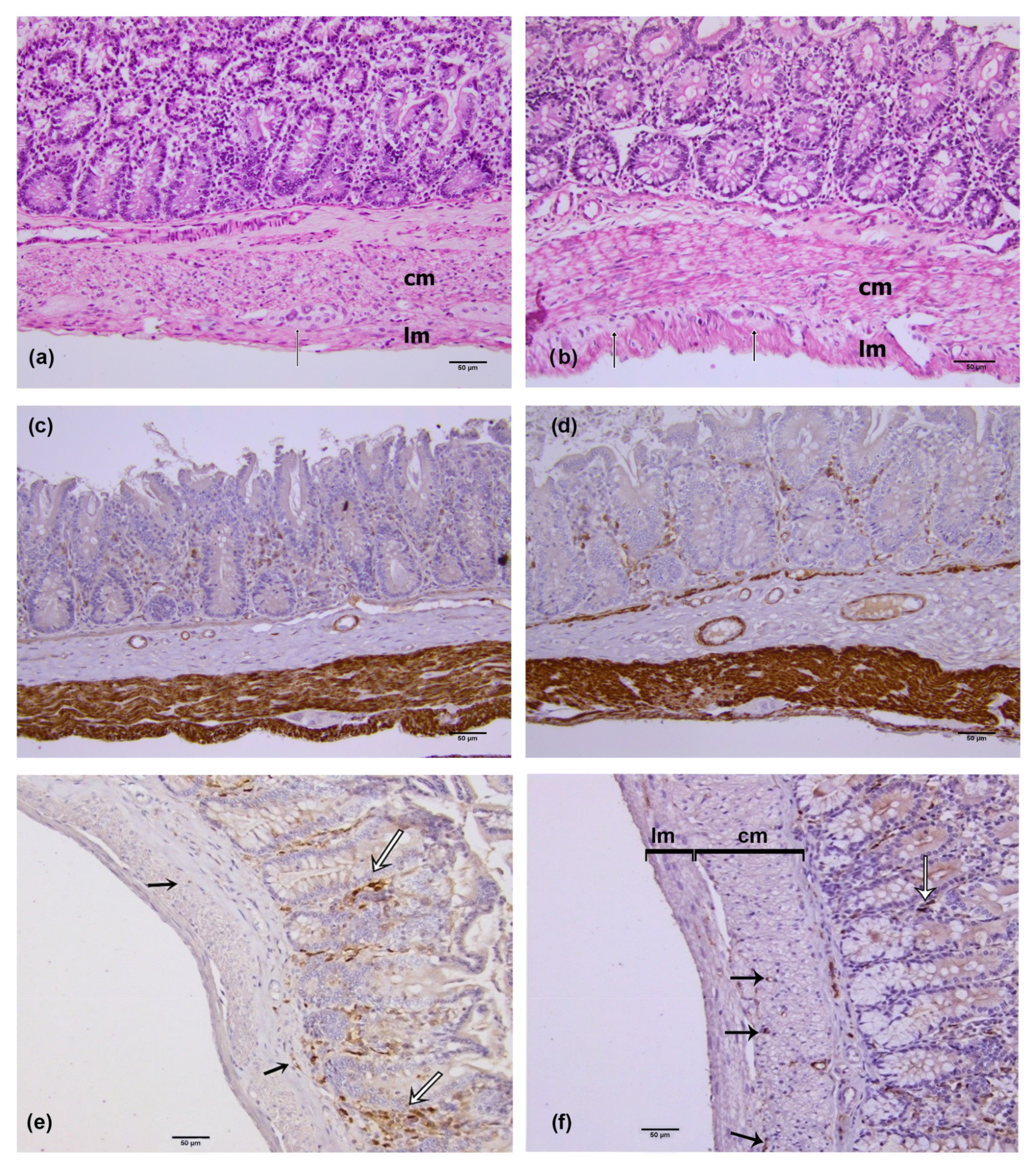
2.2. Hematoxylin and Eosin (he) Staining Method
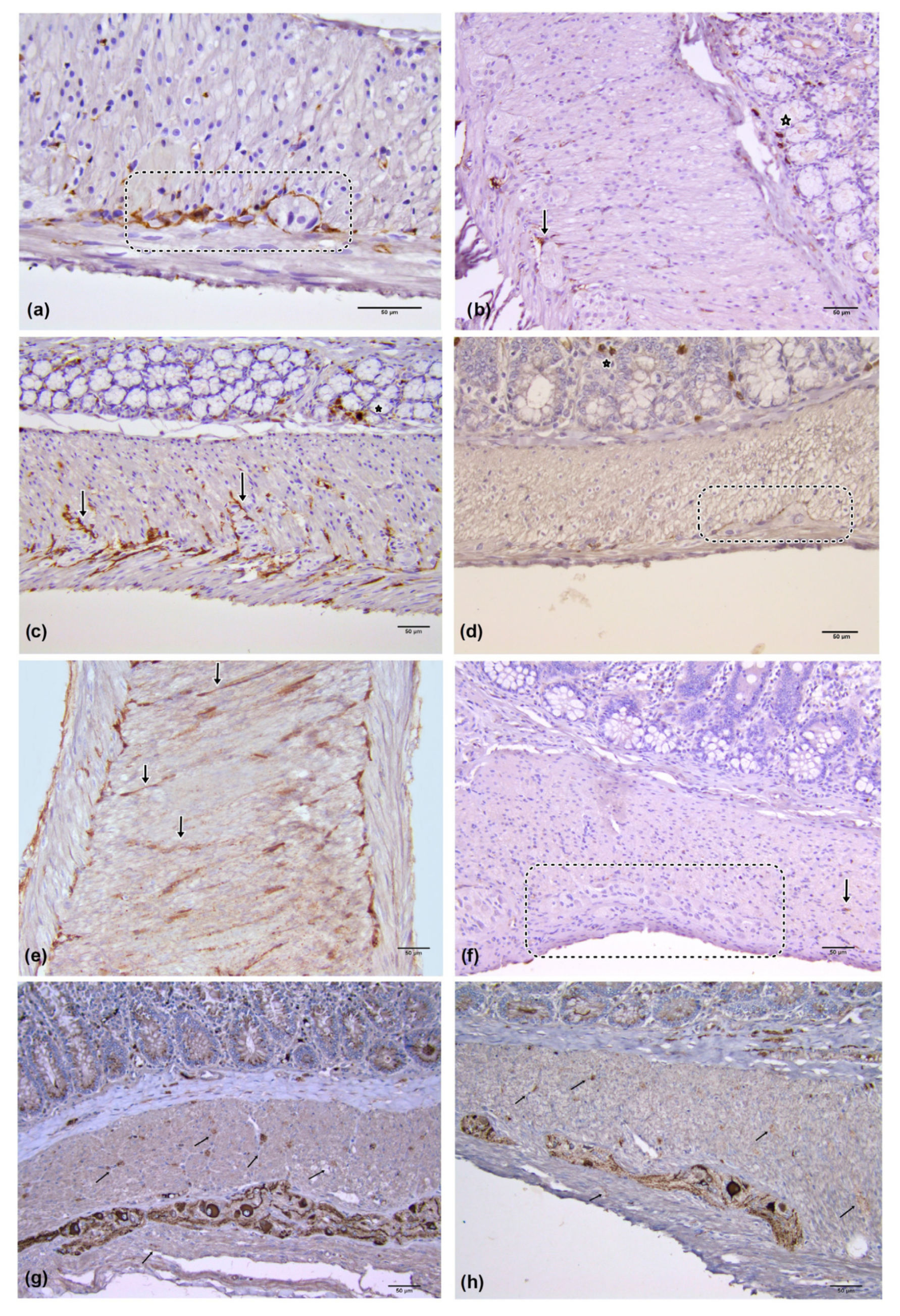
2.3. Immunohistochemistry
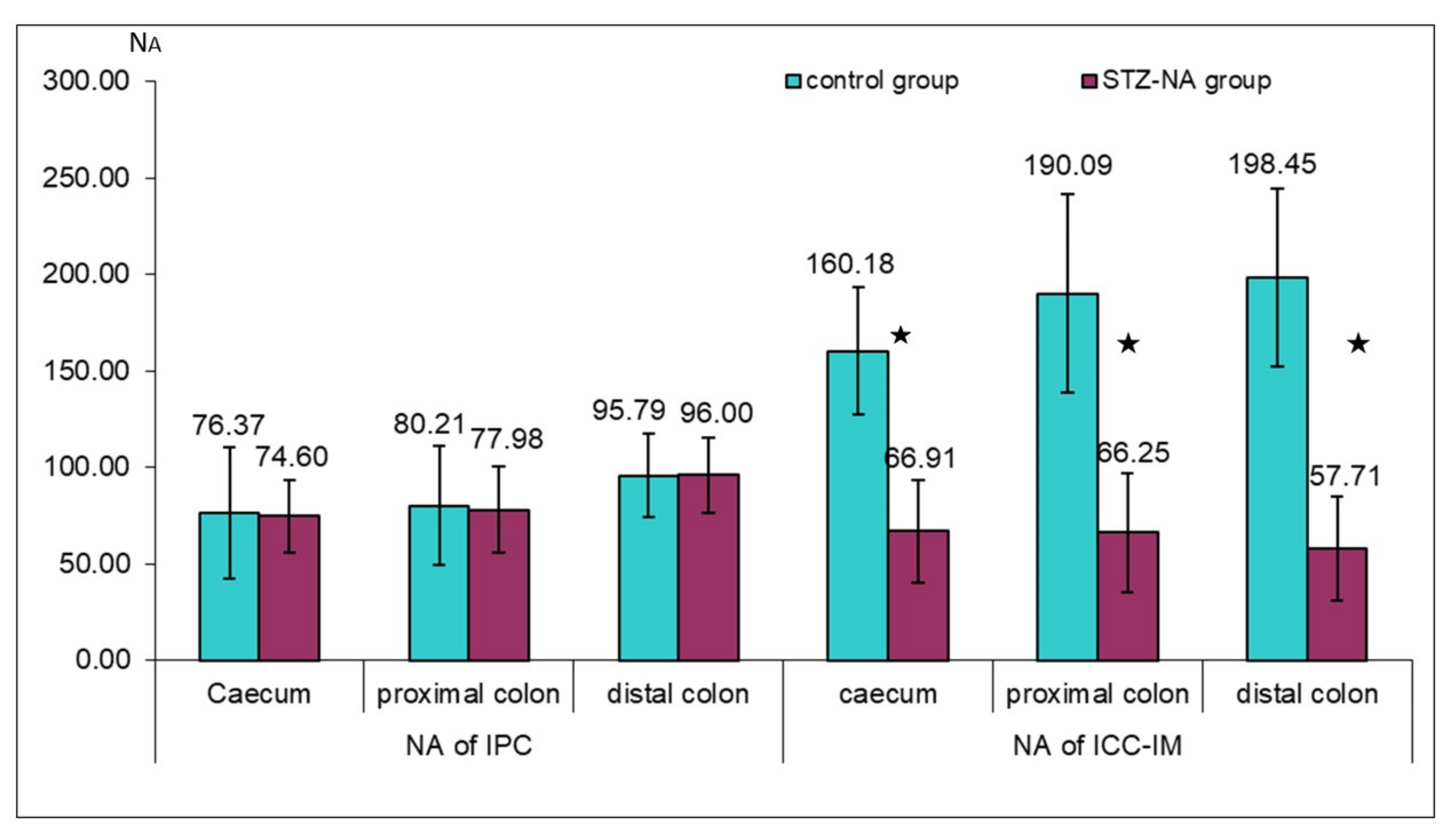
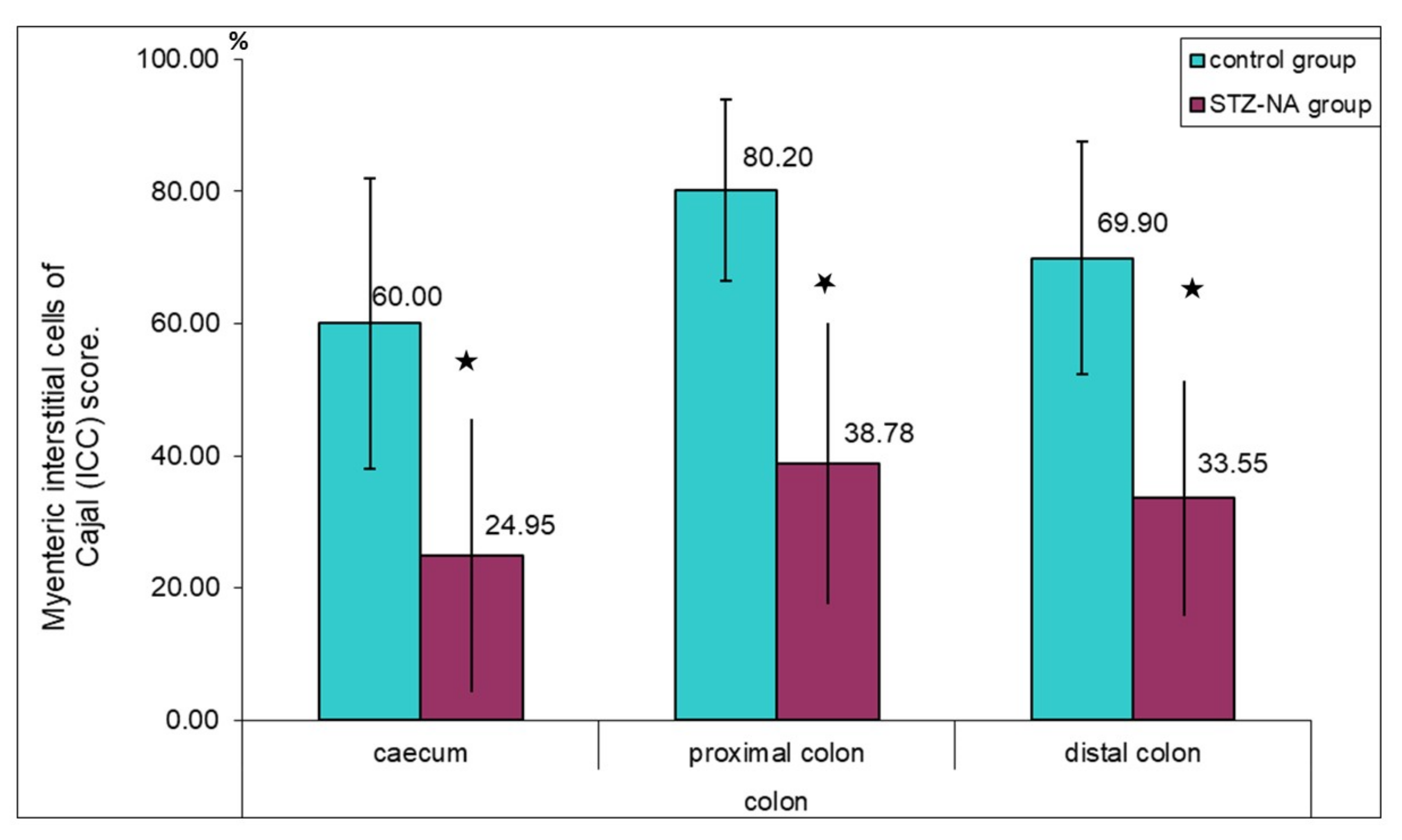
2.4. Quantitative Image Analysis
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krishnan, B.; Babu, S.; Walker, J.; Walker, A.B.; Pappachan, J.M. Gastrointestinal complications of diabetes mellitus. World J. Diabetes 2013, 4, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, M.; Kushiyama, A.; Yamazaki, H.; Kaneko, S.; Koketsu, Y.; Yamamotoya, T.; Kikuchi, T.; Sakoda, H.; Suzuki, R.; Kadowaki, T. Gastrointestinal symptom prevalence depends on disease duration and gastrointestinal region in type 2 diabetes mellitus. World J. Gastroenterol. 2017, 23, 6694–6704. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Ward, S.M.; Koh, S.D. Interstitial cells: Regulators of smooth muscle function. Physiol. Rev. 2014, 94, 859–907. [Google Scholar] [CrossRef] [PubMed]
- Komuro, T. Structure and organization of interstitial cells of Cajal in the gastrointestinal tract. J. Physiol. 2006, 576, 653–658. [Google Scholar] [CrossRef]
- Pieri, L.; Vannucchi, M.G.; Faussone-Pellegrini, M.S. Histochemical and ultrastructural characteristics of an interstitial cell type different from ICC and resident in the muscle coat of human gut. J. Cell. Mol. Med. 2008, 12, 1944–1955. [Google Scholar] [CrossRef]
- Ward, S.M.; Beckett, E.A.; Wang, X.; Baker, F.; Khoyi, M.; Sanders, K.M. Interstitial cells of Cajal mediate cholinergic neurotransmission from enteric motor neurons. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 1393–1403. [Google Scholar] [CrossRef]
- Fredriksson, L.; Li, H.; Eriksson, U. The PDGF family: Four gene products form five dimeric isoforms. Cytokine Growth Factor Rev. 2004, 15, 197–204. [Google Scholar] [CrossRef]
- Hoch, R.V.; Soriano, P. Roles of PDGF in animal development. Development 2003, 130, 4769–4784. [Google Scholar] [CrossRef]
- Andrae, J.; Gallini, R.; Betsholtz, C. Role of platelet-derived growth factors in physiology and medicine. Genes Dev. 2008, 22, 1276–1312. [Google Scholar] [CrossRef]
