Alternans in atria: Mechanisms and clinical relevance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Putative mechanisms of alternans
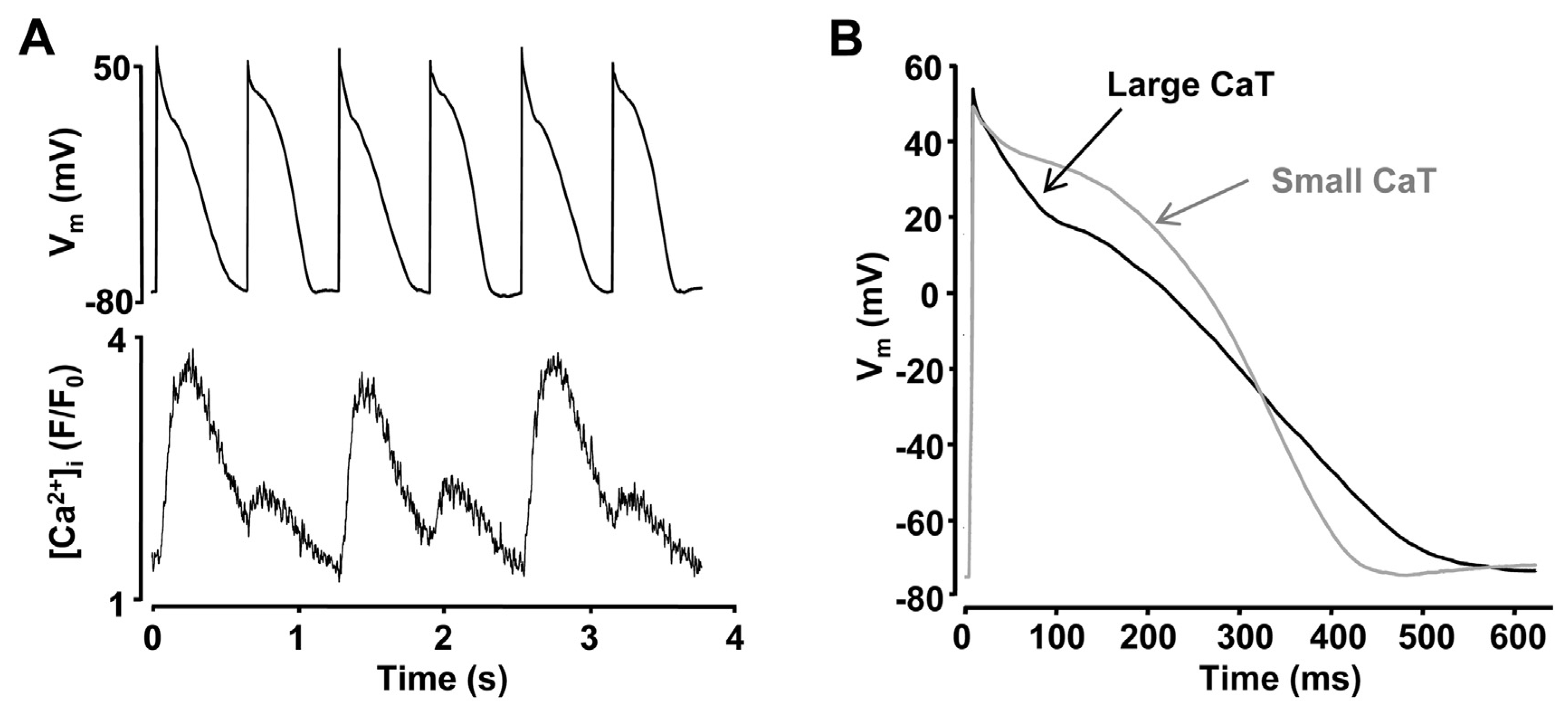
2.1 Vm as key mechanism for the development of alternans
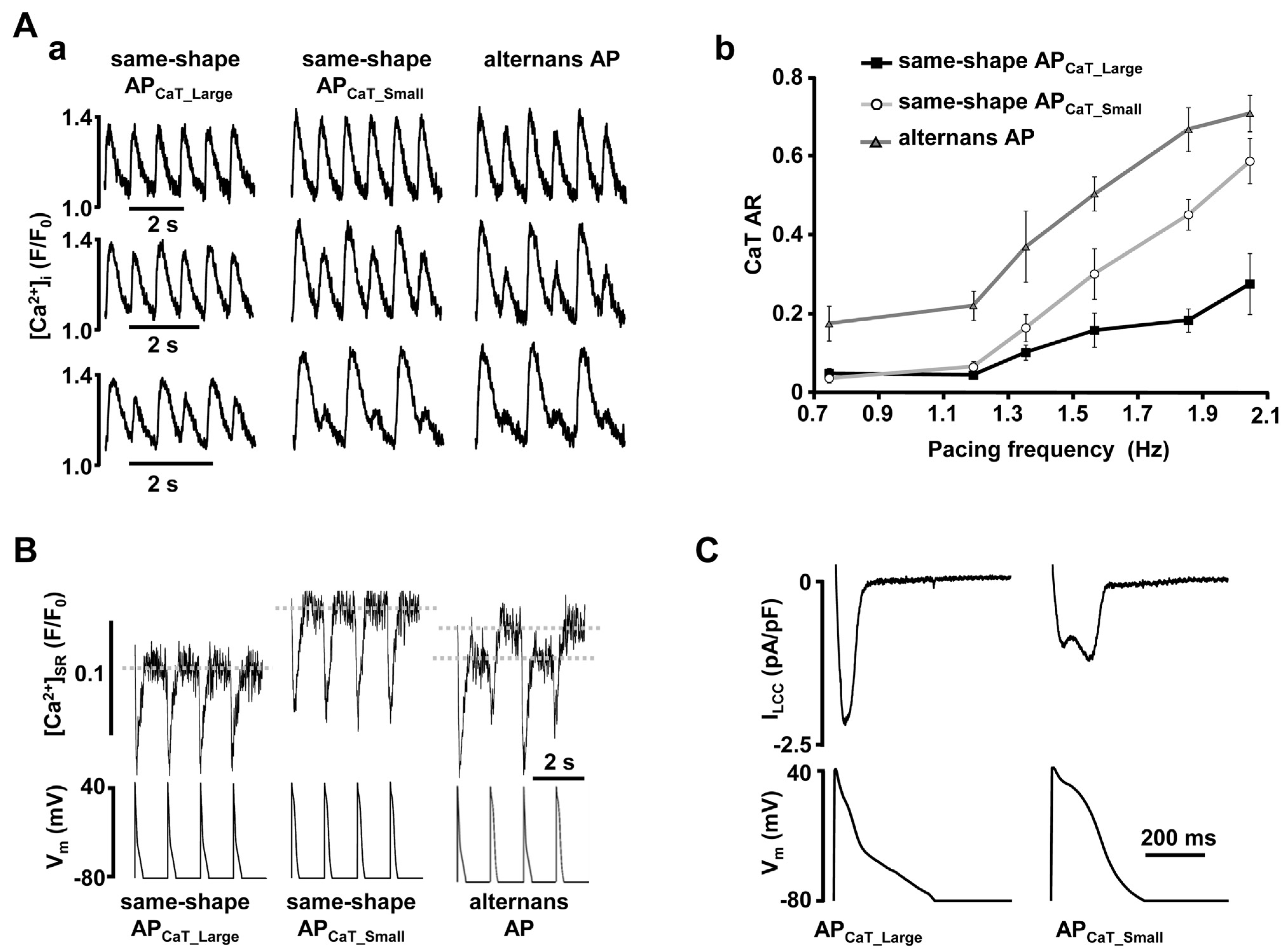
2.2 Intracellular Ca2+ cycling as an underlying mechanism of alternans
3. Differences between atrial and ventricular alternans
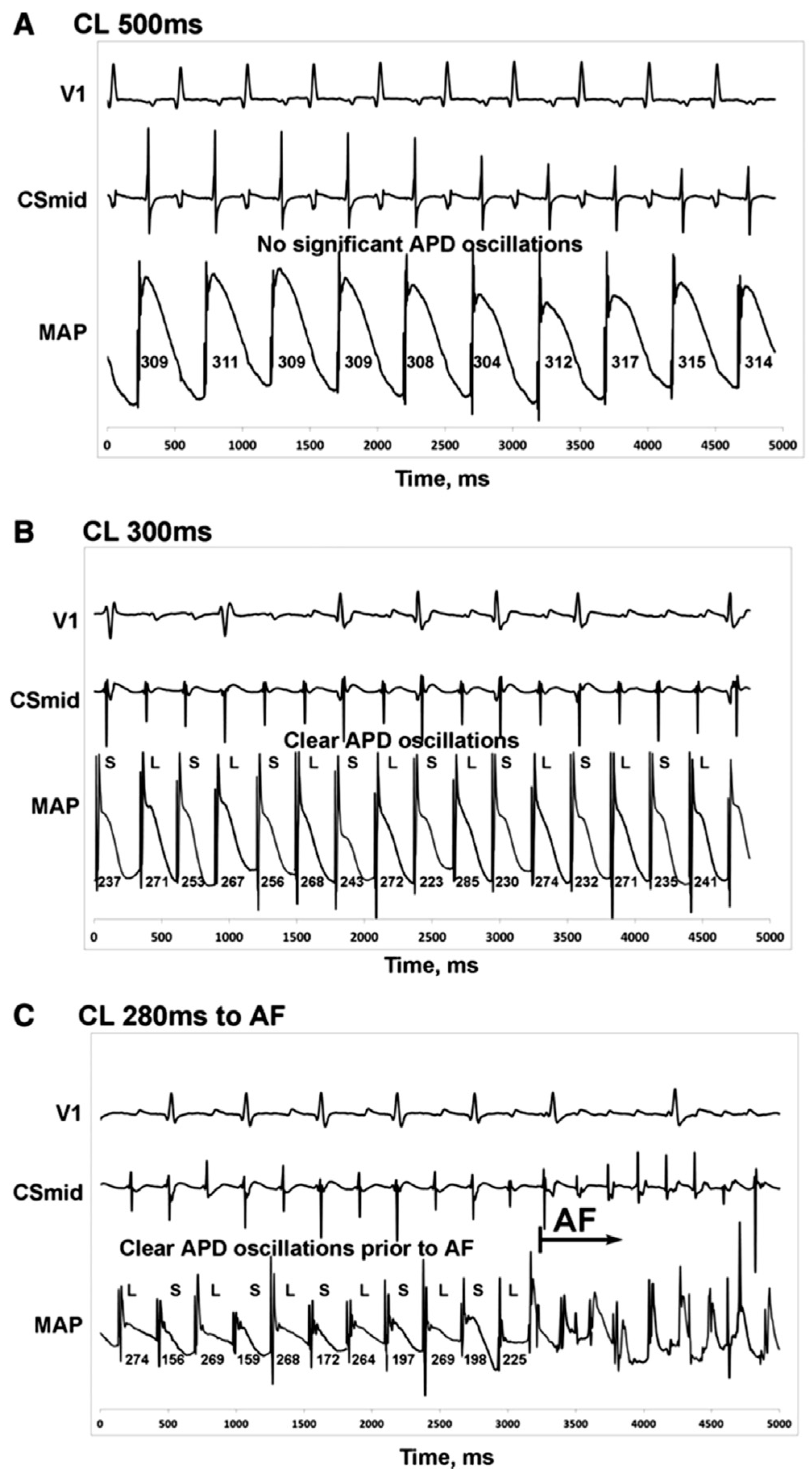
4. Clinical relevance of cardiac alternans
5. Mechanisms of spatially discordant alternans
5.1. Calcium cycling heterogeneity
5.2. Insufficient cell-to-cell coupling
5.3. Conduction velocity and APD restitution
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Chugh, SS; Havmoeller, R; Narayanan, K; Singh, D; Rienstra, M; Benjamin, EJ; et al. Worldwide epidemiology of atrial fibrillation: a Global Burden of Disease 2010 Study. Circulation 2014, 129, 837–47. [Google Scholar] [CrossRef] [PubMed]
- Chinitz, JS; Vaishnava, P; Narayan, RL; Fuster, V. Atrial fibrillation through the years: contemporary evaluation and management. Circulation 2013, 127, 408–16. [Google Scholar] [CrossRef] [PubMed]
- Comtois, P; Nattel, S. Atrial repolarization alternans as a path to atrial fibrillation. J Cardiovasc Electrophysiol 2012, 23, 1013–5. [Google Scholar] [CrossRef] [PubMed]
- Berenfeld, O; Jalife, J. Mechanisms of atrial fibrillation: rotors, ionic determinants, and excitation frequency. Cardiol Clin 2014, 32, 495–506. [Google Scholar]
- Kottkamp, H. Human atrial fibrillation substrate: towards a specific fibrotic atrial cardiomyopathy. Eur Heart J 2013, 34, 2731–8. [Google Scholar] [CrossRef] [PubMed]
- Franz, MR; Jamal, SM; Narayan, SM. The role of action potential alternans in the initiation of atrial fibrillation in humans: a review and future directions. Europace 2012, 14 (Suppl 5), v58–64. [Google Scholar] [CrossRef] [PubMed]
- Hiromoto, K; Shimizu, H; Furukawa, Y; Kanemori, T; Mine, T; Masuyama, T; et al. Discordant repolarization alternans-induced atrial fibrillation is suppressed by verapamil. Circ J 2005, 69, 1368–73. [Google Scholar] [CrossRef] [PubMed]
- Narayan, SM; Bode, F; Karasik, PL; Franz, MR. Alternans of atrial action potentials during atrial flutter as a precursor to atrial fibrillation. Circulation 2002, 106, 1968–73. [Google Scholar] [CrossRef] [PubMed]
- Coyne, KS; Paramore, C; Grandy, S; Mercader, M; Reynolds, M; Zimetbaum, P. Assessing the direct costs of treating nonvalvular atrial fibrillation in the United States. Value Health 2006, 9, 348–56. [Google Scholar] [CrossRef] [PubMed]
- Kim, BS; Kim, YH; Hwang, GS; Pak, HN; Lee, SC; Shim, WJ; et al. Action potential duration restitution kinetics in human atrial fibrillation. J Am Coll Cardiol 2002, 39, 1329–36. [Google Scholar] [CrossRef]
- Narayan, SM; Franz, MR; Clopton, P; Pruvot, EJ; Krummen, DE. Repolarization alternans reveals vulnerability to human atrial fibrillation. Circulation 2011, 123, 2922–30. [Google Scholar] [CrossRef] [PubMed]
- Jousset, F; Tenkorang, J; Vesin, JM; Pascale, P; Ruchat, P; Rollin, AG; et al. Kinetics of atrial repolarization alternans in a free-behaving ovine model. J Cardiovasc Electrophysiol 2012, 23, 1003–12. [Google Scholar] [CrossRef] [PubMed]
- Gaeta, SA; Christini, DJ. Non-linear dynamics of cardiac alternans: subcellular to tissue-level mechanisms of arrhythmia. Front Physiol 2012, 3, 157. [Google Scholar] [CrossRef] [PubMed]
