
Bioinformatic Analysis of the BCL-xL/BCL2L1 Interactome in Patients with Pancreatic Cancer
 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nucleotide Sequential Analysis (NSA) of BCL-xL and Construction of Its Gene Network
2.2. In Silico Assessment of the Expression Levels of the BCL-xL Gene
2.3. Validation of Discrimination and Calibration Traits of BCL-xL in the Context of PDAC
2.4. Gene Set Enrichment Analysis (GSEA) Regarding the Molecular Functions and Regulating miRNAs of BCL-xL
2.5. Statistical Analysis
3. Results
3.1. NSA of BCL-xL and Building of Its Interactome
3.2. In Silico Assessment of the Expression Levels of the BCL-xL Network
3.3. Validation of the Discrimination and Calibration Traits of BCL-xL in the Context of PDAC
3.4. GSEA of BCL-xL
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Ervik, M.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide; IARC CancerBase No. 11; International Agency for Research on Cancer: Lyon, France, 2013. Available online: http://globocan.iarc.fr (accessed on 5 June 2019).
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.H.; Chu, P. Immunohistochemical features of the gastrointestinal tract tumors. J. Gastrointest. Oncol. 2012, 3, 262–284. [Google Scholar] [PubMed]
- Neoptolemos, J.P.; Urrutia, R.; Abbruzzese, J.; Büchler, M.W. (Eds.) Pancreatic Cancer; Springer: New York, NY, USA, 2010; Volume LVIII, 1390p. [Google Scholar]
- Pancreatic Cancer—Cancer Stat Facts. Available online: https://seer.cancer.gov/statfacts/html/pancreas.html-80K (accessed on 11 March 2022).
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, M.C.; Zong, W.-X.; Cheng, E.H.-Y.; Lindsten, T.; Panoutsakopoulou, V.; Ross, A.J.; Roth, K.A.; MacGregor, G.R.; Thompson, C.B.; Korsmeyer, S.J. Proapoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opferman, J.T.; Kothari, A. Anti-apoptotic BCL-2 family members in development. Cell Death Differ. 2018, 25, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikezawa, K.; Hikita, H.; Shigekawa, M.; Iwahashi, K.; Eguchi, H.; Sakamori, R.; Tatsumi, T.; Takehara, T. Increased Bcl-xL Expression in Pancreatic Neoplasia Promotes Carcinogenesis by Inhibiting Senescence and Apoptosis. Cell Mol. Gastroenterol. Hepatol. 2017, 4, 185–200.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Takai, D.; Jones, P.A. Comprehensive analysis of CpG islands in human chromosomes 21 and 22. Proc. Natl. Acad. Sci. USA 2002, 99, 3740–3745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLong, E.R.; DeLong, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Hosmer, D.W.; Hosmer, T.; Le Cessie, S.; Lemeshow, S. A comparison of goodness-of-fit tests for the logistic regression model. Stat. Med. 