- Iino, S.; Horiguchi, K.; Horiguchi, S.; Nojyo, Y. c-Kit-negative fibroblast-like cells express platelet-derived growth factor receptor alpha in the murine gastrointestinal musculature. Histochem. Cell Biol. 2009, 131, 691–702. [Google Scholar] [CrossRef]
- Kurahashi, M.; Baker, S.A.; Kito, Y.; Bartlett, A.; Hara, M.; Takeyama, H.; Hashitani, H.; Sanders, K.M. PDGFRα+ Interstitial Cells are Effector Cells of PACAP Signaling in Mouse and Human Colon. Cell. Mol. Gastroenterol. Hepatol. 2022, 14, 357–373. [Google Scholar] [CrossRef]
- Kurahashi, M.; Nakano, Y.; Hennig, G.W.; Ward, S.M.; Sanders, K.M. Platelet-derived growth factor receptor α-positive cells in the tunica muscularis of human colon. J. Cell. Mol. Med. 2012, 16, 1397. [Google Scholar] [CrossRef]
- Grover, M.; Bernard, C.E.; Pasricha, P.J.; Parkman, H.P.; Abell, T.L.; Nguyen, L.A.; Snape, W.; Shen, K.R.; Sarr, M.; Swain, J.; et al. Platelet-derived growth factor receptor α (PDGFRα)-expressing “fibroblast-like cells” in diabetic and idiopathic gastroparesis of humans. Neurogastroenterol. Motil. 2012, 24, 844–852. [Google Scholar] [CrossRef]
- Blair, P.J.; Bayguinov, Y.; Sanders, K.M.; Ward, S.M. Relationship between enteric neurons and interstitial cells in the primate gastrointestinal tract. Neurogastroenterol. Motil. 2012, 24, e437. [Google Scholar] [CrossRef]
- Ashman, L.K. The biology of stem cell factor and its receptor C-kit. Int. J. Biochem. Cell Biol. 1999, 31, 1037–1051. [Google Scholar] [CrossRef]
- Hulzinga, J.D.; Thuneberg, L.; Klüppel, M.; Malysz, J.; Mikkelsen, H.B.; Bernstein, A. W/kit gene required for interstitial cells of Cajal and for intestinal pacemaker activity. Nature 1995, 373, 347–349. [Google Scholar] [CrossRef]
- Komuro, T. Atlas of Interstitial Cells of Cajal in the Gastrointestinal Tract; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Faussone-Pellegrini, M.-S.; Cortesini, C.; Romagnoli, P. The ultrastructure of the muscle coat of human gastro-oesophageal junction, with special reference to “interstitial cells of Cajal”. Front. Neurosci. 2013, 7, 49. [Google Scholar] [CrossRef]
