
Topical Application of Galgeunhwanggeumhwangryeon-Tang Recovers Skin-Lipid Barrier and Ameliorates Inflammation via Filaggrin-Thymic Stromal Lymphopoietin-Interleukin 4 Pathway

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of GGRT Extract
2.3. HPLC Analysis
2.4. Bioinformatic Analysis
2.5. Animals
2.6. Induction of Model and Drug Treatment
2.7. Evaluation of Skin Dermatitis Severity
2.8. TEWL and pH Measurement
2.9. Tissue Chemistry
2.10. Immunohistochemistry
2.11. Image Analysis
2.12. Statistical Analysis
3. Results
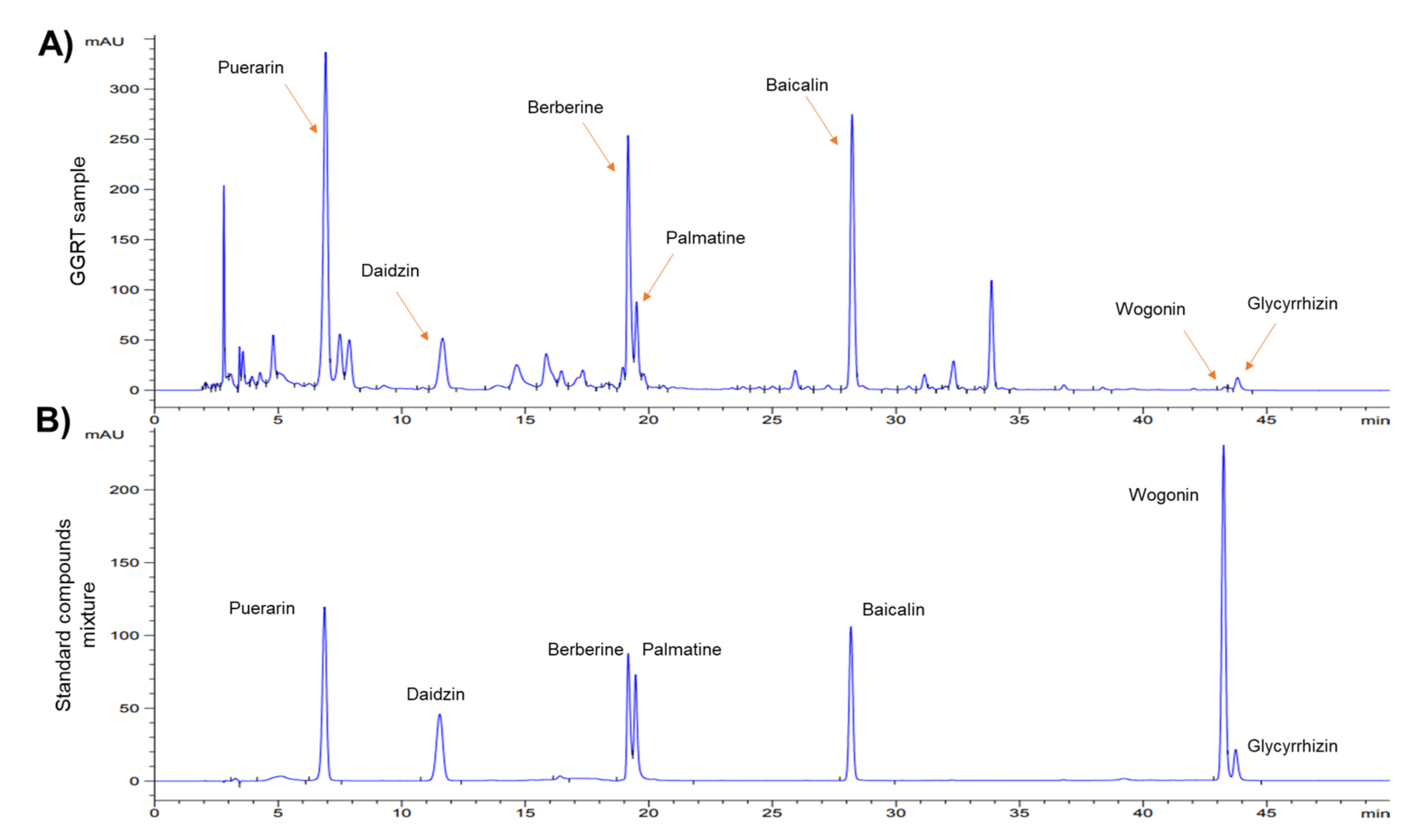
3.1. Identification of GGRT Extract by HPLC Analysis
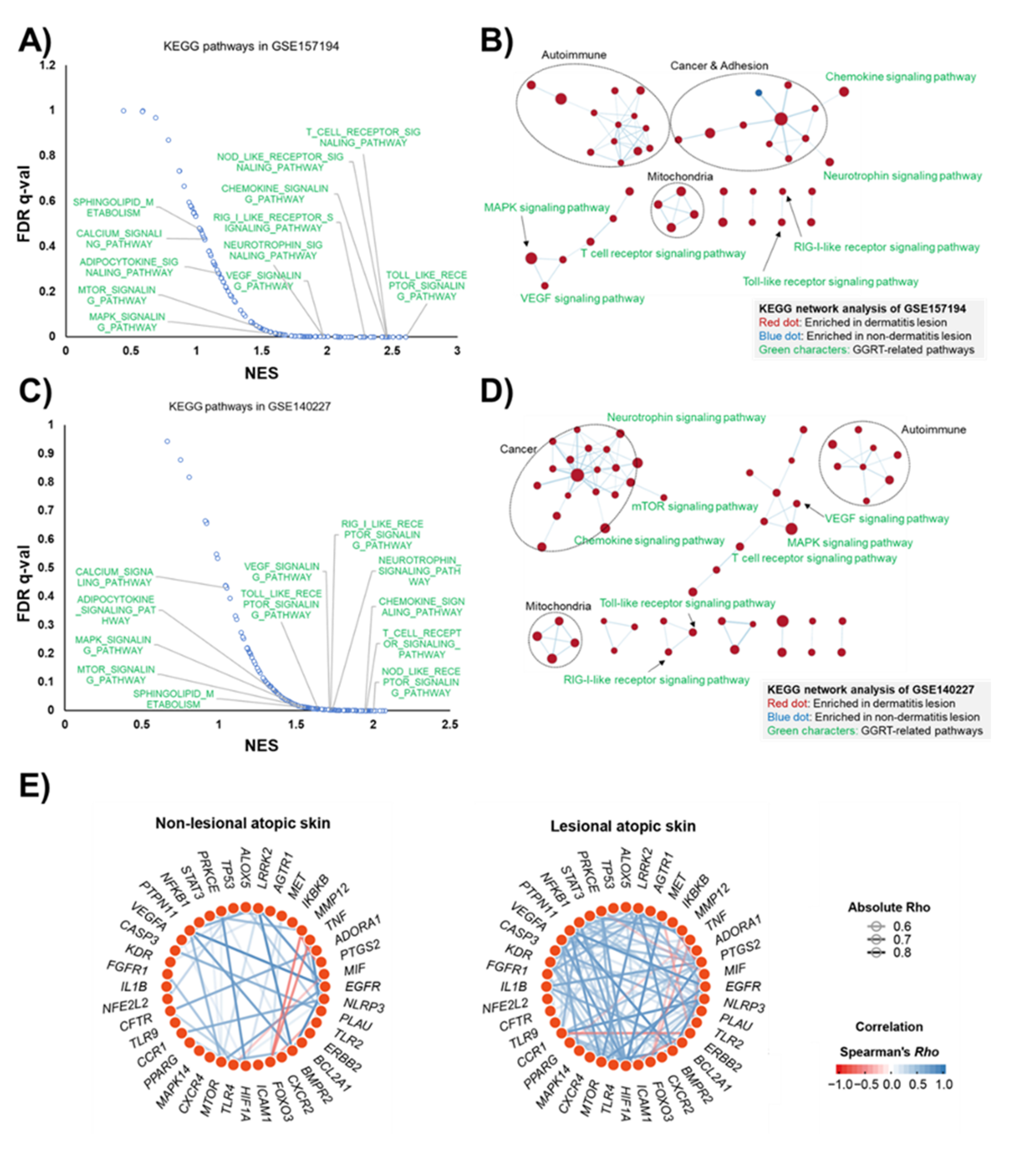
3.2. Human Transcriptomic Analysis Indicates GGRT as a Therapeutic for Atopic Dermatitis
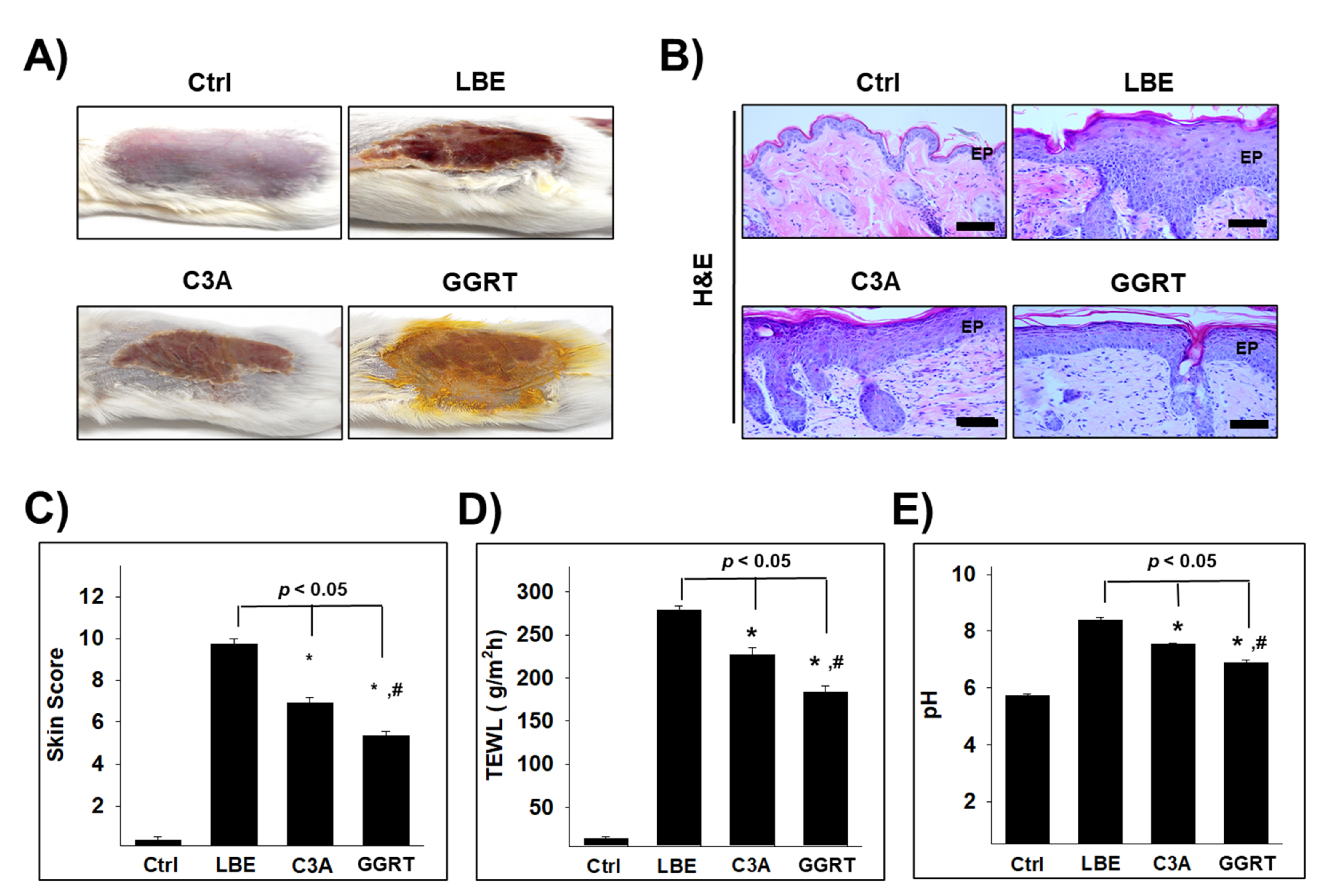
3.3. GGRT Restores the Skin Lipid Barrier
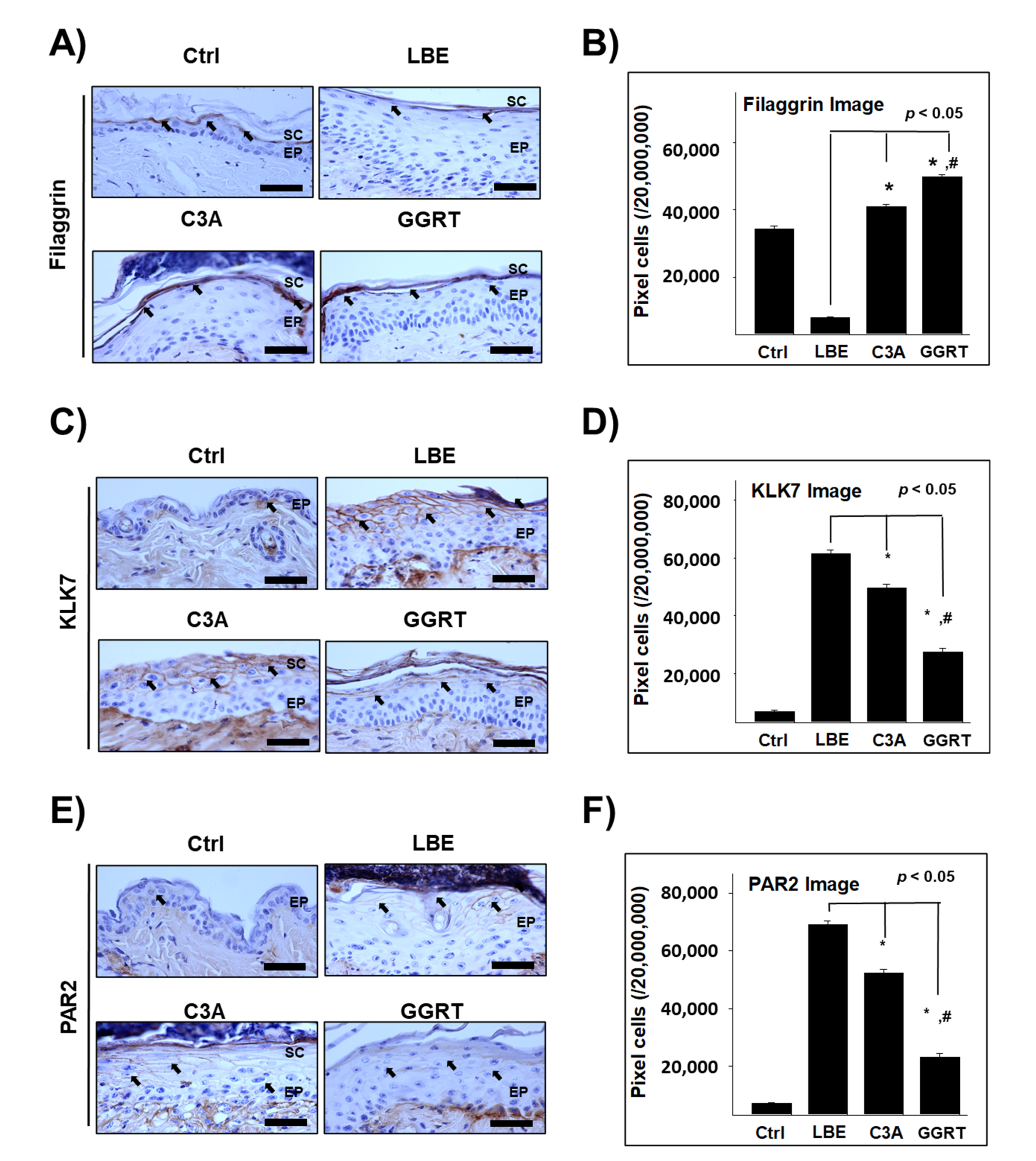
3.4. GGRT Recovers the Skin Barrier-Related Proteins
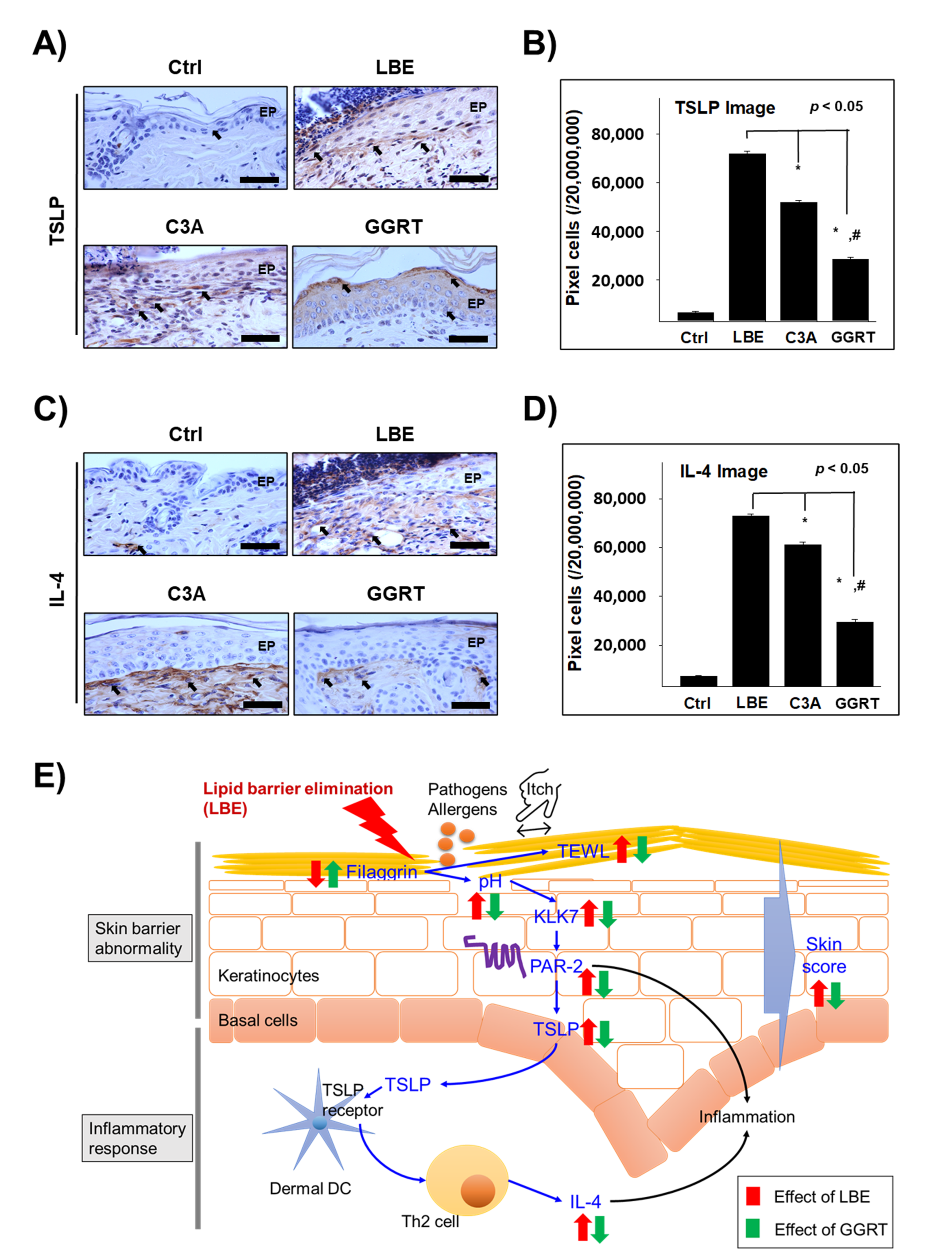
3.5. GGRT Reduces the Th2-Related Inflammatory Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Schmidt, A.D.; De Guzman Strong, C. Current understanding of epigenetics in atopic dermatitis. Exp. Dermatol. 2021, 30, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Baroni, A.; Buommino, E.; De Gregorio, V.; Ruocco, E.; Ruocco, V.; Wolf, R. Structure and function of the epidermis related to barrier properties. Clin. Dermatol. 2012, 30, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Dattola, A.; Bennardo, L.; Silvestri, M.; Nistico, S.P. What’s new in the treatment of atopic dermatitis? Dermatol. Ther. 2019, 32, e12787. [Google Scholar] [CrossRef]
- Panarese, F.; Auriemma, M.; Carbone, A.; Amerio, P. Atopic dermatitis treatment: What’s new on the horizon? G. Ital. Dermatol. Venereol. 2018, 153, 95–101. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y.M. Significance of skin barrier dysfunction in atopic dermatitis. Allergy Asthma Immunol. Res. 2018, 10, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Moreno, A.S.; McPhee, R.; Arruda, L.K.; Howell, M.D. Targeting the T helper 2 inflammatory axis in atopic dermatitis. Int. Arch. Allergy Immunol. 2016, 171, 71–80. [Google Scholar] [CrossRef]
- Tokura, Y.; Phadungsaksawasdi, P.; Ito, T. Atopic dermatitis as Th2 disease revisited. J. Cutan. Immunol. Allergy 2018, 1, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Morelli, P.; Gaspari, M.; Gabriele, C.; Dastoli, S.; Bennardo, L.; Pavel, A.B.; Patruno, C.; Del Duca, E.; Nistico, S.P. Proteomic analysis from skin swabs reveals a new set of proteins identifying skin impairment in atopic dermatitis. Exp. Dermatol. 2021, 30, 811–819. [Google Scholar] [CrossRef]