- Myles, RC; Burton, FL; Cobbe, SM; Smith, GL. The link between repolarisation alternans and ventricular arrhythmia: does the cellular phenomenon extend to the clinical problem? J Mol Cell Cardiol 2008, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Traube, L. Ein fall von pulsus bigeminus nebst bemerkungen tiber die lebershwellungen bei klappenfehlern und uber acute leberatrophic. Ber Klin Wschr 1872, 185. [Google Scholar]
- Lewis, T. Notes upon alternation of the heart. Quart J Med 1911, 141–4. [Google Scholar]
- Kanaporis, G; Blatter, LA. The mechanisms of calcium cycling and action potential dynamics in cardiac alternans. Circ Res 2015, 116, 846–56. [Google Scholar] [CrossRef] [PubMed]
- Pruvot, EJ; Katra, RP; Rosenbaum, DS; Laurita, KR. Role of calcium cycling versus restitution in the mechanism of repolarization alternans. Circ Res 2004, 94, 1083–90. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, MA; Fenton, FH; Evans, SJ; Hastings, HM; Karma, A. Mechanisms for discordant alternans. J Cardiovasc Electrophysiol 2001, 12, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw, Y; Sato, D; Karma, A. Coupled dynamics of voltage and calcium in paced cardiac cells. Phys Rev E Stat Nonlin Soft Matter Phys 2005, 71, 021903. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, MA; Koller, ML. Mathematical analysis of dynamics of cardiac memory and accommodation: theory and experiment. Am J Physiol Heart Circ Physiol 2002, 282, H1534–47. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z; Garfinkel, A; Chen, PS; Weiss, JN. Mechanisms of discordant alternans and induction of reentry in simulated cardiac tissue. Circulation 2002, 102, 1664–70. [Google Scholar] [CrossRef]
- Tao, T; O’Neill, SC; Diaz, ME; Li, YT; Eisner, DA; Zhang, H. Alternans of cardiac calcium cycling in a cluster of ryanodine receptors: a simulation study. Am J Physiol Heart Circ Physiol 2008, 295, H598–609. [Google Scholar] [CrossRef] [PubMed]
- Pawlak Cieslik, A; Szturmowicz, M; Fijalkowska, A; Gatarek, J; Gralec, R; Blasinska-Przerwa, K; et al. Diagnosis of malignant pericarditis: a single centre experience. Kardiol Pol 2012, 70, 1147–53. [Google Scholar] [PubMed]
- Nolasco, JB; Dahlen, RW. A graphic method for the study of alternation in cardiac action potentials. J Appl Physiol 1968, 25, 191–6. [Google Scholar] [CrossRef] [PubMed]
- Tolkacheva, EG; Romeo, MM; Guerraty, M; Gauthier, DJ. Condition for alternans and its control in a two-dimensional mapping model of paced cardiac dynamics. Phys Rev E Stat Nonlin Soft Matter Phys 2004, 69, 031904. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw, Y; Karma, A. Turing instability mediated by voltage and calcium diffusion in paced cardiac cells. Proc Natl Acad Sci U S A 2006, 103, 5670–5. [Google Scholar] [CrossRef] [PubMed]
- Jordan, PN; Christini, DJ. Characterizing the contribution of voltage- and calcium-dependent coupling to action potential stability: implications for repolarization alternans. Am J Physiol Heart Circ Physiol 2007, 293, H2109–18. [Google Scholar] [CrossRef] [PubMed]
- Jordan, PN; Christini, DJ. Action potential morphology influences intracellular calcium handling stability and the occurrence of alternans. Biophys J 2006, 90, 672–80. [Google Scholar] [CrossRef] [PubMed]
