Similar Allergenicity to Different Artemisia Species Is a Consequence of Highly Cross-Reactive Art v 1-Like Molecules


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients’ Sera
2.2. Preparation of Artemisia spp. Pollen Extracts
2.3. Protein Gel Electrophoresis and Immunoblotting
2.4. Protein Purification
2.5. Mass Spectrometry and N-Terminal Sequencing
2.6. cDNA Cloning
2.7. ELISA and Cross-Inhibition Studies
2.8. Mediator Release Assay
2.9. T Cell Reactivity
2.10. ISAC Inhibition Studies
2.11. Statistics
3. Results
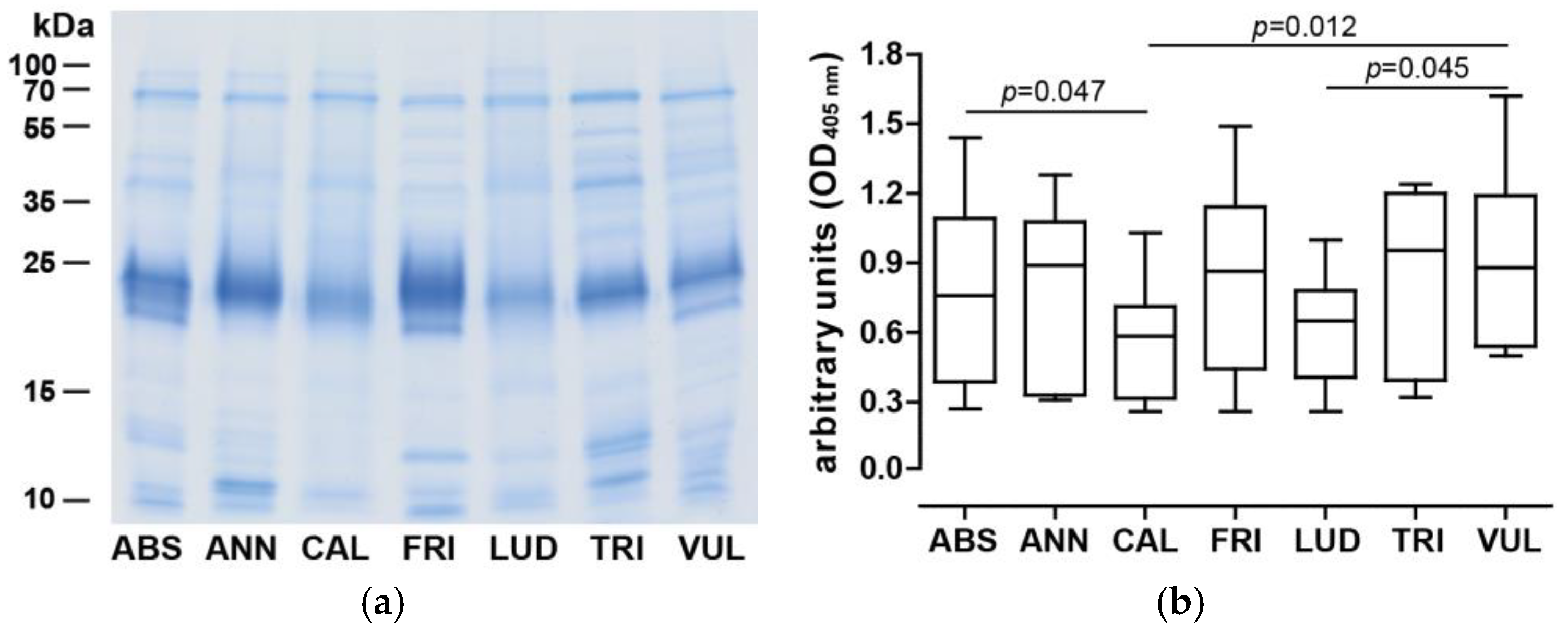
3.1. Geographic Distribution of Artemisia spp. and Analysis of Pollen Proteins
3.2. IgE Binding Capacity and Art v 1 Cross-Reactivity Pattern of Artemisia Extracts
3.3. Purification of DPLPs
3.4. cDNA Cloning and Amino Acid Sequence Identification
3.5. IgE Reactivity to Purified DPLPs
3.6. Mediator Release Assay
3.7. Art v 1 and DPLPs Presented T cell Cross-Reactivity
3.8. IgE Cross-Reactivity within Other Allergenic Protein Families
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pablos, I.; Wildner, S.; Asam, C.; Wallner, M.; Gadermaier, G. Pollen Allergens for Molecular Diagnosis. Curr. Allergy Asthma Rep. 2016, 16, 31. [Google Scholar] [CrossRef] [PubMed]
- Cecchi, L.; D’Amato, G.; Ayres, J.G.; Galan, C.; Forastiere, F.; Forsberg, B.; Gerritsen, J.; Nunes, C.; Behrendt, H.; Akdis, C.; et al. Projections of the effects of climate change on allergic asthma: The contribution of aerobiology. Allergy 2010, 65, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Storkey, J.; Stratonovitch, P.; Chapman, D.S.; Vidotto, F.; Semenov, M.A. A process-based approach to predicting the effect of climate change on the distribution of an invasive allergenic plant in europe. PLoS ONE 2014, 9, e88156. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Chauhan, B.S.; Farooq, M.; Shabbir, A.; Adkins, S.W. What do we really know about alien plant invasion? A review of the invasion mechanism of one of the world’s worst weeds. Planta 2016, 244, 39–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bielory, L.; Mi, Z.; Cai, T.; Robock, A.; Georgopoulos, P. Allergenic pollen season variations in the past two decades under changing climate in the United States. Glob. Chang. Biol. 2015, 21, 1581–1589. [Google Scholar] [CrossRef] [PubMed]
- Lake, I.R.; Jones, N.R.; Agnew, M.; Goodess, C.M.; Giorgi, F.; Hamaoui-Laguel, L.; Semenov, M.A.; Solomon, F.; Storkey, J.; Vautard, R.; et al. Climate Change and Future Pollen Allergy in Europe. Environ. Health Perspect. 2016, 125, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Jager, S.; Berger, U.; Sikoparija, B.; Hallsdottir, M.; Sauliene, I.; Bergmann, K.C.; Pashley, C.H.; de Weger, L.; Majkowska-Wojciechowska, B.; et al. Geographic and temporal variations in pollen exposure across Europe. Allergy 2014, 69, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.W.; Lee, H.B.; Kang, I.J.; Kim, S.W.; Park, K.S.; Kook, M.H.; Kim, B.S.; Baek, H.S.; Kim, J.H.; Kim, J.K.; et al. The revised edition of korean calendar for allergenic pollens. Allergy Asthma Immunol. Res. 2012, 4, 5–11. [Google Scholar] [CrossRef]