- Koseki, R.; Morii, W.; Noguchi, E.; Ishikawa, M.; Yang, L.; Yamamoto-Hanada, K.; Narita, M.; Saito, H.; Ohya, Y. Effect of filaggrin loss-of-function mutations on atopic dermatitis in young age: A longitudinal birth cohort study. J. Hum. Genet. 2019, 64, 911–917. [Google Scholar] [CrossRef]
- Yang, G.; Seok, J.K.; Kang, H.C.; Cho, Y.Y.; Lee, H.S.; Lee, J.Y. Skin barrier abnormalities and immune dysfunction in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 2867. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.P.; Chen, F.J.; Jong, M.S.; Tsai, H.L.; Wang, J.R.; Hwang, S.J. Modern use of Chinese herbal formulae from Shang-Han Lun. Chin. Med. J. 2009, 122, 1889–1894. [Google Scholar]
- An, R.; You, L.; Zhang, Y.; Wang, X.; Ma, Y. A rapid UPLC method for simultaneous determination of eleven components in ‘Ge-Gen-Qin-Lian’ decoction. Pharmacogn. Mag. 2014, 10, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Fan, Y.; Du, L.; Mei, Z.; Fu, Y. In silico and in vivo studies on the mechanisms of Chinese medicine formula (gegen qinlian decoction) in the treatment of ulcerative colitis. Front. Pharmacol. 2021, 12, 665102. [Google Scholar] [CrossRef]
- Li, H.; Zhao, L.; Zhang, B.; Jiang, Y.; Wang, X.; Guo, Y.; Liu, H.; Li, S.; Tong, X. A network pharmacology approach to determine active compounds and action mechanisms of ge-gen-qin-lian decoction for treatment of type 2 diabetes. Evid.-Based Complement. Altern. Med. 2014, 2014, 495840. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Zhong, R.; Yang, Y.; Xia, T.; Wang, W.; Wang, Y.; Xing, N.; Luo, Y.; Li, S.; Shang, L.; et al. Systems pharmacology reveals the mechanism of activity of Ge-Gen-Qin-Lian decoction against LPS-induced acute lung injury: A novel strategy for exploring active components and effective mechanism of TCM formulae. Pharmacol. Res. 2020, 156, 104759. [Google Scholar] [CrossRef]
- Xu, J.; Lian, F.; Zhao, L.; Zhao, Y.; Chen, X.; Zhang, X.; Guo, Y.; Zhang, C.; Zhou, Q.; Xue, Z.; et al. Structural modulation of gut microbiota during alleviation of type 2 diabetes with a Chinese herbal formula. ISME J. 2015, 9, 552–562. [Google Scholar] [CrossRef]
- Wang, C.; Xu, N.; Cui, S. Comparative transcriptome analysis of roots, stems, and leaves of Pueraria lobata (Willd.) Ohwi: Identification of genes involved in isoflavonoid biosynthesis. PeerJ 2021, 9, e10885. [Google Scholar] [CrossRef]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharm. 2021, 133, 110917. [Google Scholar] [CrossRef]
- Lv, X.; Li, Y.; Tang, C.; Zhang, Y.; Zhang, J.; Fan, G. Integration of HPLC-based fingerprint and quantitative analyses for differentiating botanical species and geographical growing origins of Rhizoma coptidis. Pharm. Biol. 2016, 54, 3264–3271. [Google Scholar] [CrossRef] [Green Version]