- Koller, ML; Maier, SK; Gelzer, AR; Bauer, WR; Meesmann, M; Gilmour, RF, Jr. Altered dynamics of action potential restitution and alternans in humans with structural heart disease. Circulation 2005, 112, 1542–8. [Google Scholar] [CrossRef] [PubMed]
- Tolkacheva, EG; Anumonwo, JM; Jalife, J. Action potential duration restitution portraits of mammalian ventricular myocytes: role of calcium current. Biophys J 2006, 91, 2735–45. [Google Scholar] [CrossRef] [PubMed]
- Bayer, JD; Narayan, SM; Lalani, GG; Trayanova, NA. Rate-dependent action potential alternans in human heart failure implicates abnormal intracellular calcium handling. Heart Rhythm 2010, 7, 1093–101. [Google Scholar] [CrossRef] [PubMed]
- Cherry, EM; Hastings, HM; Evans, SJ. Dynamics of human atrial cell models: restitution, memory, and intracellular calcium dynamics in single cells. Prog Biophys Mol Biol 2008, 98, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Kalb, SS; Dobrovolny, HM; Tolkacheva, EG; Idriss, SF; Krassowska, W; Gauthier, DJ. The restitution portrait: a new method for investigating rate-dependent restitution. J Cardiovasc Electrophysiol 2004, 15, 698–709. [Google Scholar]
- Pruvot, EJ; Katra, RP; Rosenbaum, DS; Laurita, KR. Role of calcium cycling versus restitution in the mechanism of repolarization alternans. Circ Res 2004, 94, 1083–90. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, H; Bailey, JC; Surawicz, B. Alternans of action potential duration after abrupt shortening of cycle length: differences between dog Purkinje and ventricular muscle fibers. Circ Res 1988, 62, 1027–40. [Google Scholar] [CrossRef] [PubMed]
- Banville, I; Gray, RA. Effect of action potential duration and conduction velocity restitution and their spatial dispersion on alternans and the stability of arrhythmias. J Cardiovasc Electrophysiol 2002, 13, 1141–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, R; Patwardhan, A. Mechanism of repolarization alternans has restitution of action potential duration dependent and independent components. J Cardiovasc Electrophysiol 2006, 17, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Sah, R; Ramirez, RJ; Kaprielian, R; Backx, PH. Alterations in action potential profile enhance excitation-contraction coupling in rat cardiac myocytes. J Physiol 2001, 533, 201–14. [Google Scholar] [CrossRef] [PubMed]
- Bassani, RA; Altamirano, J; Puglisi, JL; Bers, DM. Action potential duration determines sarcoplasmic reticulum Ca2+ reloading in mammalian ventricular myocytes. J Physiol 2004, 559, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Livshitz, LM; Rudy, Y. Regulation of Ca2+ and electrical alternans in cardiac myocytes: role of CAMKII and repolarizing currents. Am J Physiol Heart Circ Physiol 2007, 292, H2854–66. [Google Scholar] [CrossRef] [PubMed]
- Wan, X; Cutler, M; Song, Z; Karma, A; Matsuda, T; Baba, A; et al. New experimental evidence for mechanism of arrhythmogenic membrane potential alternans based on balance of electrogenic I(NCX)/I(Ca) currents. Heart Rhythm 2012, 9, 1698–705. [Google Scholar] [CrossRef] [PubMed]
- Weber, CR; Piacentino, V, 3rd; Ginsburg, KS; Houser, SR; Bers, DM. Na+-Ca2+ exchange current and submembrane [Ca2+] during the cardiac action potential. Circ Res 2002, 90, 182–9. [Google Scholar] [CrossRef] [PubMed]
- Guo, D; Zhao, X; Wu, Y; Liu, T; Kowey, PR; Yan, GX. L-type calcium current reactivation contributes to arrhythmogenesis associated with action potential triangulation. J Cardiovasc Electrophysiol 2007, 18, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Clark, RB; Bouchard, RA; Giles, WR. Action potential duration modulates calcium influx, Na+-Ca2+ exchange, and intracellular calcium release in rat ventricular myocytes. Ann N Y Acad Sci 1996, 779, 417–29. [Google Scholar] [CrossRef] [PubMed]
- Linz, KW; Meyer, R. Profile and kinetics of L-type calcium current during the cardiac ventricular action potential compared in guinea-pigs, rats and rabbits. Pflugers Arch 2000, 439, 588–99. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z; Xie, Y; Garfinkel, A; Weiss, JN. T-wave alternans and arrhythmogenesis in cardiac diseases. Front Physiol 2010, 1, 154. [Google Scholar] [CrossRef] [PubMed]
- Franz, MR. The electrical restitution curve revisited: steep or flat slope – which is better? J Cardiovasc Electrophysiol 2003, 14, S140–7. [Google Scholar] [CrossRef] [PubMed]
- Lau, E; Kossidas, K; Kim, TY; Kunitomo, Y; Ziv, O; Song, Z; et al. spatially discordant alternans and arrhythmias in tachypacing-induced cardiac myopathy in transgenic LQT1 rabbits: the importance of IKs and Ca2+ cycling. PLoS One 2015, 10, e0122754. [Google Scholar]
- Goldhaber, JI; Xie, LH; Duong, T; Motter, C; Khuu, K; Weiss, JN. Action potential duration restitution and alternans in rabbit ventricular myocytes: the key role of intracellular calcium cycling. Circ Res 2005, 96, 459–66. [Google Scholar] [CrossRef] [PubMed]
- Chudin, E; Goldhaber, J; Garfinkel, A; Weiss, J; Kogan, B. Intracellular Ca2+ dynamics and the stability of ventricular tachycardia. Biophys J 1999, 77, 2930–41. [Google Scholar] [CrossRef]
- Kanaporis, G; Blatter, LA. Membrane potential determines calcium alternans through modulation of SR Ca2+ load and L-type Ca2+ current. J Mol Cell Cardiol 2017, 105, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Diaz, ME; O’Neill, SC; Eisner, DA. Sarcoplasmic reticulum calcium content fluctuation is the key to cardiac alternans. Circ Res 2004, 94, 650–6. [Google Scholar] [CrossRef] [PubMed]
- Shkryl, VM; Maxwell, JT; Domeier, TL; Blatter, LA. Refractoriness of sarcoplasmic reticulum Ca2+ release determines Ca2+ alternans in atrial myocytes. Am J Physiol Heart Circ Physiol 2012, 302, H2310–2. [Google Scholar] [CrossRef] [PubMed]
- Shannon, TR; Ginsburg, KS; Bers, DM. Potentiation of fractional sarcoplasmic reticulum calcium release by total and free intra-sarcoplasmic reticulum calcium concentration. Biophys J 2000, 78, 334–43. [Google Scholar] [CrossRef]
- Li, Y; Diaz, ME; Eisner, DA; O’Neill, S. The effects of membrane potential, SR Ca2+ content and RyR responsiveness on systolic Ca2+ alternans in rat ventricular myocytes. J Physiol 2009, 587, 1283–92. [Google Scholar]
- Huser, J; Wang, YG; Sheehan, KA; Cifuentes, F; Lipsius, SL; Blatter, LA. Functional coupling between glycolysis and excitation-contraction coupling underlies alternans in cat heart cells. J Physiol 2000, 524 (Pt 3), 795–806. [Google Scholar] [CrossRef]
- Picht, E; DeSantiago, J; Blatter, LA; Bers, DM. Cardiac alternans do not rely on diastolic sarcoplasmic reticulum calcium content fluctuations. Circ Res 2006, 99, 740–8. [Google Scholar] [CrossRef] [PubMed]
