Interaction of ethidium and tetraphenylphosphonium cations with Salmonella enterica cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and methods
2.1. Bacteria cultivation and preparation for experiments
2.2. Fluorescence measurements
2.3. Potentiometric measurements
3. Results
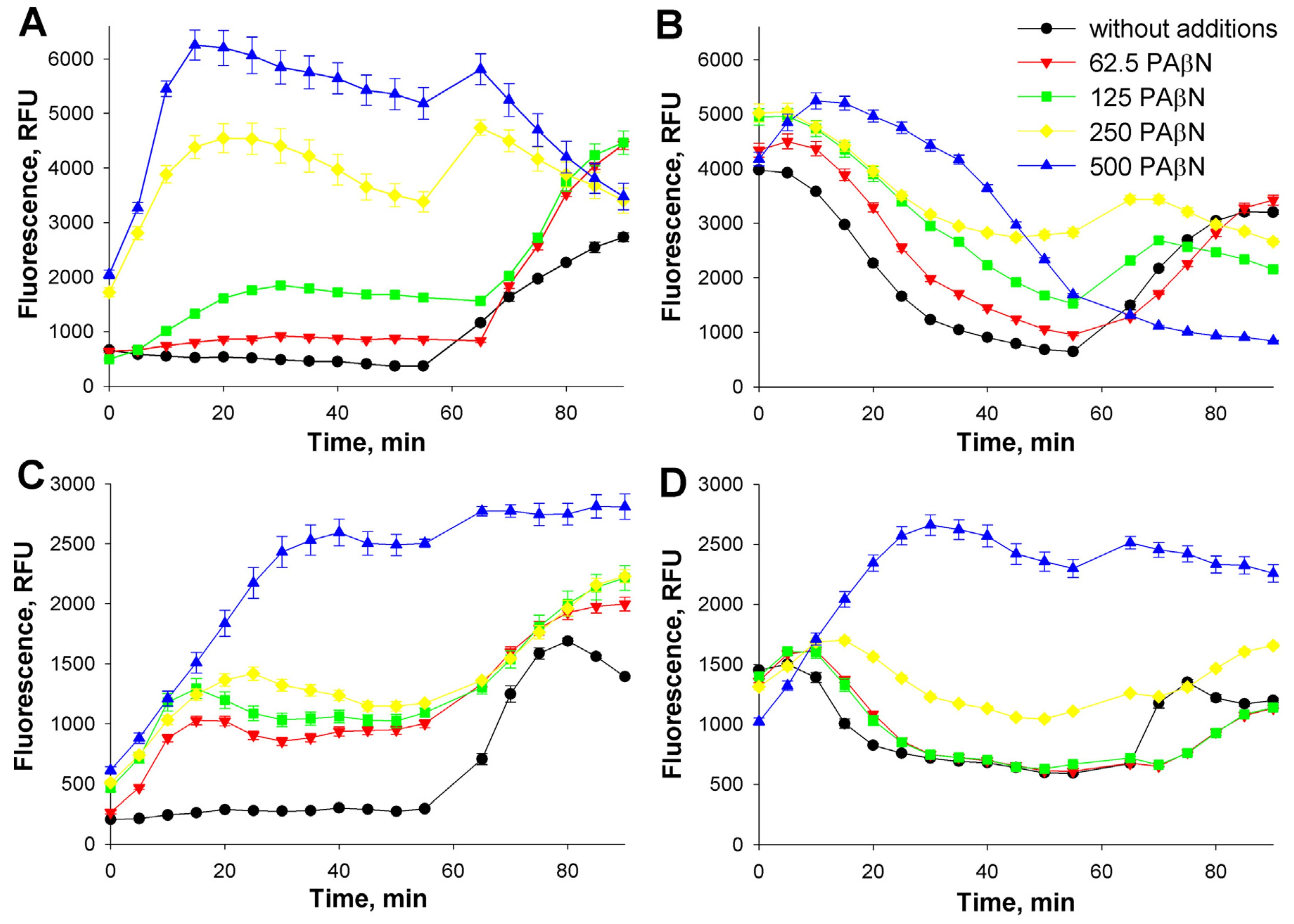
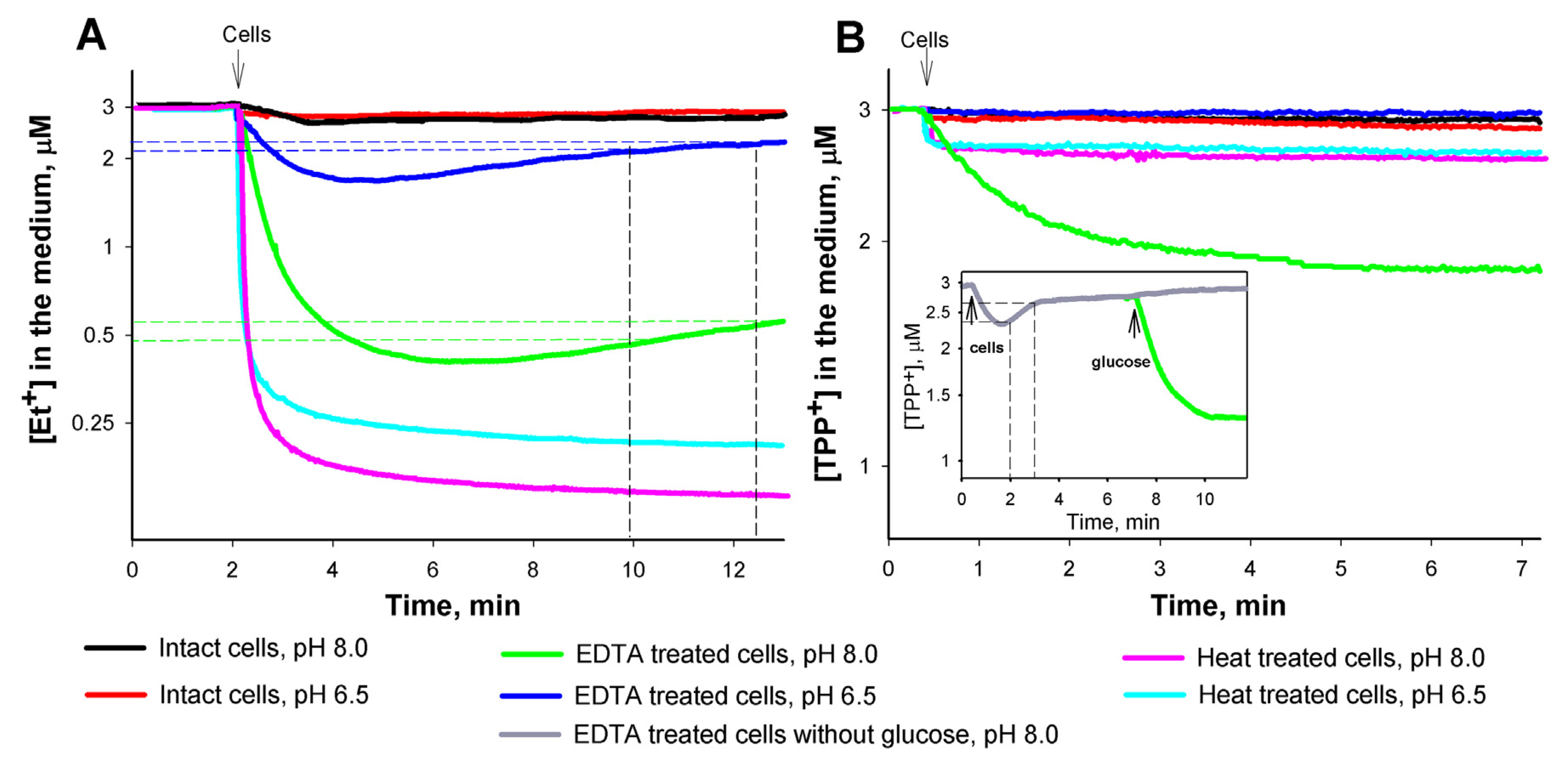
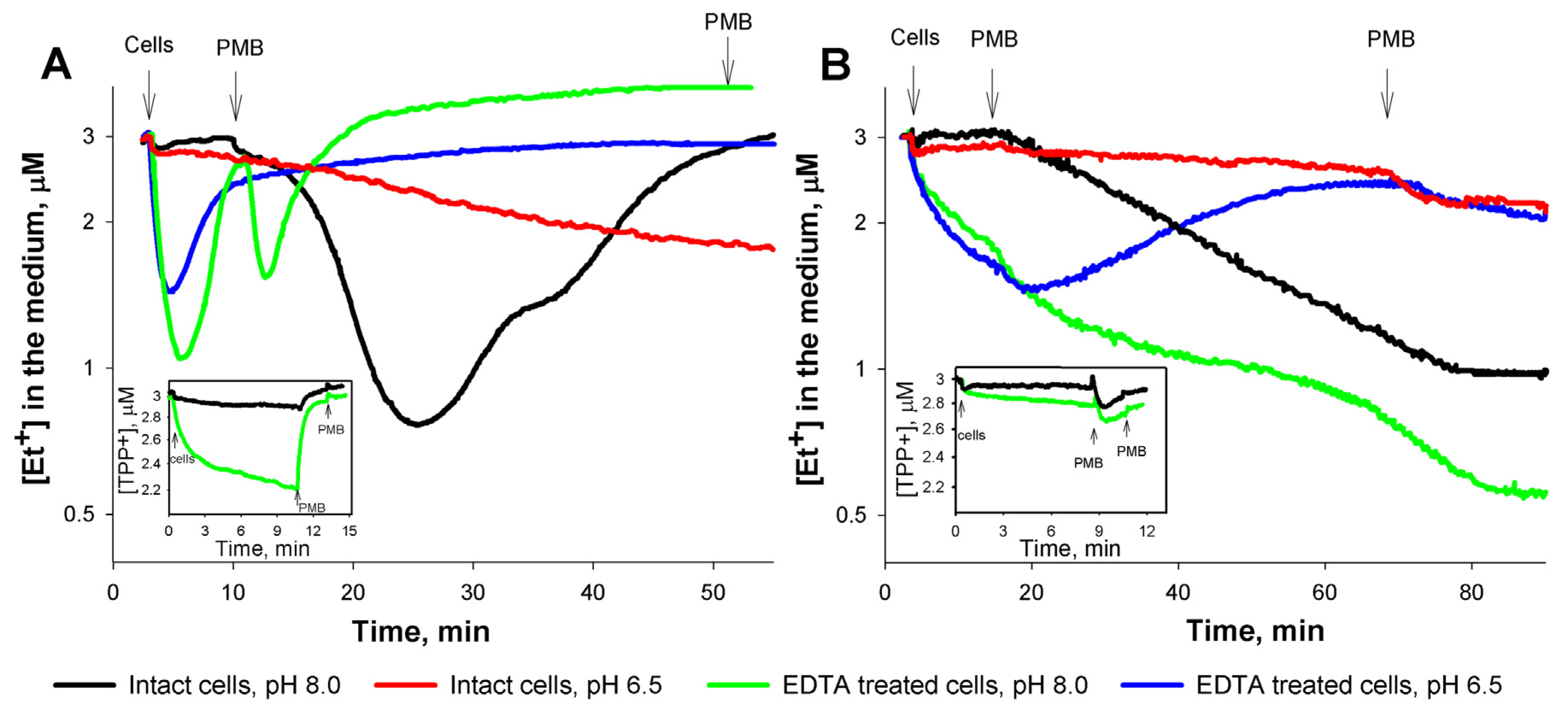
3.1. Role of the medium pH and the outer membrane (OM) permeability in the interaction of Et+ with S. enterica cells
3.2. Influence of the incubation buffer concentration on the efflux
3.3. Influence of the incubation temperature on the interaction of indicator compounds with S. enterica cells
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
Acknowledgments
References
- Nikaido, H; Pagès, JM. Broad specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol Rev 2012, 36, 340–63. [Google Scholar] [CrossRef] [PubMed]
- Li, X; Plésiat, P; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin Microbiol Rev 2015, 28, 337–418. [Google Scholar] [CrossRef] [PubMed]
- Blair, J; Smith, HE; Ricci, V; Lawler, AJ; Thompson, LJ; Piddock, LJV. Expression of homologous RND efflux pump genes is dependent upon AcrB expression: implications for efflux and virulence inhibitor design. J Antimicrob Chemother 2015, 70, 424–31. [Google Scholar] [CrossRef] [PubMed]
- Dhanani, A; Block, G; Dewar, K; Forgetta, V; Topp, E; Beiko, RG; et al. Genomic comparison of non-typhoidal Salmonella enterica serovars Typhimurium, Enteritidis, Heidelberg, Hadar and Kentucky isolates from broiler chickens. PLOS ONE 2015, 10, e0128773. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y; Huang, YW; Chen, SJ; Chang, CW; Yang, TC. The SmeYZ efflux pump of Stenotrophomonas maltophilia contributes to drug resistance, virulence-related characteristics, and virulence in mice. Antimicrob Agents Chemother 2015, 59, 4067–73. [Google Scholar] [CrossRef] [PubMed]
- Baugh, S; Phillips, CR; Ekanayaka, AS; