

Osteogenic Potential of Monosodium Urate Crystals in Synovial Mesenchymal Stem Cells
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation, Culture, and Immunophenotyping
2.2. Immunomagnetic Separation and Immunophenotyping Assay
2.3. Characterization of CD90+ Cells and Colony-Forming Unit (CFU) Assay
2.4. Cell Stimulation
2.5. Analysis of Cell Viability
2.6. Determination of Calcium Nodules
2.7. Analysis of Proinflammatory Cytokines
2.8. Extraction and Quantification of Total Protein
2.9. Analysis of Runx2 Protein Expression by Western Blot
2.10. Statistical Analysis
3. Results
3.1. Immunophenotypic Characterization of Cells Isolated from SM
3.2. Immunomagnetic Separation of CD90+ Cells
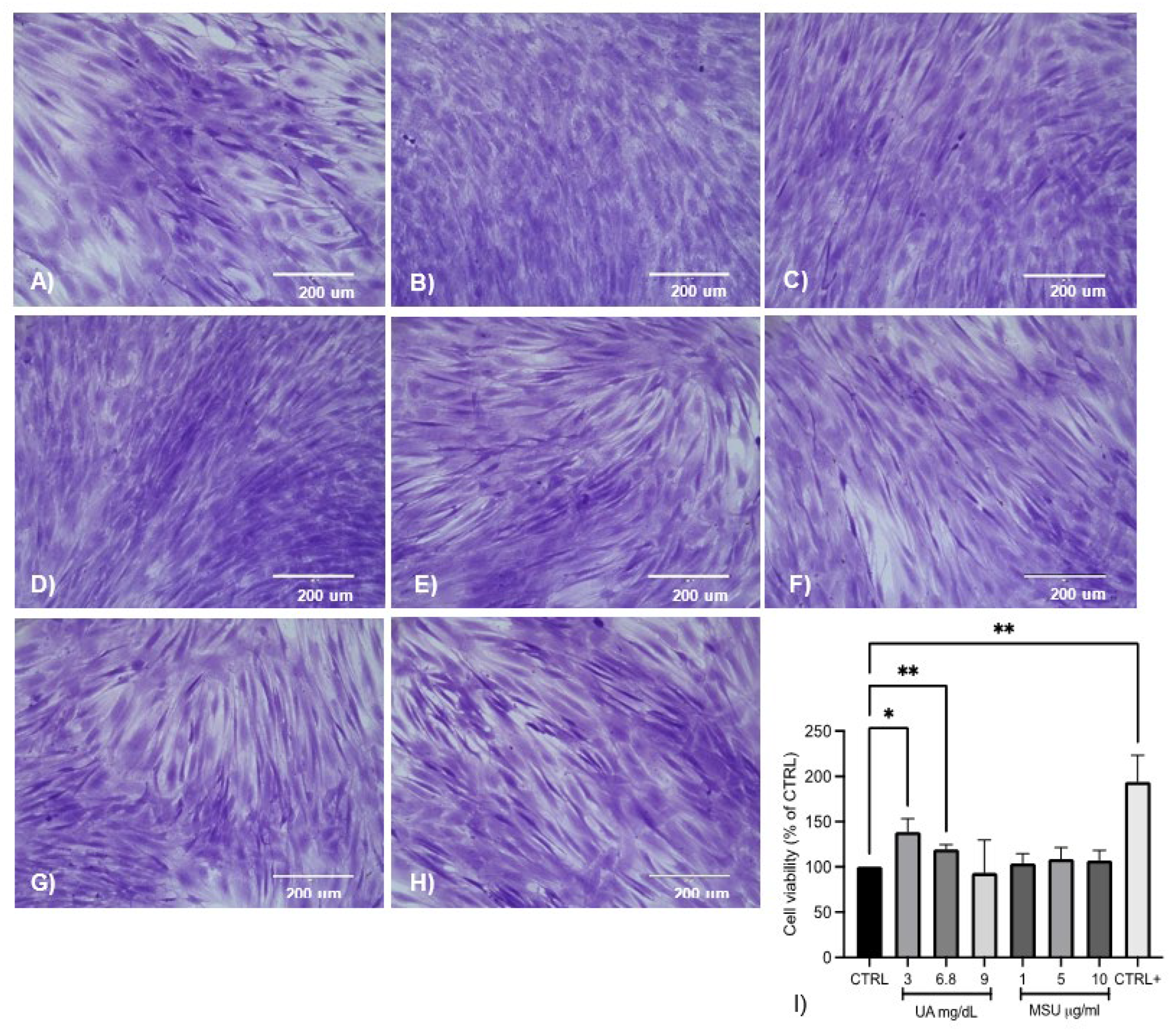
3.3. Effect of UA and MSU Crystals on the Viability of SM-MSCs
3.4. Effect of UA and MSU Crystals on the Formation of Calcium Nodules
3.5. Effect of UA and MSU Crystals on Inflammation in SM-MSCs
3.6. Analysis of Runx2 Protein Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Anghelescu, A. Multiarticular deforming and erosive tophaceous gout with severe comorbidities. J. Clin. Rheumatol. 2019, 26, e269–e271. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.J.; Moon, K.W. Presence of tophi is associated with a rapid decline in the renal function in patients with gout. Sci. Rep. 2021, 11, 5684. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Chen, J.Y.; Wu, H.X.; Zhou, Q.J.; Chen, H.Y.; Lu, Y.F.; Yu, R.S. Relationship between urate within tophus and bone erosion according to the anatomic location of urate deposition in gout: A quantitative analysis using dual-energy CT volume measurements. Medicine 2019, 98, e18431. [Google Scholar] [CrossRef] [PubMed]
- McQueen, F.M.; Doyle, A.; Reeves, Q.; Gao, A.; Tsai, A.; Gamble, G.D.; Curteis, B.; Williams, M.; Dalbeth, N. Bone erosions in patients with chronic gouty arthropathy are associated with tophi but not bone oedema or synovitis: New insights from a 3 T MRI study. Rheumatol 2014, 53, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalbeth, N.; Smith, T.; Nicolson, B.; Clark, B.; Callon, K.; Naot, D.; Haskard, D.O.; McQueen, F.M.; Reid, I.R.; Cornish, J. Enhanced osteoclastogenesis in patients with tophaceous gout: Urate crystals promote osteoclast development through interactions with stromal cells. Arthritis. Rheum. 2008, 58, 1854–1865. [Google Scholar] [CrossRef]