- Shkryl, VM; Maxwell, JT; Domeier, TL; Blatter, LA. Refractoriness of sarcoplasmic reticulum Ca release determines Ca alternans in atrial myocytes. Am J Physiol Heart Circ Physiol 2012, 302, H2310–2. [Google Scholar] [CrossRef] [PubMed]
- Wang, L; Myles, RC; De Jesus, NM; Ohlendorf, AK; Bers, DM; Ripplinger, CM. Optical mapping of sarcoplasmic reticulum Ca2+ in the intact heart: ryanodine receptor refractoriness during alternans and fibrillation. Circ Res 2014, 114, 1410–21. [Google Scholar] [CrossRef] [PubMed]
- Edwards, JN; Blatter, LA. Cardiac alternans and intracellular calcium cycling. Clin Exp Pharmacol Physiol 2014, 41, 524–32. [Google Scholar] [CrossRef] [PubMed]
- Boknik, P; Unkel, C; Kirchhefer, U; Kleideiter, U; Klein-Wiele, O; Knapp, J; et al. Regional expression of phospholamban in the human heart. Cardiovasc Res 1999, 43, 67–76. [Google Scholar] [CrossRef]
- Vangheluwe, P; Schuermans, M; Zador, E; Waelkens, E; Raeymaekers, L; Wuytack, F. Sarcolipin and phospholamban mRNA and protein expression in cardiac and skeletal muscle of different species. Biochem J 2005, 389, 151–9. [Google Scholar]
- Luss, I; Boknik, P; Jones, LR; Kirchhefer, U; Knapp, J; Linck, B; et al. Expression of cardiac calcium regulatory proteins in atrium v ventricle in different species. J Mol Cell Cardiol 1999, 31, 1299–314. [Google Scholar] [CrossRef] [PubMed]
- Fox, JJ; McHarg, JL; Gilmour, RF, Jr. Ionic mechanism of electrical alternans. Am J Physiol Heart Circ Physiol 2002, 282, H516–30. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw, Y; Watanabe, MA; Garfinkel, A; Weiss, JN; Karma, A. Model of intracellular calcium cycling in ventricular myocytes. Biophys J 2003, 85, 3666–86. [Google Scholar] [CrossRef]
- Wan, X; Laurita, KR; Pruvot, EJ; Rosenbaum, DS. Molecular correlates of repolarization alternans in cardiac myocytes. J Mol Cell Cardiol 2005, 39, 419–28. [Google Scholar] [CrossRef] [PubMed]
- Lugo, CA; Cantalapiedra, IR; Penaranda, A; Hove-Madsen, L; Echebarria, B. Are SR Ca content fluctuations or SR refractoriness the key to atrial cardiac alternans?: insights from a human atrial model. Am J Physiol Heart Circ Physiol 2014, 306, H1540–52. [Google Scholar] [CrossRef] [PubMed]
- Ellis, WS; SippensGroenewegen, A; Auslander, DM; Lesh, MD. The role of the crista terminalis in atrial flutter and fibrillation: a computer modeling study. Ann Biomed Eng 2000, 28, 742–54. [Google Scholar] [CrossRef] [PubMed]
- Bootman, MD; Smyrnias, I; Thul, R; Coombes, S; Roderick, HL. Atrial cardiomyocyte calcium signalling. Biochim Biophys Acta 2011, 1813, 922–34. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, KA; Zima, AV; Blatter, LA. Regional differences in spontaneous Ca2+ spark activity and regulation in cat atrial myocytes. J Physiol 2006, 572, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Blatter, LA; Kockskamper, J; Sheehan, KA; Zima, AV; Huser, J; Lipsius, SL. Local calcium gradients during excitation– contraction coupling and alternans in atrial myocytes. J Physiol 2003, 546, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Li, Q; O’Neill, SC; Tao, T; Li, Y; Eisner, D; Zhang, H. Mechanisms by which cytoplasmic calcium wave propagation and alternans are generated in cardiac atrial myocytes lacking T-tubules-insights from a simulation study. Biophys J 2012, 102, 1471–82. [Google Scholar] [CrossRef] [PubMed]