- Ji, S.; Li, Z.; Song, W.; Wang, Y.; Liang, W.; Li, K.; Tang, S.; Wang, Q.; Qiao, X.; Zhou, D.; et al. Bioactive constituents of Glycyrrhiza uralensis (Licorice): Discovery of the effective components of a traditional herbal medicine. J. Nat. Prod. 2016, 79, 281–292. [Google Scholar] [CrossRef]
- Zhang, H.; Ryu, D.; Wu, Y.; Gariani, K.; Wang, X.; Luan, P.; D’Amico, D.; Ropelle, E.R.; Lutolf, M.P.; Aebersold, R.; et al. NAD(+) repletion improves mitochondrial and stem cell function and enhances life span in mice. Science 2016, 352, 1436–1443. [Google Scholar] [CrossRef] [Green Version]
- Mobus, L.; Rodriguez, E.; Harder, I.; Stolzl, D.; Boraczynski, N.; Gerdes, S.; Kleinheinz, A.; Abraham, S.; Heratizadeh, A.; Handrick, C.; et al. Atopic dermatitis displays stable and dynamic skin transcriptome signatures. J. Allergy Clin. Immunol. 2021, 147, 213–223. [Google Scholar] [CrossRef]
- Hwangbo, H.; Lee, H.; Jin, E.J.; Lee, J.; Jo, Y.; Ryu, D.; Kim, G. Bio-printing of aligned GelMa-based cell-laden structure for muscle tissue regeneration. Bioact. Mater. 2022, 8, 57–70. [Google Scholar] [CrossRef]
- Leung, D.Y.; Hirsch, R.L.; Schneider, L.; Moody, C.; Takaoka, R.; Li, S.H.; Meyerson, L.A.; Mariam, S.G.; Goldstein, G.; Hanifin, J.M. Thymopentin therapy reduces the clinical severity of atopic dermatitis. J. Allergy Clin. Immunol. 1990, 85, 927–933. [Google Scholar] [CrossRef]
- Drislane, C.; Irvine, A.D. The role of filaggrin in atopic dermatitis and allergic disease. Ann. Allergy Asthma Immunol. 2020, 124, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.X.J.; Mack, M.R.; Oetjen, L.K.; Trier, A.M.; Council, M.L.; Pavel, A.B.; Guttman-Yassky, E.; Kim, B.S.; Liu, Q. Kallikrein 7 promotes atopic dermatitis-associated itch independently of skin inflammation. J. Investig. Dermatol. 2020, 140, 1244–1252. [Google Scholar] [CrossRef]
- Frateschi, S.; Camerer, E.; Crisante, G.; Rieser, S.; Membrez, M.; Charles, R.P.; Beermann, F.; Stehle, J.C.; Breiden, B.; Sandhoff, K.; et al. PAR2 absence completely rescues inflammation and ichthyosis caused by altered CAP1/Prss8 expression in mouse skin. Nat. Commun. 2011, 2, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, Y.A.; Chung, H.; Yoon, S.; Park, J.I.; Lee, J.E.; Myung, C.H.; Hwang, J.S. Skin barrier recovery by protease-activated receptor-2 antagonist lobaric acid. Biomol. Ther. 2016, 24, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsuno, K.; Fujiyama, T.; Yamaguchi, H.; Waki, M.; Tokura, Y. TSLP directly interacts with skin-homing Th2 cells highly expressing its receptor to enhance IL-4 production in atopic dermatitis. J. Investig. Dermatol. 2015, 135, 3017–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbe-Burger, A.; Egyed, A.; Olt, S.; Klubal, R.; Mann, U.; Rappersberger, K.; Rot, A.; Stingl, G. Overexpression of IL-4 alters the homeostasis in the skin. J. Investig. Dermatol. 2002, 118, 767–778. [Google Scholar] [CrossRef] [Green Version]