- Maeda, H.; Yamagata, A.; Nishikawa, S.; Yoshinaga, K.; Kobayashi, S.; Nishi, K.; Nishikawa, S. Requirement of c-kit for development of intestinal pacemaker system. Development 1992, 116, 369–375. [Google Scholar] [CrossRef]
- Hwang, S.J.; Blair, P.J.A.; Britton, F.C.; O’Driscoll, K.E.; Hennig, G.; Bayguinov, Y.R.; Rock, J.R.; Harfe, B.D.; Sanders, K.M.; Ward, S.M. Expression of anoctamin 1/TMEM16A by interstitial cells of Cajal is fundamental for slow wave activity in gastrointestinal muscles. J. Physiol. 2009, 587, 4887–4904. [Google Scholar] [CrossRef]
- Huizinga, J.D.; Zarate, N.; Farrugia, G. Physiology, injury, and recovery of interstitial cells of Cajal: Basic and clinical science. Gastroenterology 2009, 137, 1548–1556. [Google Scholar] [CrossRef] [Green Version]
- Ward, S.M.; McLaren, G.J.; Sanders, K.M. Interstitial cells of Cajal in the deep muscular plexus mediate enteric motor neurotransmission in the mouse small intestine. J. Physiol. 2006, 573, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Kito, Y. The functional role of intramuscular interstitial cells of Cajal in the stomach. J. Smooth Muscle Res. 2011, 47, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D.; Reed, D.E.; Berezin, I.; Wang, X.-Y.; Valdez, D.T.; Liu, L.W.C.; Diamant, N.E. Survival dependency of intramuscular ICC on vagal afferent nerves in the cat esophagus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R302–R310. [Google Scholar] [CrossRef] [PubMed]
- Powley, T.L.; Wang, X.Y.; Fox, E.A.; Phillips, R.J.; Liu, L.W.C.; Huizinga, J.D. Ultrastructural evidence for communication between intramuscular vagal mechanoreceptors and interstitial cells of Cajal in the rat fundus. Neurogastroenterol. Motil. 2008, 20, 69–79. [Google Scholar] [CrossRef]
- Vanderwinden, J.M.; Rumessen, J.J. Interstitial cells of Cajal in human gut and gastrointestinal disease. Microsc. Res. Tech. 1999, 47, 344–360. [Google Scholar] [CrossRef]
- Mostafa, R.M.; Moustafa, Y.M.; Hamdy, H. Interstitial cells of Cajal, the Maestro in health and disease. World J. Gastroenterol. 2010, 16, 3239. [Google Scholar] [CrossRef]