- Kockskamper, J; Blatter, LA. Subcellular Ca2+ alternans represents a novel mechanism for the generation of arrhythmogenic Ca2+ waves in cat atrial myocytes. J Physiol 2002, 545, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Eisner, DA; Li, Y; O’Neill, SC. Alternans of intracellular calcium: mechanism and significance. Heart Rhythm 2006, 3, 743–5. [Google Scholar] [CrossRef] [PubMed]
- Stary, V; Puppala, D; Scherrer-Crosbie, M; Dillmann, WH; Armoundas, AA. SERCA2a upregulation ameliorates cellular alternans induced by metabolic inhibition. J Appl Physiol (1985) 2016, 120, 865–75. [Google Scholar] [CrossRef] [PubMed]
- Cutler, MJ; Wan, X; Laurita, KR; Hajjar, RJ; Rosenbaum, DS. Targeted SERCA2a gene expression identifies molecular mechanism and therapeutic target for arrhythmogenic cardiac alternans. Circ Arrhythm Electrophysiol 2009, 2, 686–94. [Google Scholar] [CrossRef] [PubMed]
- Ordog, B; Brutyo, E; Puskas, LG; Papp, JG; Varro, A; Szabad, J; et al. Gene expression profiling of human cardiac potassium and sodium channels. Int J Cardiol 2006, 111, 386–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z; Yue, L; White, M; Pelletier, G; Nattel, S. Differential distribution of inward rectifier potassium channel transcripts in human atrium versus ventricle. Circulation 1998, 98, 2422–8. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, D; Xu, D; Timofeyev, V; Lu, L; Sharma, D; Zhang, Z; et al. Differential expression of small-conductance Ca2+-activated K+ channels SK1, SK2, and SK3 in mouse atrial and ventricular myocytes. Am J Physiol Heart Circ Physiol 2005, 289, H2714–23. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, CH; Chang, PC; Hsieh, YC; Reher, T; Chen, PS; Lin, SF. Proarrhythmic effect of blocking the small conductance calcium activated potassium channel in isolated canine left atrium. Heart Rhythm 2013, 10, 891–8. [Google Scholar] [CrossRef] [PubMed]
- Szigeti, G; Rusznak, Z; Kovacs, L; Papp, Z. Calcium-activated transient membrane currents are carried mainly by chloride ions in isolated atrial, ventricular and Purkinje cells of rabbit heart. Exp Physiol 1998, 83, 137–53. [Google Scholar] [CrossRef] [PubMed]
- Dobrzynski, H; Marples, DD; Musa, H; Yamanushi, TT; Henderson, Z; Takagishi, Y; et al. Distribution of the muscarinic K+ channel proteins Kir3.1 and Kir3.4 in the ventricle, atrium, and sinoatrial node of heart. J Histochem Cytochem 2001, 49, 1221–34. [Google Scholar] [CrossRef] [PubMed]
- Bingen, BO; Neshati, Z; Askar, SF; Kazbanov, IV; Ypey, DL; Panfilov, AV; et al. Atrium-specific Kir3.x determines inducibility, dynamics, and termination of fibrillation by regulating restitution-driven alternans. Circulation 2013, 128, 2732–44. [Google Scholar] [CrossRef] [PubMed]
- Ravens, U; Wettwer, E. Ultra-rapid delayed rectifier channels: molecular basis and therapeutic implications. Cardiovasc Res 2011, 89, 776–85. [Google Scholar] [CrossRef] [PubMed]
- Kanaporis, G; Blatter, LA. Calcium-activated chloride current determines action potential morphology during calcium alternans in atrial myocytes. J Physiol 2016, 594, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Kanaporis, G; Blatter, LA. Ca2+-activated chloride channel activity during Ca2+ alternans in ventricular myocytes. Channels (Austin) 2016, 10, 507–17. [Google Scholar] [CrossRef] [PubMed]
- Al Abed, A; Lovell, NH; Dokos, S. Local heterogeneous electrical restitution properties of rabbit atria. J Cardiovasc Electrophysiol 2016, 27, 743–53. [Google Scholar] [CrossRef] [PubMed]
