Modulation of Renal Insulin Signaling Pathway and Antioxidant Enzymes with Streptozotocin-Induced Diabetes: Effects of Resveratrol

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Treatments
2.2. Tissue Homogenization and Measurement of Renal Malondialdehyde (MDA) Contents
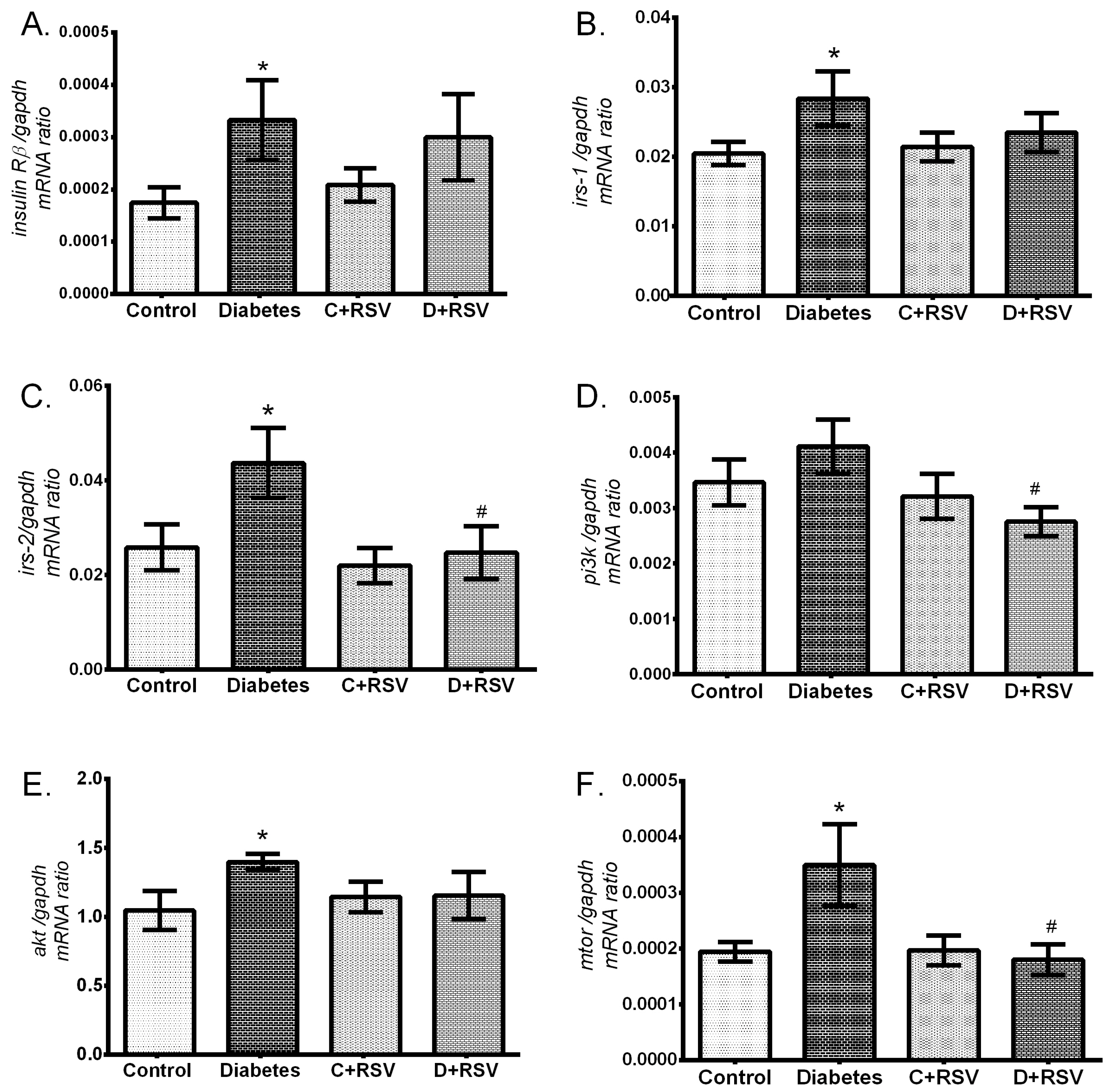
2.3. Determination of Gene Expressions of Antioxidant Enzymes, Insulin Signaling Pathway Components, and nfκb with Real-Time Polymerase Chain Reaction
2.4. Immunoblot Analysis of Antioxidant Enzymes; CAT, SOD1, SOD2 and Insulin Signaling Pathway Components; Insulin Rβ, PI3K, AKT1
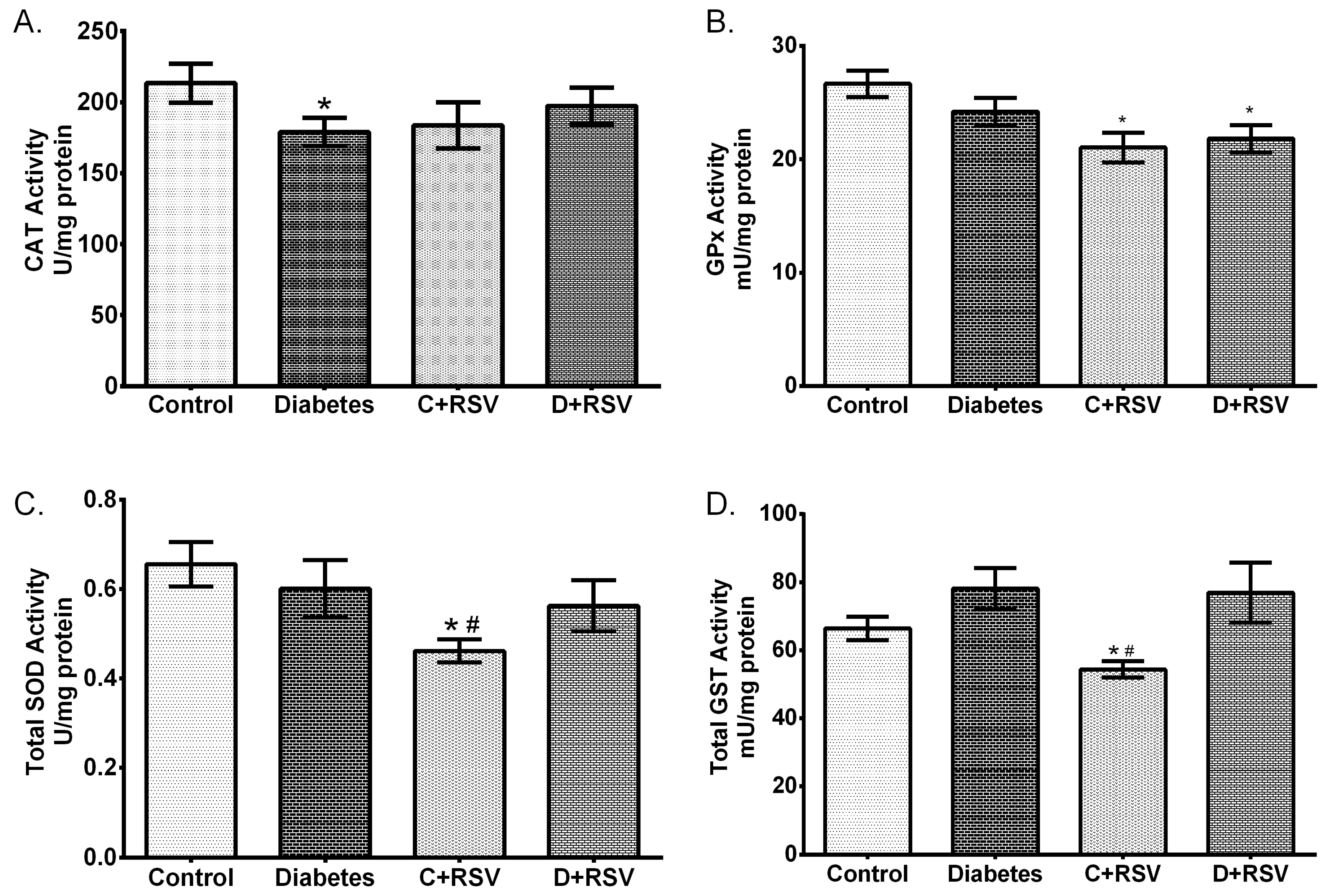
2.5. Determination of Antioxidant Enzyme Activities
2.6. Statistical Analysis
3. Results
3.1. Effects of Diabetes and/or Resveratrol on Some Metabolic Parameters
3.2. Changes in Gene Expression Levels of Antioxidant Enzymes
3.3. Changes in Protein Expression of Antioxidant Enzymes
3.4. Regulation of Renal Insulin Signaling Pathway Components
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850–867. [Google Scholar] [CrossRef]
- Okur, M.E.; Karantas, I.D.; Siafaka, P.I. Diabetes mellitus: A review on pathophysiology, current status of oral medications and future perspectives. Acta Pharm. Sci. 2017, 55, 61–82. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, oxidative stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Sadi, G.; Konat, D. Resveratrol regulates oxidative biomarkers and antioxidant enzymes in the brain of streptozotocin-induced diabetic rats. Pharm. Biol. 2016, 54. [Google Scholar] [CrossRef] [PubMed]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed]
- Pektaş, A.; Pektaş, M.B.; Koca, H.B.; Tosun, M.; Aslan, E.; Koca, S.; Sadi, G. Effects of resveratrol on diabetes-induced vascular tissue damage and inflammation in male rats. Turkish J. Biochem. 2017, 42. [Google Scholar] [CrossRef]