- Huizinga, J.D.; Hussain, A.; Chen, J.-H. Interstitial cells of Cajal and human colon motility in health and disease. Am. J. Physiol. Liver Physiol. 2021, 321, G552–G575. [Google Scholar] [CrossRef]
- Masiello, P.; Broca, C.; Gross, R.; Roye, M.; Manteghetti, M.; Hillaire-Buys, D.; Novelli, M.; Ribes, G. Experimental NIDDM: Development of a new model in adult rats administered streptozotocin and nicotinamide. Diabetes 1998, 47, 224–229. [Google Scholar] [CrossRef]
- den Braber-Ymker, M.; Heijker, S.; Lammens, M.; Nagtegaal, I.D. Practical and reproducible estimation of myenteric interstitial cells of Cajal in the bowel for diagnostic purposes. Neurogastroenterol. Motil. 2016, 28, 1261–1267. [Google Scholar] [CrossRef]
- Aronson, D. Hyperglycemia and the pathobiology of diabetic complications. Adv. Cardiol. 2008, 45, 1–16. [Google Scholar]
- Szkudelski, T. Streptozotocin-nicotinamide-induced diabetes in the rat. Characteristics of the experimental model. Exp. Biol. Med. 2012, 237, 481–490. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Huizinga, J.D.; Diamond, J.; Liu, L.W.C. Loss of intramuscular and submuscular interstitial cells of Cajal and associated enteric nerves is related to decreased gastric emptying in streptozotocin-induced diabetes. Neurogastroenterol. Motil. 2009, 21, 1095-e92. [Google Scholar] [CrossRef]
- Yamamoto, T.; Watabe, K.; Nakahara, M.; Ogiyama, H.; Kiyohara, T.; Tsutsui, S.; Tamura, S.; Shinomura, Y.; Hayashi, N. Disturbed gastrointestinal motility and decreased interstitial cells of Cajal in diabetic db/db mice. J. Gastroenterol. Hepatol. 2008, 23, 660–667. [Google Scholar] [CrossRef]
- Lin, L.; Xu, L.; Zhang, W.; Ge, Y.; Tang, Y.; Zhang, H.; Li, X.-L.; Chen, J.D.Z. Roles of stem cell factor on the depletion of interstitial cells of Cajal in the colon of diabetic mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G241–G247. [Google Scholar] [CrossRef]