- Kindermans, J.M.; Pilloy, J.; Olliaro, P.; Gomes, M. Ensuring sustained ACT production and reliable artemisinin supply. Malar. J. 2007, 6, 125. [Google Scholar] [CrossRef]
- White, N.J. Qinghaosu (artemisinin): The price of success. Science 2008, 320, 330–334. [Google Scholar] [CrossRef]
- Ho, W.E.; Peh, H.Y.; Chan, T.K.; Wong, W.S. Artemisinins: Pharmacological actions beyond anti-malarial. Pharmacol. Ther. 2014, 142, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Wopfner, N.; Gadermaier, G.; Egger, M.; Asero, R.; Ebner, C.; Jahn-Schmid, B.; Ferreira, F. The spectrum of allergens in ragweed and mugwort pollen. Int. Arch. Allergy Immunol. 2005, 138, 337–346. [Google Scholar] [CrossRef]
- Qin, P.; Waltoft, B.L.; Mortensen, P.B.; Postolache, T.T. Suicide risk in relation to air pollen counts: A study based on data from Danish registers. BMJ Open 2013, 3. [Google Scholar] [CrossRef]
- Annesi-Maesano, I.; Rouve, S.; Desqueyroux, H.; Jankovski, R.; Klossek, J.M.; Thibaudon, M.; Demoly, P.; Didier, A. Grass pollen counts, air pollution levels and allergic rhinitis severity. Int. Arch. Allergy Immunol. 2012, 158, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.D.; Zheng, Y.W.; Gjesing, B.; Kong, X.A.; Wang, J.Y.; Song, Z.J.; Lai, X.X.; Zhong, N.S.; Spangfort, M.D. Prevalence of sensitization to weed pollens of Humulus scandens, Artemisia vulgaris, and Ambrosia artemisiifolia in northern China. J. Zhejiang Univ. Sci. B 2013, 14, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.L.; Lee, S.K.; Oh, S.H.; Moon, B.S.; Park, H.S.; Hong, C.S. A study of allergy skin tests with Korean pollen extracts. Yonsei Med J. 1987, 28, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Himly, M.; Jahn-Schmid, B.; Dedic, A.; Kelemen, P.; Wopfner, N.; Altmann, F.; van Ree, R.; Briza, P.; Richter, K.; Ebner, C.; et al. Art v 1, the major allergen of mugwort pollen, is a modular glycoprotein with a defensin-like and a hydroxyproline-rich domain. Faseb J. 2003, 17, 106–108. [Google Scholar] [CrossRef]
- Gao, Z.S.; Fu, W.Y.; Sun, Y.M.; Gao, B.Y.; Wang, H.Y.; Liu, M.L.; Luo, F.M.; Zhou, X.; Jin, J.; Zhao, L.; et al. Artemisia pollen allergy in China: Component-resolved diagnosis reveals allergic asthma patients have significant multiple allergen sensitization. Allergy 2018, 74, 284–293. [Google Scholar] [CrossRef]
- Lombardero, M.; Garcia-Selles, F.J.; Polo, F.; Jimeno, L.; Chamorro, M.J.; Garcia-Casado, G.; Sanchez-Monge, R.; Diaz-Perales, A.; Salcedo, G.; Barber, D. Prevalence of sensitization to Artemisia allergens Art v 1, Art v 3 and Art v 60 kDa. Cross-reactivity among Art v 3 and other relevant lipid-transfer protein allergens. Clin. Exp. Allergy 2004, 34, 1415–1421. [Google Scholar] [CrossRef]
- Dedic, A.; Gadermaier, G.; Vogel, L.; Ebner, C.; Vieths, S.; Ferreira, F.; Egger, M. Immune recognition of novel isoforms and domains of the mugwort pollen major allergen Art v 1. Mol. Immunol. 2009, 46, 416–421. [Google Scholar] [CrossRef]
- Gadermaier, G.; Jahn-Schmid, B.; Vogel, L.; Egger, M.; Himly, M.; Briza, P.; Ebner, C.; Vieths, S.; Bohle, B.; Ferreira, F. Targeting the cysteine-stabilized fold of Art v 1 for immunotherapy of Artemisia pollen allergy. Mol. Immunol. 2010, 47, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Razzera, G.; Gadermaier, G.; de Paula, V.; Almeida, M.S.; Egger, M.; Jahn-Schmid, B.; Almeida, F.C.; Ferreira, F.; Valente, A.P. Mapping the interactions between a major pollen allergen and human IgE antibodies. Structure 2010, 18, 1011–1021. [Google Scholar] [CrossRef] [PubMed]
- Leonard, R.; Petersen, B.O.; Himly, M.; Kaar, W.; Wopfner, N.; Kolarich, D.; van Ree, R.; Ebner, C.; Duus, J.O.; Ferreira, F.; et al. Two novel types of O-glycans on the mugwort pollen allergen Art v 1 and their role in antibody binding. J. Biol. Chem. 2005, 280, 7932–7940. [Google Scholar] [CrossRef] [PubMed]
- Jahn-Schmid, B.; Fischer, G.F.; Bohle, B.; Fae, I.; Gadermaier, G.; Dedic, A.; Ferreira, F.; Ebner, C. Antigen presentation of the immunodominant T-cell epitope of the major mugwort pollen allergen, Art v 1, is associated with the expression of HLA-DRB1 *01. J. Allergy Clin. Immunol. 2005, 115, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Jahn-Schmid, B.; Sirven, P.; Leb, V.; Pickl, W.F.; Fischer, G.F.; Gadermaier, G.; Egger, M.; Ebner, C.; Ferreira, F.; Maillere, B.; et al. Characterization of HLA class II/peptide-TCR interactions of the immunodominant T cell epitope in Art v 1, the major mugwort pollen allergen. J. Immunol. 2008, 181, 3636–3642. [Google Scholar] [CrossRef] [PubMed]
- Jahn-Schmid, B.; Kelemen, P.; Himly, M.; Bohle, B.; Fischer, G.; Ferreira, F.; Ebner, C. The T cell response to Art v 1, the major mugwort pollen allergen, is dominated by one epitope. J. Immunol. 2002, 169, 6005–6011. [Google Scholar] [CrossRef] [PubMed]
- Leb, V.M.; Jahn-Schmid, B.; Schmetterer, K.G.; Kueng, H.J.; Haiderer, D.; Neunkirchner, A.; Fischer, G.F.; Nissler, K.; Hartl, A.; Thalhamer, J.; et al. Molecular and functional analysis of the antigen receptor of Art v 1-specific helper T lymphocytes. J. Allergy Clin. Immunol. 2008, 121, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Neunkirchner, A.; Kratzer, B.; Kohler, C.; Smole, U.; Mager, L.F.; Schmetterer, K.G.; Trapin, D.; Leb-Reichl, V.; Rosloniec, E.; Naumann, R.; et al. Genetic restriction of antigen-presentation dictates allergic sensitization and disease in humanized mice. EBioMedicine 2018, 31, 66–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratzer, B.; Kohler, C.; Hofer, S.; Smole, U.; Trapin, D.; Iturri, J.; Pum, D.; Kienzl, P.; Elbe-Burger, A.; Gattinger, P.; et al. Prevention of allergy by virus-like nanoparticles (VNP) delivering shielded versions of major allergens in a humanized murine allergy model. Allergy 2018, 74, 246–260. [Google Scholar] [CrossRef]