- Chhana, A.; Callon, K.E.; Pool, B.; Naot, D.; Watson, M.; Gamble, G.D.; McQueen, F.M.; Cornish, J.; Dalbeth, N. Monosodium urate monohydrate crystals inhibit osteoblast viability and function: Implications for development of bone erosion in gout. Ann. Rheum. Dis. 2011, 70, 1684–1691. [Google Scholar] [CrossRef]
- Chhana, A.; Pool, B.; Callon, K.E.; Tay, M.L.; Musson, D.; Naot, D.; McCarthy, G.; McGlashan, S.; Cornish, J.; Dalbeth, N. Monosodium urate crystals reduce osteocyte viability and indirectly promote a shift in osteocyte function towards a proinflammatory and proresorptive state. Arthritis Res. Ther. 2018, 20, 208. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Xu, L.; Lin, P.; Cui, J. Uric acid promotes neuronal differentiation of human placenta-derived mesenchymal stem cells in a time- and concentration-dependent manner. Neural. Regen. Res. 2012, 7, 756–760. [Google Scholar] [CrossRef]
- Xu, L.; Han, Y.; Li, P.; Ma, L.; Xin, Y.; Hao, X.X.; Huang, H.; Liu, B.; Yang, N. The Effects of Uric Acid on Bone Mesenchymal Stem Cells Osteogenic Differentiation. J. Appl. Sci. Eng. Innov. 2017, 4, 39–45. Available online: http://www.jasei.org/PDF/4-2/4-39-45.pdf (accessed on 22 August 2022).
- Li, H.Z.; Chen, Z.; Hou, C.L.; Tang, Y.X.; Wang, F.; Fu, Q.G. Uric Acid Promotes Osteogenic Differentiation and Inhibits Adipogenic Differentiation of Human Bone Mesenchymal Stem Cells. J. Biochem. Mol. Toxicol. 2015, 29, 382–387. [Google Scholar] [CrossRef]
- Von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gotzsche, P.; Vandenbroucke, J.P. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. J. Clin. Epidemiol. 2008, 61, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Makhmalbaf, H.; Moradi, A.; Ganji, S.; Omidi-Kashani, F. Accuracy of lachman and anterior drawer tests for anterior cruciate ligament injuries. Arch. Bone Jt. Surg. 2013, 1, 94–97. [Google Scholar] [PubMed]
- Stemcell Technologies. Technical Manual: Human Colony-Forming Unit (CFU) Assays Using MethoCultTM. 2019. (Document #28404/Version 4.6.0). Available online: https://cdn.stemcell.com/media/files/manual/MA28404-Human_Colony_Forming_Unit_Assays_Using_MethoCult.pdf (accessed on 22 August 2022).