- Lammers, W.J.E.P.; Al-Bloushi, H.M.; Al-Eisaei, S.A.; Al-Dhaheri, F.A.; Stephen, B.; John, R.; Dhanasekaran, S.; Karam, S.M. Slow wave propagation and plasticity of interstitial cells of Cajal in the small intestine of diabetic rats. Exp. Physiol. 2011, 96, 1039–1048. [Google Scholar] [CrossRef]
- Song, N.-N.; Lu, H.-L.; Lu, C.; Tong, L.; Huang, S.-Q.; Huang, X.; Chen, J.; Kim, Y.-C.; Xu, W.-X. Diabetes-induced colonic slow transit mediated by the up-regulation of PDGFRα(+) cells/SK3 in streptozotocin-induced diabetic mice. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2018, 30, e13326. [Google Scholar] [CrossRef]
- Wang, Q.; Zang, J.; Huang, X.; Lu, H.; Xu, W.; Chen, J. Colonic Dysmotility in Murine Partial Colonic Obstruction Due to Functional Changes in Interstitial Cells. J. Neurogastroenterol. Motil. 2019, 25, 589–601. [Google Scholar] [CrossRef]
- Kurahashi, M.; Nakano, Y.; Peri, L.E.; Townsend, J.B.; Ward, S.M.; Sanders, K.M. A novel population of subepithelial platelet-derived growth factor receptor α-positive cells in the mouse and human colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G823–G834. [Google Scholar] [CrossRef]
- Chan, F.; Liu, Y.; Sun, H.; Li, X.; Shang, H.; Fan, D.; An, J.; Zhou, D. Distribution and possible role of PDGF-AA and PDGFR-alpha in the gastrointestinal tract of adult guinea pigs. Virchows Arch. 2010, 457, 381–388. [Google Scholar] [CrossRef]
- Walton, K.D.; Kolterud, A.; Czerwinski, M.J.; Bell, M.J.; Prakash, A.; Kushwaha, J.; Grosse, A.S.; Schnell, S.; Gumucio, D.L. Hedgehog-responsive mesenchymal clusters direct patterning and emergence of intestinal villi. Proc. Natl. Acad. Sci. USA 2012, 109, 15817–15822. [Google Scholar] [CrossRef]
- Iino, S.; Horiguchi, S.; Horiguchi, K.; Hashimoto, T. Interstitial cells of Cajal in W(sh)/W(sh) c-kit mutant mice. J. Smooth Muscle Res. 2020, 56, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Al-Shboul, O.A. The importance of interstitial cells of cajal in the gastrointestinal tract. Saudi J. Gastroenterol. Off. J. Saudi Gastroenterol. Assoc. 2013, 19, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Ward, S.M.; Friebe, A. CrossTalk proposal: Interstitial cells are involved and physiologically important in neuromuscular transmission in the gut. J. Physiol. 2016, 594, 1507–1509. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D.; Pervez, M.; Nirmalathasan, S.; Chen, J.-H. Characterization of haustral activity in the human colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 320, G1067–G1080. [Google Scholar] [CrossRef] [PubMed]
- Knowles, C.H.; De Giorgio, R.; Kapur, R.P.; Bruder, E.; Farrugia, G.; Geboes, K.; Gershon, M.D.; Hutson, J.; Lindberg, G.; Martin, J.E.; et al. Gastrointestinal neuromuscular pathology: Guidelines for histological techniques and reporting on behalf of the Gastro 2009 International Working Group. Acta Neuropathol. 2009, 118, 271–301. [Google Scholar] [CrossRef]
- Ordög, T. Interstitial cells of Cajal in diabetic gastroenteropathy. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2008, 20, 8–18. [Google Scholar] [CrossRef]
- Choi, K.M.; Gibbons, S.J.; Nguyen, T.V.; Stoltz, G.J.; Lurken, M.S.; Ordog, T.; Szurszewski, J.H.; Farrugia, G. Heme oxygenase-1 protects interstitial cells of Cajal from oxidative stress and reverses diabetic gastroparesis. Gastroenterology 2008, 135, 2055–2064. [Google Scholar] [CrossRef]
- Horváth, V.J.; Vittal, H.; Ordög, T. Reduced insulin and IGF-I signaling, not hyperglycemia, underlies the diabetes-associated depletion of interstitial cells of Cajal in the murine stomach. Diabetes 2005, 54, 1528–1533. [Google Scholar] [CrossRef]
- Ordög, T.; Takayama, I.; Cheung, W.K.; Ward, S.M.; Sanders, K.M. Remodeling of networks of interstitial cells of Cajal in a murine model of diabetic gastroparesis. Diabetes 2000, 49, 1731–1739. [Google Scholar] [CrossRef]
- Velickov, A.; Mitrovic, O.; Djordjevic, B.; Sokolovic, D.; Zivkovic, V.; Velickov, A.; Pantovic, V.; Poklar Urlih, N.; Radenkovic, G. The effect of bilberries on diabetes-related alterations of interstitial cells of Cajal in the lower oesophageal sphincter in rats. Histol. Histopathol. 2017, 32, 639–647. [Google Scholar]
- Lu, C.; Huang, X.; Lu, H.-L.; Liu, S.-H.; Zang, J.-Y.; Li, Y.-J.; Chen, J.; Xu, W.-X. Different distributions of interstitial cells of Cajal and platelet-derived growth factor receptor-α positive cells in colonic smooth muscle cell/interstitial cell of Cajal/platelet-derived growth factor receptor-α positive cell syncytium in mice. World J. Gastroenterol. 2018, 24, 4989–5004. [Google Scholar] [CrossRef]
- Iwasaki, H.; Kajimura, M.; Osawa, S.; Kanaoka, S.; Furuta, T.; Ikuma, M.; Hishida, A. A deficiency of gastric interstitial cells of Cajal accompanied by decreased expression of neuronal nitric oxide synthase and substance P in patients with type 2 diabetes mellitus. J. Gastroenterol. 2006, 41, 1076–1087. [Google Scholar] [CrossRef]
- Jin, B.; Ha, S.E.; Wei, L.; Singh, R.; Zogg, H.; Clemmensen, B.; Heredia, D.J.; Gould, T.W.; Sanders, K.M.; Ro, S. Colonic Motility Is Improved by the Activation of 5-HT(2B) Receptors on Interstitial Cells of Cajal in Diabetic Mice. Gastroenterology 2021, 161, 608–622.e7. [Google Scholar] [CrossRef]
- Li, Y.-J.; Ao, J.-P.; Huang, X.; Lu, H.-L.; Fu, H.-Y.; Song, N.-N.; Xu, W.-X.; Chen, J. Involvement of PAR2 in platelet-derived growth factor receptor-α-positive cell proliferation in the colon of diabetic mice. Physiol. Rep. 2021, 9, e15099. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Zarate, N.; Soderholm, J.D.; Bourgeois, J.M.; Liu, L.W.C.; Huizinga, J.D. Ultrastructural injury to interstitial cells of Cajal and communication with mast cells in Crohn’s disease. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2007, 19, 349–364. [Google Scholar] [CrossRef]
- Zarate, N.; Wang, X.Y.; Tougas, G.; Anvari, M.; Birch, D.; Mearin, F.; Malagelada, J.-R.; Huizinga, J.D. Intramuscular interstitial cells of Cajal associated with mast cells survive nitrergic nerves in achalasia. Neurogastroenterol. Motil. 2006, 18, 556–568. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Supplier | Dilution | Incubation |
|---|---|---|---|
| PDGFRα (Rabbit polyclonal anti-PDGFR alpha antibody) | Abcam (Cambridge, UK) Ab65258 | 1:100 | 1.5 h at room temperature |
| C-kit (Rabbit monoclonal anti-c-Kit antibody) | Abcam (Cambridge, UK) Ab32363 | 1:100 | 24 h overnight at 4 °C |