- Espinosa-Diez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sánchez-Pérez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox. Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-M.; Johnson, J.A. An important role of Nrf2-ARE pathway in the cellular defense mechanism. J. Biochem. Mol. Biol. 2004, 37, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Soufi, F.G.; Mohammad-Nejad, D.; Ahmadieh, H. Resveratrol improves diabetic retinopathy possibly through oxidative stress - nuclear factor κB - apoptosis pathway. Pharmacol. Rep. 2012, 64, 1505–1514. [Google Scholar] [CrossRef]
- Sadi, G.; Bozan, D.; Yildiz, H.B. Redox regulation of antioxidant enzymes: Post-translational modulation of catalase and glutathione peroxidase activity by resveratrol in diabetic rat liver. Mol. Cell Biochem. 2014, 393, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Szkudelska, K.; Szkudelski, T. Resveratrol, obesity and diabetes. Eur. J. Pharmacol. 2010, 635, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hamadi, N.; Mansour, A.; Hassan, M.H.; Khalifi-Touhami, F.; Badary, O. Ameliorative effects of resveratrol on liver injury in streptozotocin-induced diabetic rats. J. Biochem. Mol. Toxicol. 2012, 26, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Koca, H.; Pektas, M.; Koca, S.; Pektas, G.; Sadi, G. Diabetes-induced renal failure is associated with tissue inflammation and neutrophil gelatinase-associated lipocalin: Effects of resveratrol. Arch. Biol. Sci. 2016, 68, 747–752. [Google Scholar] [CrossRef]
- Schmatz, R.; Perreira, L.B.; Stefanello, N.; Mazzanti, C.; Spanevello, R.; Gutierres, J.; Bagatini, M.; Martins, C.C.; Abdalla, F.H.; da Silva Serres, J.D.; et al. Effects of resveratrol on biomarkers of oxidative stress and on the activity of delta aminolevulinic acid dehydratase in liver and kidney of streptozotocin-induced diabetic rats. Biochimie 2012, 94, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Pecoits-Filho, R.; Abensur, H.; Betônico, C.C.R.; Machado, A.D.; Parente, E.B.; Queiroz, M.; Salles, J.E.; Titan, S.; Vencio, S. Interactions between kidney disease and diabetes: Dangerous liaisons. Diabetol. Metab. Syndr. 