- Hori, Y; Nakahara, S; Anjo, N; Nakagawa, A; Nishiyama, N; Yamada, K; et al. Investigation of the atrial conduction time measured by tissue Doppler imaging at the left atrial appendage and the actual electrical conduction time: consideration of left atrial remodeling in atrial fibrillation patients. J Interv Card Electrophysiol 2016. [Google Scholar] [CrossRef] [PubMed]
- Monigatti-Tenkorang, J; Jousset, F; Pascale, P; Vesin, JM; Ruchat, P; Fromer, M; et al. Intermittent atrial tachycardia promotes repolarization alternans and conduction slowing during rapid rates, and increases susceptibility to atrial fibrillation in a free-behaving sheep model. J Cardiovasc Electrophysiol 2014, 25, 418–27. [Google Scholar] [CrossRef] [PubMed]
- Greiser, M; Schotten, U. Dynamic remodeling of intracellular Ca2+ signaling during atrial fibrillation. J Mol Cell Cardiol 2013, 58, 134–42. [Google Scholar] [CrossRef] [PubMed]
- Gaborit, N; Steenman, M; Lamirault, G; Le Meur, N; Le Bouter, S; Lande, G; et al. Human atrial ion channel and transporter subunit gene-expression remodeling associated with valvular heart disease and atrial fibrillation. Circulation 2005, 112, 471–81. [Google Scholar] [CrossRef] [PubMed]
- Gemel, J; Levy, AE; Simon, AR; Bennett, KB; Ai, X; Akhter, S; et al. Connexin40 abnormalities and atrial fibrillation in the human heart. J Mol Cell Cardiol 2014, 76, 159–68. [Google Scholar] [CrossRef] [PubMed]
- Boldt, A; Wetzel, U; Lauschke, J; Weigl, J; Gummert, J; Hindricks, G; et al. Fibrosis in left atrial tissue of patients with atrial fibrillation with and without underlying mitral valve disease. Heart 2004, 90, 400–5. [Google Scholar] [CrossRef] [PubMed]
- Merchant, FM; Sayadi, O; Moazzami, K; Puppala, D; Armoundas, AA. T-wave alternans as an arrhythmic risk stratifier: state of the art. Curr Cardiol Rep 2013, 15, 398. [Google Scholar] [CrossRef] [PubMed]
- Verrier, RL; Fuller, H; Justo, F; Nearing, BD; Rajamani, S; Belardinelli, L. Unmasking atrial repolarization to assessalternans, spatiotemporal heterogeneity, and susceptibility to atrial fibrillation. Heart Rhythm 2016, 13, 953–61. [Google Scholar] [CrossRef] [PubMed]
- Narayan, SM; Wright, M; Derval, N; Jadidi, A; Forclaz, A; Nault, I; et al. Classifying fractionated electrograms in human atrial fibrillation using monophasic action potentials and activation mapping: evidence for localized drivers, rate acceleration, and nonlocal signal etiologies. Heart Rhythm 2011, 8, 244–53. [Google Scholar] [CrossRef] [PubMed]
- Lalani, GG; Schricker, AA; Clopton, P; Krummen, DE; Narayan, SM. Frequency analysis of atrial action potential alternans: a sensitive clinical index of individual propensity to atrial fibrillation. Circ Arrhythm Electrophysiol 2013, 6, 859–67. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y; Xie, F; Stein, KM; Garfinkel, A; Culianu, CA; Lerman, BB; et al. Mechanism underlying initiation of paroxysmal atrial flutter/atrial fibrillation by ectopic foci: a simulation study. Circulation 2007, 115, 2094–102. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z; Cui, B; He, B; Hu, X; Wu, W; Wu, L; et al. Distinct restitution properties in vagally mediated atrial fibrillation and six-hour rapid pacing-induced atrial fibrillation. Cardiovasc Res 2011, 89, 834–42. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, YC; Lin, JC; Hung, CY; Li, CH; Lin, SF; Yeh, HI; et al. Gap junction modifier rotigaptide decreases the susceptibility to ventricular arrhythmia by enhancing conduction velocity and suppressing discordant alternans during therapeutic hypothermia in isolated rabbit hearts. Heart Rhythm 2016, 13, 251–61. [Google Scholar] [CrossRef] [PubMed]