- Flick, D.A.; Gifford, G.E. Comparison of in vitro cell cytotoxic assays for tumor necrosis factor. J. Immunol. Methods. 1984, 68, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Lozada, L.G.; Lanaspa, M.A.; Cristóbal-García, M.; García-Arroyo, F.; Soto, V.; Cruz-Robles, D.; Nakagawa, T.; Yu, M.A.; Kang, D.H.; Johnson, R.J. Uric acid-induced endothelial dysfunction is associated with mitochondrial alterations and decreased intracellular ATP concentrations. Nephron Exp. Nephrol. 2013, 121, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamudio-Cuevas, Y.E.; Martínez-Flores, K.; Fernández-Torres, J.; Loissell-Baltazar, Y.A.; Medina-Luna, D.; López Macay, A.; Camacho-Galindo, J.; Hernández-Díaz, C.; Santamaría-Olmedo, M.G.; López-Villegas, E.O.; et al. Monosodium urate crystals induce oxidative stress in human synoviocytes. Arthritis Res. Ther. 2016, 18, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebs, A.; Nyffeler, J.; Rahnenführer, J.; Leist, M. Corrigendum to Normalization of data for viability and relative cell function curves. ALTEX Altern to Anim Exp. 2019, 36, 505. [Google Scholar] [CrossRef]
- Arufe, M.C.; De La Fuente, A.; Fuentes, I.; de Toro, F.J.; Blanco, F.J. Chondrogenic potential of subpopulations of cells expressing mesenchymal stem cell markers derived from human synovial membranes. J. Cell Biochem. 2010, 111, 834–845. [Google Scholar] [CrossRef] [Green Version]
- Harvanová, D.; Tóthová, T.; Šarišský, M.; Amrichová, J.; Rosocha, J. Isolation and characterization of synovial mesenchymal stem cells. Folia Biol. 2011, 57, 119–124. [Google Scholar]
- Zamudio-Cuevas, Y.; Plata-Rodríguez, R.; Fernández-Torres, J.; Martínez-Flores, K.; Cárdenas-Soria, V.H.; Olivos-Meza, A.; Hernández-Rangel, A.; Landa-Solís, C. Synovial membrane mesenchymal stem cells for cartilaginous tissues repair. Mol. Biol. Rep. 2022, 49, 2503–2517. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Xie, H.Q.; Silini, A.; Parolini, O.; Zhang, Y.; Deng, L.; Huang, Y.C. Mesenchymal Stem/Progenitor Cells Derived from Articular Cartilage, Synovial Membrane and Synovial Fluid for Cartilage Regeneration: Current Status and Future Perspectives. Stem. Cell Rev. Rep. 2017, 13, 575–586. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Muneta, T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis Rheum. 2005, 52, 2521–2529. [Google Scholar] [CrossRef]
- Segawa, Y.; Muneta, T.; Makino, H.; Nimura, A.; Mochizuki, T.; Ju, Y.J.; Ezura, Y.; Umezawa, A.; Sekiya, I. Mesenchymal stem cells derived from synovium, meniscus, anterior cruciate ligament, and articular chondrocytes share similar gene expression profiles. J. Orthop. Res. 2009, 27, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Prado, A.A.F.; Favaron, P.O.; da Silva, L.C.L.C.; Baccarin, R.Y.A.; Miglino, M.A.; Maria, D.A. Characterization of mesenchymal stem cells derived from the equine synovial fluid and membrane. BMC Vet. Res. 2015, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, A.; Uchida, S.; Utsunomiya, H.; Tsukamoto, M.; Nakashima, H.; Nakamura, E.; Pascual-Garrido, C.; Sekiya, I.; Sakai, A. Isolation and Characterization of Synovial Mesenchymal Stem Cell Derived from Hip Joints: A Comparative Analysis with a Matched Control Knee Group. Stem. Cells Int. 2017, 2017, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Liang, Y.; Xu, X.; Li, X.; Liu, Q.; Ou, Y.; Duan, L.; Zhu, W.; Lu, W.; Xiong, J.; et al. Isolation and characterization of human mesenchymal stem cells derived from synovial fluid by magnetic-activated cell sorting (MACS). Cell Biol. Int. 2018, 42, 262–271. [Google Scholar] [CrossRef]