- Egawa, G.; Kabashima, K. Barrier dysfunction in the skin allergy. Allergol Int. 2018, 67, 3–11. [Google Scholar] [CrossRef]
- Haftek, M. Epidermal barrier disorders and corneodesmosome defects. Cell Tissue Res. 2015, 360, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Steven, A.C.; Steinert, P.M. Protein composition of cornified cell envelopes of epidermal keratinocytes. J. Cell Sci. 1994, 107 (Pt 2), 693–700. [Google Scholar] [CrossRef]
- Le Lamer, M.; Pellerin, L.; Reynier, M.; Cau, L.; Pendaries, V.; Leprince, C.; Mechin, M.C.; Serre, G.; Paul, C.; Simon, M. Defects of corneocyte structural proteins and epidermal barrier in atopic dermatitis. Biol. Chem. 2015, 396, 1163–1179. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y. Epidermal barrier in atopic dermatitis. Allergy Asthma Immunol. Res. 2012, 4, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Jung, I.K.; Choi, J.; Nam, J.; No, K.T. Modeling lipid layers of atopic skin and observation of changes in lipid layer properties with changes in ceramide content. J. Cosmet. Dermatol. 2021, 20, 2924–2931. [Google Scholar] [CrossRef]
- Van Smeden, J.; Bouwstra, J.A. Stratum Corneum Lipids: Their Role for the Skin Barrier Function in Healthy Subjects and Atopic Dermatitis Patients. Curr. Probl. Dermatol. 2016, 49, 8–26. [Google Scholar] [CrossRef]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- O’Regan, G.M.; Sandilands, A.; McLean, W.H.; Irvine, A.D. Filaggrin in atopic dermatitis. J. Allergy Clin. Immunol. 2009, 124, R2–R6. [Google Scholar] [CrossRef]
- Van den Oord, R.A.; Sheikh, A. Filaggrin gene defects and risk of developing allergic sensitisation and allergic disorders: Systematic review and meta-analysis. BMJ 2009, 339, b2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.J.; Irvine, A.D.; Terron-Kwiatkowski, A.; Sandilands, A.; Campbell, L.E.; Zhao, Y.; Liao, H.; Evans, A.T.; Goudie, D.R.; Lewis-Jones, S.; et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris. Nat. Genet. 2006, 38, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Kezic, S.; O’Regan, G.M.; Yau, N.; Sandilands, A.; Chen, H.; Campbell, L.E.; Kroboth, K.; Watson, R.; Rowland, M.; McLean, W.H.; et al. Levels of filaggrin degradation products are influenced by both filaggrin genotype and atopic dermatitis severity. Allergy 2011, 66, 934–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danby, S.G.; Cork, M.J. pH in Atopic Dermatitis. Curr. Probl. Dermatol. 2018, 54, 95–107. [Google Scholar] [CrossRef]