2016, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Sadi, G.; Pektaş, M.B.; Koca, H.B.; Tosun, M.; Koca, T. Resveratrol improves hepatic insulin signaling and reduces the inflammatory response in streptozotocin-induced diabetes. Gene 2015, 570, 213–220. [Google Scholar] [CrossRef]
- Lowry, O.; Roserbrough, N.; Farr, A.; Randall, R. Protein measurement with folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Aebi, H. Catalase in Vitro. In Methods in Enzymology; Academy Press: New York, NY, USA, 1984; pp. 121–126. [Google Scholar]
- Paglia, D.; Valentine, W. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione, S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, G.; Singh, A.K.; Kumar, A.; Prakash, O.; Singh, M.P. Resveratrol modulates pyrogallol-induced changes in hepatic toxicity markers, xenobiotic metabolizing enzymes and oxidative stress. Eur. J. Pharmacol. 2008, 596, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, B.; Du, L.; Chen, J.; Lu, Q. Resveratrol ameliorates cadmium induced renal oxidative damage and inflammation. Int. J. Clin. Exp. Med. 2017, 10, 7563–7572. [Google Scholar]
- Palsamy, P.; Subramanian, S. Resveratrol protects diabetic kidney by attenuating hyperglycemia-mediated oxidative stress and renal inflammatory cytokines via Nrf2-Keap1 signaling. Biochim. Biophys. Acta 2011, 1812, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.-S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NFkappaB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Sainz, R.M.; Antoli, I.; Herrera, F.; Martin, V.; Rodriguez, C. Melatonin regulation of antioxidant enzyme gene expression. Cell Mol. Life Sci. 2002, 59, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Strehlow, K.; Rotter, S.; Wassmann, S.; Adam, O.; Grohé, C.; Laufs, K.; Böhm, M.; Nickenig, G. Modulation of Antioxidant Enzyme Expression and Function by Estrogen. Circ. Res. 2003, 93, 170–177. [Google Scholar] [CrossRef]