| NF m (Rabbit polyclonal anti Neurofilament Medium antibody) | Abcam (Cambridge, UK) Ab9034 | 1:1000 | 24 h overnight at 4 °C |
| Desmin (Monoclonal Mouse Anti-Human Clone D33) | Daco (Denmark) MO760 | 1:100 | 1.5 h at room temperature |
| Parameters | Groups | |
|---|---|---|
| Control | STZ–NA | |
| Initial body weight (g) | 351.7 ± 7.5 | 352.5 ± 14.9 |
| Final body weight (g) | 395.0 ± 12.3 | 413.7 ± 19.2 ** |
| Blood glucose levels (mmol/L) | 6.36 ± 0.01 | 12.02 ± 1.65 * |
| Serum insulin levels (pmol/L) | 217.1 ± 21.2 | 191.2 ± 16.3 ** |
| Group | Caecum | Proximal Colon | Distal Colon | |||
|---|---|---|---|---|---|---|
| Circular Muscle Layer | Longitudinal Muscle Layer | Circular Muscle Layer | Longitudinal Muscle Layer | Circular Muscle Layer | Longitudinal Muscle Layer | |
| CONTROL | 174.015 ± 25.57 | 25.786 ± 5.34 | 160.152 ± 28.41 | 23.85 ± 3.605 | 156.152 ± 26.23 | 26.11 ± 4.77 |
| STZ–NA | 178.441 ± 35.57 | 25.719 ± 5.42 | 155.786 ± 27.31 | 23.208 ± 5.12 | 155.786 ± 27.31 | 25.91 ± 4.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veličkov, A.I.; Djordjević, B.; Lazarević, M.; Veličkov, A.V.; Petrović, V.; Jović, M.; Denčić, T.; Radenković, G. Distributions of Platelet-Derived Growth Factor Receptor-α Positive Cells and Interstitial Cells of Cajal in the Colon of Rats with Diabetes Mellitus Type 2. Medicina 2023, 59, 308. https://doi.org/10.3390/medicina59020308
Veličkov AI, Djordjević B, Lazarević M, Veličkov AV, Petrović V, Jović M, Denčić T, Radenković G. Distributions of Platelet-Derived Growth Factor Receptor-α Positive Cells and Interstitial Cells of Cajal in the Colon of Rats with Diabetes Mellitus Type 2. Medicina. 2023; 59(2):308. https://doi.org/10.3390/medicina59020308
Chicago/Turabian StyleVeličkov, Aleksandra Ivana, Branka Djordjević, Milica Lazarević, Asen Veselin Veličkov, Vladimir Petrović, Marko Jović, Tijana Denčić, and Goran Radenković. 2023. "Distributions of Platelet-Derived Growth Factor Receptor-α Positive Cells and Interstitial Cells of Cajal in the Colon of Rats with Diabetes Mellitus Type 2" Medicina 59, no. 2: 308. https://doi.org/10.3390/medicina59020308