- Jang, H.; Matsuda, A.; Jung, K.; Karasawa, K.; Matsuda, K.; Oida, K.; Ishizaka, S.; Ahn, G.; Amagai, Y.; Moon, C.; et al. Skin pH is the master switch of kallikrein 5-mediated skin barrier destruction in a murine atopic dermatitis model. J. Investig. Dermatol. 2016, 136, 127–135. [Google Scholar] [CrossRef]
- Yamamoto, M.; Miyai, M.; Matsumoto, Y.; Tsuboi, R.; Hibino, T. Kallikrein-related peptidase-7 regulates caspase-14 maturation during keratinocyte terminal differentiation by generating an intermediate form. J. Biol. Chem. 2012, 287, 32825–32834. [Google Scholar] [CrossRef] [Green Version]
- Henehan, M.; De Benedetto, A. Update on protease-activated receptor 2 in cutaneous barrier, differentiation, tumorigenesis and pigmentation, and its role in related dermatologic diseases. Exp. Dermatol. 2019, 28, 877–885. [Google Scholar] [CrossRef] [Green Version]
- Redhu, D.; Franke, K.; Kumari, V.; Francuzik, W.; Babina, M.; Worm, M. Thymic stromal lymphopoietin production induced by skin irritation results from concomitant activation of protease-activated receptor 2 and interleukin 1 pathways. Br. J. Dermatol. 2020, 182, 119–129. [Google Scholar] [CrossRef]
- Briot, A.; Lacroix, M.; Robin, A.; Steinhoff, M.; Deraison, C.; Hovnanian, A. Par2 inactivation inhibits early production of TSLP, but not cutaneous inflammation, in Netherton syndrome adult mouse model. J. Investig. Dermatol. 2010, 130, 2736–2742. [Google Scholar] [CrossRef] [Green Version]
- Barr, T.P.; Garzia, C.; Guha, S.; Fletcher, E.K.; Nguyen, N.; Wieschhaus, A.J.; Ferrer, L.; Covic, L.; Kuliopulos, A. PAR2 Pepducin-Based Suppression of Inflammation and Itch in Atopic Dermatitis Models. J. Investig. Dermatol. 2019, 139, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J. TSLP in epithelial cell and dendritic cell cross talk. Adv. Immunol. 2009, 101, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Geiger, E.; Magerstaedt, R.; Wessendorf, J.H.; Kraft, S.; Hanau, D.; Bieber, T. IL-4 induces the intracellular expression of the alpha chain of the high-affinity receptor for IgE in in vitro-generated dendritic cells. J. Allergy Clin. Immunol. 2000, 105, 150–156. [Google Scholar] [CrossRef]
- Kumar, S.; Jeong, Y.; Ashraf, M.U.; Bae, Y.S. Dendritic cell-mediated Th2 immunity and immune disorders. Int. J. Mol. Sci. 2019, 20, 2159. [Google Scholar] [CrossRef] [Green Version]
- Cha, H.Y.; Ahn, S.H.; Cheon, J.H.; Park, S.Y.; Kim, K. Hataedock treatment has preventive therapeutic effects for atopic dermatitis through skin barrier protection in Dermatophagoides farinae-induced NC/Nga mice. J. Ethnopharmacol. 2017, 206, 327–336. [Google Scholar] [CrossRef]
- Li, Z.G. Comparative study on WHO Western Pacific Region and World Federation of Chinese Medicine Societies international standard terminologies on traditional medicine: Diseases of Gynecology and Obstetrics (Part 1). J. Integr. Med. 2014, 12, 127–130. [Google Scholar] [CrossRef]