1997, 16, 965–980. [Google Scholar] [CrossRef]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, K.F.; Odinokova, I.V.; Mareninova, O.A.; Rakonczay, Z., Jr.; Hegyi, P.; Pandol, S.J.; Gukovsky, I.; Gukovskaya, A.S.; Rakonczay, Z. Prosurvival Bcl-2 proteins stabilize pancreatic mitochondria and protect against necrosis in experimental pancreatitis. Exp. Cell Res. 2009, 315, 1975–1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, S.N.; Chen, L.; Dewson, G.; Wei, A.; Naik, E.; Fletcher, J.I.; Adams, J.M.; Huang, D.C. Proapoptotic Bak is sequestered by Mcl-1 and Bcl-xL, but not Bcl-2, until displaced by BH3-only proteins. Genes Dev. 2005, 19, 1294–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattler, M.; Liang, H.; Nettesheim, D.; Meadows, R.P.; Harlan, J.E.; Eberstadt, M.; Yoon, H.S.; Shuker, S.B.; Chang, B.S.; Minn, A.J.; et al. Structure of Bcl-xL-Bak peptide complex: Recognition between regulators of apoptosis. Science 1997, 275, 983–986. [Google Scholar] [CrossRef] [PubMed]
- Loo, L.S.W.; Soetedjo, A.A.P.; Lau, H.H.; Ng, N.H.J.; Ghosh, S.; Nguyen, L.; Krishnan, V.G.; Choi, H.; Roca, X.; Hoon, S.; et al. BCL-xL/BCL2L1 is a critical anti-apoptotic protein that promotes the survival of differentiating pancreatic cells from human pluripotent stem cells. Cell Death Dis. 2020, 11, 378. [Google Scholar] [CrossRef] [PubMed]
- Carrington, E.M.; McKenzie, M.D.; Jansen, E.; Myers, M.; Fynch, S.; Kos, C.; Strasser, A.; Kay, T.W.; Scott, C.L.; Allison, J. Islet beta-cells deficient in Bcl-xL develop but are abnormally sensitive to apoptotic stimuli. Diabetes 2009, 58, 2316–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, K.; Suda, T. Metabolic requirements for the maintenance of self-renewing stem cells. Nat. Rev. Mol. Cell Biol. 2014, 15, 243–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.B.; Aon, M.A.; Hsu, Y.T.; Soane, L.; Teng, X.; McCaffery, J.M.; Cheng, W.C.; Qi, B.; Li, H.; Alavian, K.N.; et al. Bcl-xL regulates mitochondrial energetics by stabilizing the inner membrane potential. J. Cell Biol. 2011, 195, 263–276. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Description |
|---|---|
| BCL2L1 | BCL2 Like 1 |
| BCL2 | BCL2 Apoptosis Regulator |
| BID | BH3 Interacting Domain Death Agonist |
| BAK1 | BCL2 Antagonist/Killer 1 |
| BAX | BCL2 Associated X, Apoptosis Regulator |
| BAD | BCL2 Associated Agonist Of Cell Death |
| RAF1 | Raf-1 Proto-Oncogene, Serine/Threonine Kinase |
| BCL2L11 | BCL2 Like 11 |
| BIK | BCL2 Interacting Killer |
| CASP8 | Caspase 8 |
| CASP2 | Caspase 2 |
| PAX3 | Paired Box 3 |
| CFLAR | CASP8 And FADD Like Apoptosis Regulator |
| HRK | Harakiri, BCL2 Interacting Protein |
| PMAIP1 | Phorbol-12-Myristate-13-Acetate-Induced Protein 1 |
| TMBIM6 | Transmembrane BAX Inhibitor Motif Containing 6 |
| BBC3 | BCL2 Binding Component 3 |
| VDAC1 | Voltage Dependent Anion Channel 1 |
| CYCS | Cytochrome C, Somatic |
| SIVA1 | SIVA1 Apoptosis Inducing Factor |
| BCL2L14 | BCL2 Like 14 |
| ID | Name | Source | p-Value | FDR B&H | FDR B&Y | Bonferroni | |
| 1 | GO:0051434 | BH3 domain binding | ToppFun | 2.598 × 10−4 | 4.832 × 10−3 | 1.946 × 10−2 | 8.053 × 10−3 |
| 2 | GO:0097371 | MDM2/MDM4 family protein binding | ToppFun | 4.676 × 10−4 | 4.832 × 10−3 | 1.946 × 10−2 | 1.450 × 10−2 |
| 3 | GO:0070513 | death domain binding | ToppFun | 5.195 × 10−4 | 4.832 × 10−3 | 1.946 × 10−2 | 1.611 × 10−2 |
| 4 | GO:0051400 | BH domain binding | ToppFun | 6.234 × 10−4 | 4.832 × 10−3 | 1.946 × 10−2 | 1.933 × 10−2 |
| 5 | GO:0043027 | cysteine-type endopeptidase inhibitor activity involved in apoptotic process | ToppFun | 1.351 × 10−3 | 8.375 × 10−3 | 3.373 × 10−2 | 4.187 × 10−2 |
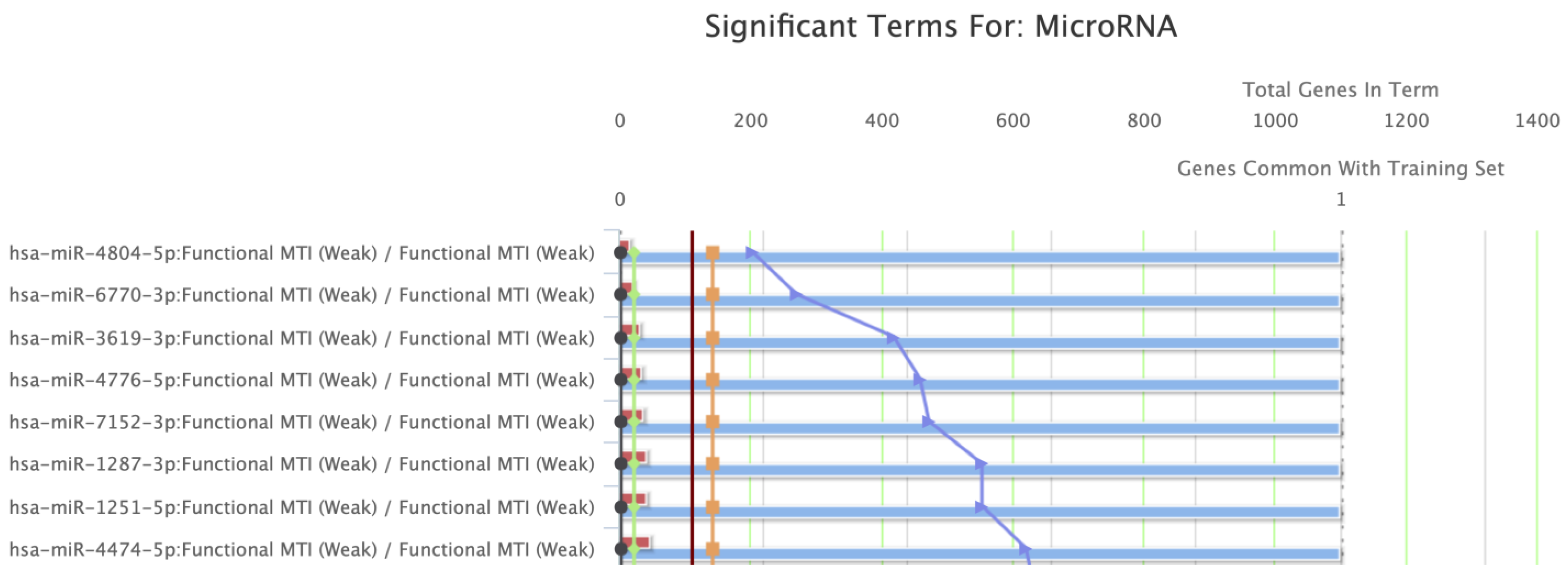
| Regulating miRNA Families | |||||||
| Name | Source | p-Value | FDR B&H | FDR B&Y | Bonferroni | ||
| 1 | hsa-miR-4804-5p | miRTarBase | 2.076 × 10−4 | 9.614 × 10−3 | 6.412 × 10−2 | 9.177 × 10−2 | |
| 2 | hsa-miR-6770-3p | miRTarBase | 2.768 × 10−4 | 9.614 × 10−3 | 6.412 × 10−2 | 1.224 × 10−1 | |
| 3 | hsa-miR-3619-3p | miRTarBase | 4.291 × 10−4 | 9.614 × 10−3 | 6.412 × 10−2 | 1.897 × 10−1 | |
| 4 | hsa-miR-4776-5p | miRTarBase | 4.706 × 10−4 | 9.614 × 10−3 | 6.412 × 10−2 | 2.080 × 10−1 | |
| 5 | hsa-miR-7152-3p | miRTarBase | 4.845 × 10−4 | 9.614 × 10−3 | 6.412 × 10−2 | 2.141 × 10−1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magouliotis, D.E.; Karamolegkou, A.P.; Zotos, P.-A.; Tatsios, E.; Samara, A.A.; Alexopoulou, D.; Koutsougianni, F.; Sakellaridis, N.; Zacharoulis, D.; Dimas, K. Bioinformatic Analysis of the BCL-xL/BCL2L1 Interactome in Patients with Pancreatic Cancer. Medicina 2022, 58, 1663. https://doi.org/10.3390/medicina58111663
Magouliotis DE, Karamolegkou AP, Zotos P-A, Tatsios E, Samara AA, Alexopoulou D, Koutsougianni F, Sakellaridis N, Zacharoulis D, Dimas K. Bioinformatic Analysis of the BCL-xL/BCL2L1 Interactome in Patients with Pancreatic Cancer. Medicina. 2022; 58(11):1663. https://doi.org/10.3390/medicina58111663
Chicago/Turabian StyleMagouliotis, Dimitrios E., Anna P. Karamolegkou, Prokopis-Andreas Zotos, Evangelos Tatsios, Athina A. Samara, Dimitra Alexopoulou, Fani Koutsougianni, Nikos Sakellaridis, Dimitris Zacharoulis, and Konstantinos Dimas. 2022. "Bioinformatic Analysis of the BCL-xL/BCL2L1 Interactome in Patients with Pancreatic Cancer" Medicina 58, no. 11: 1663. https://doi.org/10.3390/medicina58111663