- Wilson, LD; Jeyaraj, D; Wan, X; Hoeker, GS; Said, TH; Gittinger, M; et al. Heart failure enhances susceptibility to arrhythmogenic cardiac alternans. Heart Rhythm 2009, 6, 251–9. [Google Scholar] [CrossRef] [PubMed]
- Gelzer, AR; Koller, ML; Otani, NF; Fox, JJ; Enyeart, MW; Hooker, GJ; et al. Dynamic mechanism for initiation of ventricular fibrillation in vivo. Circulation 2008, 118, 1123–9. [Google Scholar] [CrossRef] [PubMed]
- Wilson, LD; Rosenbaum, DS. Mechanisms of arrythmogenic cardiac alternans. Europace 2007, 9 (Suppl 6), vi77–82. [Google Scholar] [CrossRef] [PubMed]
- Katra, RP; Pruvot, E; Laurita, KR. Intracellular calcium handling heterogeneities in intact guinea pig hearts. Am J Physiol Heart Circ Physiol 2004, 286, H648–56. [Google Scholar] [CrossRef] [PubMed]
- Laurita, KR; Katra, R; Wible, B; Wan, X; Koo, MH. Transmural heterogeneity of calcium handling in canine. Circ Res 2003, 92, 668–75. [Google Scholar] [CrossRef] [PubMed]
- Sato, D; Shiferaw, Y; Garfinkel, A; Weiss, JN; Qu, Z; Karma, A. Spatially discordant alternans in cardiac tissue: role of calcium cycling. Circ Res 2006, 99, 520–7. [Google Scholar] [CrossRef] [PubMed]
- Sato, D; Bers, DM; Shiferaw, Y. Formation of spatially discordant alternans due to fluctuations and diffusion of calcium. PLoS One 2013, 8, e85365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severs, NJ; Bruce, AF; Dupont, E; Rothery, S. Remodelling of gap junctions and connexin expression in diseased myocardium. Cardiovasc Res 2008, 80, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Guerra, JM; Everett, THt; Lee, W; Wilson, E; Olgin, JE. Effects of the gap junction modifier rotigaptide (ZP123) on atrial conduction and vulnerability to atrial fibrillation. Circulation 2006, 114, 110–8. [Google Scholar] [CrossRef] [PubMed]
- van Rijen, HV; Eckardt, D; Degen, J; Theis, M; Ott, T; Willecke, K; et al. Slow conduction and enhanced anisotropy increase the propensity for ventricular tachyarrhythmias in adult mice with induced deletion of connexin43. Circulation 2004, 109, 1048–55. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M; Yokoshiki, H; Mitsuyama, H; Mizukami, K; Ono, T; Tsutsui, H. Conduction and refractory disorders in the diabetic atrium. Am J Physiol Heart Circ Physiol 2012, 303, H86–95. [Google Scholar] [CrossRef] [PubMed]






© 2017 The Lithuanian University of Health Sciences. Production and hosting by Elsevier Sp. z o.o. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
Share and Cite
Kanaporis, G.; Blatter, L.A. Alternans in atria: Mechanisms and clinical relevance. Medicina 2017, 53, 139-149. https://doi.org/10.1016/j.medici.2017.04.004
Kanaporis G, Blatter LA. Alternans in atria: Mechanisms and clinical relevance. Medicina. 2017; 53(3):139-149. https://doi.org/10.1016/j.medici.2017.04.004
Chicago/Turabian StyleKanaporis, Giedrius, and Lothar A. Blatter. 2017. "Alternans in atria: Mechanisms and clinical relevance" Medicina 53, no. 3: 139-149. https://doi.org/10.1016/j.medici.2017.04.004