- Muluye, R.A.; Bian, Y.; Alemu, P.N. Anti-inflammatory and antimicrobial effects of heat-clearing chinese herbs: A current review. J. Tradit. Complement. Med. 2014, 4, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Tsuge, A.; Hisaka, S.; Hayashi, H.; Nose, M. Effect of hot water extract of a glycyrrhizin-deficient strain of Glycyrrhiza uralensis on contact hypersensitivity in mice. J. Nat. Med. 2020, 74, 415–420. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, J.; Choi, H.G.; Kim, E.K.; Jun, C.D. Licoricidin abrogates T-cell activation by modulating PTPN1 activity and attenuates atopic dermatitis in vivo. J. Investig. Dermatol. 2021, 141, 2490–2498. [Google Scholar] [CrossRef]
- Lee, J.H.; Jeon, Y.D.; Lee, Y.M.; Kim, D.K. The suppressive effect of puerarin on atopic dermatitis-like skin lesions through regulation of inflammatory mediators in vitro and in vivo. Biochem. Biophys. Res. Commun. 2018, 498, 707–714. [Google Scholar] [CrossRef]
- Jung, H.S.; Kim, M.H.; Gwak, N.G.; Im, Y.S.; Lee, K.Y.; Sohn, Y.; Choi, H.; Yang, W.M. Antiallergic effects of Scutellaria baicalensis on inflammation in vivo and in vitro. J. Ethnopharmacol. 2012, 141, 345–349. [Google Scholar] [CrossRef]
- Akiyama, H.; Nose, M.; Takiguchi, H.; Sugiyama, K.; Tsutsui, R.; Hisaka, S.; Fuchino, H.; Inui, T.; Kawano, N.; Taguchi, T.; et al. Mutagenetic and anti-allergic studies for evaluation of extracts of Coptis Rhizome produced by an artificial hydroponic system. J. Nat. Med. 2019, 73, 608–613. [Google Scholar] [CrossRef]
- Wu, S.; Yu, D.; Liu, W.; Zhang, J.; Liu, X.; Wang, J.; Yu, M.; Li, Z.; Chen, Q.; Li, X.; et al. Magnoflorine from Coptis chinese has the potential to treat DNCB-induced Atopic dermatits by inhibiting apoptosis of keratinocyte. Bioorg. Med. Chem. 2020, 28, 115093. [Google Scholar] [CrossRef]
- Tong, X.L.; Zhao, L.H.; Lian, F.M.; Zhou, Q.; Xia, L.; Zhang, J.C.; Chen, X.Y.; Ji, H.Y. Clinical observations on the dose-effect relationship of gegen qin lian decoction on 54 out-patients with type 2 diabetes. J. Tradit. Chin. Med. 2011, 31, 56–59. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Sheng, Y.; Zeng, G.; Zeng, Z.; Li, B.; Jiang, L.; Xu, G.; Zhang, Q. Metabonomic study on the plasma of high-fat diet-induced dyslipidemia rats treated with Ge Gen Qin lian decoction by ultrahigh-performance liquid chromatography-mass spectrometry. Evid.-Based Complement. Altern. Med. 2021, 2021, 6692456. [Google Scholar] [CrossRef]
- Ren, L.; Cheng, Y.; Qin, F. Herbal formula Gegen-Qinlian decoction for type 2 Diabetes mellitus: A meta-analysis of randomized controlled Trials. Evid.-Based Complement. Altern. Med. 2020, 2020, 3907920. [Google Scholar] [CrossRef]
- Yan, Y.; Du, C.; Li, Z.; Zhang, M.; Li, J.; Jia, J.; Li, A.; Qin, X.; Song, Q. Comparing the antidiabetic effects and chemical profiles of raw and fermented Chinese Ge-Gen-Qin-Lian decoction by integrating untargeted metabolomics and targeted analysis. Chin. Med. 2018, 13, 54. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Zhang, J.; Wang, Y.; Zhang, Z.; Wang, F.; Tang, X. Uncovering the mechanism of Ge-Gen-Qin-Lian decoction for treating ulcerative colitis based on network pharmacology and molecular docking verification. Biosci. Rep. 2021, 41, BSR20203565. [Google Scholar] [CrossRef]
- Lee, B.S.; Shim, S.M.; Heo, J.; Pae, H.O.; Seo, B.Y.; Han, S.Y.; Sohn, D.H.; Jang, S.I.; Chung, H.T. Wogonin suppresses TARC expression induced by mite antigen via heme oxygenase 1 in human keratinocytes. Suppressive effect of wogonin on mite antigen-induced TARC expression. J. Dermatol. Sci. 2007, 46, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Andoh, T.; Yoshihisa, Y.; Rehman, M.U.; Tabuchi, Y.; Shimizu, T. Berberine induces anti-atopic dermatitis effects through the downregulation of cutaneous EIF3F and MALT1 in NC/Nga mice with atopy-like dermatitis. Biochem. Pharmacol. 2021, 185, 114439. [Google Scholar] [CrossRef]
- Zhou, B.G.; Zhao, H.M.; Lu, X.Y.; Zhou, W.; Liu, F.C.; Liu, X.K.; Liu, D.Y. Effect of puerarin regulated mTOR signaling pathway in experimental liver injury. Front. Pharmacol. 2018, 9, 1165. [Google Scholar] [CrossRef] [Green Version]
- Li, S.J.; Sun, S.J.; Gao, J.; Sun, F.B. Wogonin induces Beclin-1/PI3K and reactive oxygen species-mediated autophagy in human pancreatic cancer cells. Oncol. Lett. 2016, 12, 5059–5067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, L.; Chen, Q.; Gong, K.; Xu, X.; Xie, Y.; Zhang, W.; Cao, H.; Hu, T.; Hong, X.; Zhan, Y.Y. Berberine decelerates glucose metabolism via suppression of mTORdependent HIF1alpha protein synthesis in colon cancer cells. Oncol. Rep. 2018, 39, 2436–2442. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Qiu, D.; Huang, G.; Chen, J.; Wu, Q.; Xiong, S.; Wu, C.; Peng, Y.; Zhang, Q. Wogonin suppresses IL-10 production in B cells via STAT3 and ERK signaling pathway. J. Immunol. Res. 2020, 2020, 3032425. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Wan, H.; Yi, L.; Chen, W.; Luo, Y.; Huang, Y.; Liu, X. Berberine administrated with different routes attenuates inhaled LPS-induced acute respiratory distress syndrome through TLR4/NF-kappaB and JAK2/STAT3 inhibition. Eur. J. Pharmacol. 2021, 908, 174349. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ge, S.; Wang, Y.; Liu, Y.; Qiu, L.; Li, J.; Huang, X.; Sun, L. Puerarin alleviates UUO-Induced inflammation and fibrosis by regulating the NF-kappaB P65/STAT3 and TGFbeta1/Smads signaling pathways. Drug Des. Devel. Ther. 2021, 15, 3697–3708. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Willenborg, S.; Bloch, W.; Wickstrom, S.A.; Wagle, P.; Brodesser, S.; Roers, A.; Jais, A.; Bruning, J.C.; Hall, M.N.; et al. Epidermal mammalian target of rapamycin complex 2 controls lipid synthesis and filaggrin processing in epidermal barrier formation. J. Allergy Clin. Immunol. 2020, 145, 283–300.e288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, W.I.; Lee, H.; Bae, H.C.; Ryu, H.J.; Son, S.W. IL-33 down-regulates filaggrin expression by inducing STAT3 and ERK phosphorylation in human keratinocytes. J. Dermatol. Sci. 2016, 82, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Ryuk, J.A.; Lixia, M.; Cao, S.; Ko, B.S.; Park, S. Efficacy and safety of Gegen Qinlian decoction for normalizing hyperglycemia in diabetic patients: A systematic review and meta-analysis of randomized clinical trials. Complement. Ther. Med. 2017, 33, 6–13. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Buist, H.; Craig, P.; Dewhurst, I.; Hougaard Bennekou, S.; Kneuer, C.; Machera, K.; Pieper, C.; Court Marques, D.; Guillot, G.; et al. Guidance on dermal absorption. EFSA J. 2017, 15, e04873. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Botanical Name | Marker Compounds | Amount (g) |
|---|---|---|---|
| Pueraria lobata (Willd.) Ohwi | Puerariae Radix | Puerarin, Daidzin | 80 |
| Scutellaria baicalensis George | Scutellaria Radix | Baicalin, Wogonin | 20 |
| Coptis japonica Makino | Coptidis Rhizoma | Berberine, Palmatine | 30 |
| Glycyrrhiza uralensis Fischer | Glycyrrhiza Radix | Glycyrrhizin | 20 |
| Total | 150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, S.-H.; Shin, S.; Do, Y.; Jo, Y.; Ryu, D.; Ha, K.-T.; Kim, K. Topical Application of Galgeunhwanggeumhwangryeon-Tang Recovers Skin-Lipid Barrier and Ameliorates Inflammation via Filaggrin-Thymic Stromal Lymphopoietin-Interleukin 4 Pathway. Medicina 2021, 57, 1387. https://doi.org/10.3390/medicina57121387
Ahn S-H, Shin S, Do Y, Jo Y, Ryu D, Ha K-T, Kim K. Topical Application of Galgeunhwanggeumhwangryeon-Tang Recovers Skin-Lipid Barrier and Ameliorates Inflammation via Filaggrin-Thymic Stromal Lymphopoietin-Interleukin 4 Pathway. Medicina. 2021; 57(12):1387. https://doi.org/10.3390/medicina57121387
Chicago/Turabian StyleAhn, Sang-Hyun, Su Shin, Yoonju Do, Yunju Jo, Dongryeol Ryu, Ki-Tae Ha, and Kibong Kim. 2021. "Topical Application of Galgeunhwanggeumhwangryeon-Tang Recovers Skin-Lipid Barrier and Ameliorates Inflammation via Filaggrin-Thymic Stromal Lymphopoietin-Interleukin 4 Pathway" Medicina 57, no. 12: 1387. https://doi.org/10.3390/medicina57121387