- Leonard, R.; Wopfner, N.; Pabst, M.; Stadlmann, J.; Petersen, B.O.; Duus, J.O.; Himly, M.; Radauer, C.; Gadermaier, G.; Razzazi-Fazeli, E.; et al. A new allergen from ragweed (Ambrosia artemisiifolia) with homology to art v 1 from mugwort. J. Biol. Chem. 2010, 285, 27192–27200. [Google Scholar] [CrossRef]
- Pablos, I.; Eichhorn, S.; Briza, P.; Asam, C.; Gartner, U.; Wolf, M.; Ebner, C.; Bohle, B.; Arora, N.; Vieths, S.; et al. Proteomic profiling of the weed feverfew, a neglected pollen allergen source. Sci. Rep. 2017, 7, 6049. [Google Scholar] [CrossRef] [PubMed]
- Pablos, I.; Eichhorn, S.; Machado, Y.; Briza, P.; Neunkirchner, A.; Jahn-Schmid, B.; Wildner, S.; Soh, W.T.; Ebner, C.; Park, J.W.; et al. Distinct epitope structures of defensin-like proteins linked to proline-rich regions give rise to differences in their allergenic activity. Allergy 2018, 73, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Gruber, P.; Gadermaier, G.; Bauer, R.; Weiss, R.; Wagner, S.; Leonard, R.; Breiteneder, H.; Ebner, C.; Ferreira, F.; Egger, M. Role of the polypeptide backbone and post-translational modifications in cross-reactivity of Art v 1, the major mugwort pollen allergen. Biol. Chem. 2009, 390, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.; Kull, S.; Rennert, S.; Becker, W.M.; Krause, S.; Ernst, M.; Gutsmann, T.; Bauer, J.; Lindner, B.; Jappe, U. Peanut defensins: Novel allergens isolated from lipophilic peanut extract. J. Allergy Clin. Immunol. 2015, 136, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Codina, R.; Lockey, R.F.; Fernandez-Caldas, E.; Rama, R. Purification and characterization of a soybean hull allergen responsible for the Barcelona asthma outbreaks. II. Purification and sequencing of the Gly m 2 allergen. Clin. Exp. Allergy 1997, 27, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Sancho, A.I.; Wallner, M.; Hauser, M.; Nagl, B.; Himly, M.; Asam, C.; Ebner, C.; Jahn-Schmid, B.; Bohle, B.; Ferreira, F. T Cell Epitope-Containing Domains of Ragweed Amb a 1 and Mugwort Art v 6 Modulate Immunologic Responses in Humans and Mice. PLoS ONE 2017, 12, e0169784. [Google Scholar] [CrossRef] [PubMed]
- Pichler, U.; Hauser, M.; Wolf, M.; Bernardi, M.L.; Gadermaier, G.; Weiss, R.; Ebner, C.; Yokoi, H.; Takai, T.; Didierlaurent, A.; et al. Pectate lyase pollen allergens: Sensitization profiles and cross-reactivity pattern. PLoS ONE 2015, 10, e0120038. [Google Scholar] [CrossRef]