- Lizcano, J.M.; Alessi, D.R. The insulin signalling pathway. Curr. Biol. 2002, 12, R236–R238. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| cat | GCGAATGGAGAGGCAGTGTAC | GAGTGACGTTGTCTTCATTAGCACTG |
| gpx1 | CCACCACCGGGTCGGACATAC | CTCTCCGCGGTGGCACAGT |
| sod1 | TAGCAGGACAGCAGATGAGT | GCAGAAGGCAAGCGGTGAAC |
| sod2 | GCACATTAACGCGCAGATCA | AGCCTCCAGCAACTCTCCTT |
| gst-mu | AGAAGCAGAAGCCAGAGTTC | GGGGTGAGGTTGAGGAGATG |
| nfκb | GGGTCAGAGGCCAATAGAGA | CCTAGCTTTCTCTGAACTGCAAA |
| insulin rβ | GTGCTGCTCATGTCCTTAGA | AATGGTCTGTGCTCTTCGTG |
| irs1 | GCCAATCTTCATCCAGTTGC | CATCGTGAAGAAGGCATAGG |
| irs2 | CTACCCACTGAGCCCAAGAG | CCAGGGATGAAGCAGGACTA |
| pi3k | ATGCAACTGCCTTGCACATT | CGCCTGAAGCTGAGCAACAT |
| akt1 | GAAGAAGAGCTCGCCTCCAT | GAAGGAGAAGGCCACAGGTC |
| mtor | GCAATGGGCACGAGTTTGTT | AGTGTGTTCACCAGGCCAAA |
| gapdh | TCCTTGGAGGCCATGTGGGCCAT | TGATGACATCAAGAAGGTGGTGAAG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadi, G.; Şahin, G.; Bostanci, A. Modulation of Renal Insulin Signaling Pathway and Antioxidant Enzymes with Streptozotocin-Induced Diabetes: Effects of Resveratrol. Medicina 2019, 55, 3. https://doi.org/10.3390/medicina55010003
Sadi G, Şahin G, Bostanci A. Modulation of Renal Insulin Signaling Pathway and Antioxidant Enzymes with Streptozotocin-Induced Diabetes: Effects of Resveratrol. Medicina. 2019; 55(1):3. https://doi.org/10.3390/medicina55010003
Chicago/Turabian StyleSadi, Gökhan, Gamze Şahin, and Aykut Bostanci. 2019. "Modulation of Renal Insulin Signaling Pathway and Antioxidant Enzymes with Streptozotocin-Induced Diabetes: Effects of Resveratrol" Medicina 55, no. 1: 3. https://doi.org/10.3390/medicina55010003