- Zamudio-Cuevas, Y.; Fernández-Torres, J.; Martínez-Nava, G.A.; Martínez-Flores, K.; Ramírez-Olvera, A.; Medina-Luna, D.; Hernández-Pérez, A.D.; Landa-Solís, C.; López-Reyes, A. Phagocytosis of monosodium urate crystals by human synoviocytes induces inflammation. Exp. Biol. Med. 2019, 244, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.C.; Zhu, X.X.; Xue, Y.; Zhang, L.H.; Zou, H.J.; Qiu, J.H.; Liu, Q. Role of the NLRP3 inflammasome in the transient release of IL-1β induced by monosodium urate crystals in human fibroblast-like synoviocytes. J. Inflamm. 2015, 12, 30. [Google Scholar] [CrossRef] [Green Version]
- Braga, T.T.; Forni, M.F.; Correa-Costa, M.; Ramos, R.N.; Barbuto, J.A.; Branco, P.; Castoldi, A.; Hiyane, M.I.; Davanso, M.; Latz, E.; et al. Soluble Uric Acid Activates the NLRP3 Inflammasome. Sci. Rep. 2017, 7, 39884. [Google Scholar] [CrossRef] [Green Version]
- Dalbeth, N.; Pool, B.; Shaw, O.M.; Harper, J.L.; Tan, P.; Franklin, C.; House, M.E.; Cornish, J.; Naot, D. Role of miR-146a in regulation of the acute inflammatory response to monosodium urate crystals. Ann. Rheum. Dis. 2015, 74, 786–790. [Google Scholar] [CrossRef]
- Zhao, J.; Wei, K.; Jiang, P.; Chang, C.; Xu, L.; Xu, L.; Shi, Y.; Guo, S.; Xue, Y.; He, D. Inflammatory Response to Regulated Cell Death in Gout and Its Functional Implications. Front. Immunol. 2022, 13, 888306. [Google Scholar] [CrossRef]
- Yan, B.; Liu, D.; Zhu, J.; Pang, X. The effects of hyperuricemia on the differentiation and proliferation of osteoblasts and vascular smooth muscle cells are implicated in the elevated risk of osteopenia and vascular calcification in gout: An in vivo and in vitro analysis. J. Cell Biochem. 2019, 120, 660–662. [Google Scholar] [CrossRef] [PubMed]
- Naot, D.; Pool, B.; Chhana, A.; Gao, R.; Munro, J.; Cornish, J.; Dalbeth, N. Factors secreted by monosodium urate crystal-stimulated macrophages promote a proinflammatory state in osteoblasts: A potential indirect mechanism of bone erosion in gout. Arthritis Res. Ther. 2022, 24, 212. [Google Scholar] [CrossRef] [PubMed]
- Jia, E.; Zhu, H.; Geng, H.; Zhong, L.; Qiu, X.; Xie, J.; Xiao, Y.; Jiang, Y.; Xiao, M.; Zhang, Y.; et al. The Inhibition of Osteoblast Viability by Monosodium Urate Crystal-Stimulated Neutrophil-Derived Exosomes. Front. Immunol. 2022, 13, 809586. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Flores, K.; Plata-Rodríguez, R.; Olivos-Meza, A.; López-Macay, A.; Fernández-Torres, J.; Landa-Solís, C.; Zamudio-Cuevas, Y. Osteogenic Potential of Monosodium Urate Crystals in Synovial Mesenchymal Stem Cells. Medicina 2022, 58, 1724. https://doi.org/10.3390/medicina58121724
Martínez-Flores K, Plata-Rodríguez R, Olivos-Meza A, López-Macay A, Fernández-Torres J, Landa-Solís C, Zamudio-Cuevas Y. Osteogenic Potential of Monosodium Urate Crystals in Synovial Mesenchymal Stem Cells. Medicina. 2022; 58(12):1724. https://doi.org/10.3390/medicina58121724
Chicago/Turabian StyleMartínez-Flores, Karina, Ricardo Plata-Rodríguez, Anell Olivos-Meza, Ambar López-Macay, Javier Fernández-Torres, Carlos Landa-Solís, and Yessica Zamudio-Cuevas. 2022. "Osteogenic Potential of Monosodium Urate Crystals in Synovial Mesenchymal Stem Cells" Medicina 58, no. 12: 1724. https://doi.org/10.3390/medicina58121724