- Diaz-Perales, A.; Lombardero, M.; Sanchez-Monge, R.; Garcia-Selles, F.J.; Pernas, M.; Fernandez-Rivas, M.; Barber, D.; Salcedo, G. Lipid-transfer proteins as potential plant panallergens: Cross-reactivity among proteins of Artemisia pollen, Castanea nut and Rosaceae fruits, with different IgE-binding capacities. Clin. Exp. Allergy 2000, 30, 1403–1410. [Google Scholar] [CrossRef]
- Gadermaier, G.; Harrer, A.; Girbl, T.; Palazzo, P.; Himly, M.; Vogel, L.; Briza, P.; Mari, A.; Ferreira, F. Isoform identification and characterization of Art v 3, the lipid-transfer protein of mugwort pollen. Mol. Immunol. 2009, 46, 1919–1924. [Google Scholar] [CrossRef]
- Oberhuber, C.; Ma, Y.; Wopfner, N.; Gadermaier, G.; Dedic, A.; Niggemann, B.; Maderegger, B.; Gruber, P.; Ferreira, F.; Scheiner, O.; et al. Prevalence of IgE-binding to Art v 1, Art v 4 and Amb a 1 in mugwort-allergic patients. Int. Arch. Allergy Immunol. 2008, 145, 94–101. [Google Scholar] [CrossRef]
- Wopfner, N.; Willeroidee, M.; Hebenstreit, D.; van Ree, R.; Aalbers, M.; Briza, P.; Thalhamer, J.; Ebner, C.; Richter, K.; Ferreira, F. Molecular and immunological characterization of profilin from mugwort pollen. Biol. Chem. 2002, 383, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Wopfner, N.; Gruber, P.; Wallner, M.; Briza, P.; Ebner, C.; Mari, A.; Richter, K.; Vogel, L.; Ferreira, F. Molecular and immunological characterization of novel weed pollen pan-allergens. Allergy 2008, 63, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Gao, Z.; Gao, L.; Jin, J.; Liu, M.; Sun, Y.; Wu, S.; Wu, L.; Ma, H.; Dong, Y.; et al. Identification of a 62-kDa major allergen from Artemisia pollen as a putative galactose oxidase. Allergy 2018, 73, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Bauer, R.; Himly, M.; Dedic, A.; Ferreira, F.; Thalhamer, J.; Hartl, A. Optimization of codon usage is required for effective genetic immunization against Art v 1, the major allergen of mugwort pollen. Allergy 2003, 58, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Erler, A.; Hawranek, T.; Kruckemeier, L.; Asam, C.; Egger, M.; Ferreira, F.; Briza, P. Proteomic profiling of birch (Betula verrucosa) pollen extracts from different origins. Proteomics 2011, 11, 1486–1498. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, M.L.; Giangrieco, I.; Camardella, L.; Ferrara, R.; Palazzo, P.; Panico, M.R.; Crescenzo, R.; Carratore, V.; Zennaro, D.; Liso, M.; et al. Allergenic lipid transfer proteins from plant-derived foods do not immunologically and clinically behave homogeneously: The kiwifruit LTP as a model. PLoS ONE 2011, 6, e27856. [Google Scholar] [CrossRef]
- Jahn-Schmid, B.; Hauser, M.; Wopfner, N.; Briza, P.; Berger, U.E.; Asero, R.; Ebner, C.; Ferreira, F.; Bohle, B. Humoral and cellular cross-reactivity between Amb a 1, the major ragweed pollen allergen, and its mugwort homolog Art v 6. J. Immunol. 2012, 188, 1559–1567. [Google Scholar] [CrossRef]
- Wolf, M.; Twaroch, T.E.; Huber, S.; Reithofer, M.; Steiner, M.; Aglas, L.; Hauser, M.; Aloisi, I.; Asam, C.; Hofer, H.; et al. Amb a 1 isoforms: Unequal siblings with distinct immunological features. Allergy 2017, 72, 1874–1882. [Google Scholar] [CrossRef] [Green Version]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatr. Allergy Immunol. 2016, 27, 1–250. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Common Name | Special Use |
|---|---|---|
| A. absinthium | absinthe wormwood, wormwood, green ginger, or grand wormwood | medicinal herb, ingredient of the liqueur “absinthe” |
| A. annua | sweet wormwood, sweet sage wort, or annual wormwood | traditional Chinese herbal medicine; leaves are used for antimalaria treatment (artemisinin) |
| A. californica | California sagebrush | spices and tea |
| A. frigida | sagebrush, prairie sage wort, or fringed wormwood | medicinal herb containing camphor |
| A. ludoviciana | silver wormwood or white sagebrush | medicinal herb |
| A. tridentata | common sagebrush or mountain sagebrush | traditional medicine of North American Indian tribes |
| A. vulgaris | mugwort or common wormwood | traditional Chinese herbal medicine |
| % MI (SD) 0.005 µg/mL nArt v 1 | % MI (SD) 0.05 µg/mL nArt v 1 | % MI (SD) 0.5 µg/mL nArt v 1 | % MI (SD) 5.0 µg/mL nArt v 1 | |
|---|---|---|---|---|
| A. absinthium | 70.4 (20.4) | 75.8 (17.9) | 80.5 (11.7) | 85.3 (6.8) |
| A. annua | 75.2 (17.5) | 81.5 (16.3) | 85.2 (9.3) | 90.2 (5.8) |
| A. californica | 72.8 (15.6) | 80.0 (15.1) | 90.5 (8.4) | 90.5 (5.2) |
| A. frigida | 74.3 (16.8) | 82.5 (12.7) | 86.7 (7.2) | 90.2 (5.4) |
| A. ludoviciana | 69.5 (21.2) | 75.6 (19.4) | 81.7 (10.3) | 87.3 (6.2) |
| A. tridentata | 73.9 (17.0) | 80.0 (14.9) | 84.1 (10.9) | 87.2 (8.0) |
| A. vulgaris | 63.8 (21.2) | 76.4 (17.9) | 78.6 (10.6) | 82.1 (6.6) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pablos, I.; Egger, M.; Vejvar, E.; Reichl, V.; Briza, P.; Zennaro, D.; Rafaiani, C.; Pickl, W.; Bohle, B.; Mari, A.; et al. Similar Allergenicity to Different Artemisia Species Is a Consequence of Highly Cross-Reactive Art v 1-Like Molecules. Medicina 2019, 55, 504. https://doi.org/10.3390/medicina55080504
Pablos I, Egger M, Vejvar E, Reichl V, Briza P, Zennaro D, Rafaiani C, Pickl W, Bohle B, Mari A, et al. Similar Allergenicity to Different Artemisia Species Is a Consequence of Highly Cross-Reactive Art v 1-Like Molecules. Medicina. 2019; 55(8):504. https://doi.org/10.3390/medicina55080504
Chicago/Turabian StylePablos, Isabel, Matthias Egger, Eva Vejvar, Victoria Reichl, Peter Briza, Danila Zennaro, Chiara Rafaiani, Winfried Pickl, Barbara Bohle, Adriano Mari, and et al. 2019. "Similar Allergenicity to Different Artemisia Species Is a Consequence of Highly Cross-Reactive Art v 1-Like Molecules" Medicina 55, no. 8: 504. https://doi.org/10.3390/medicina55080504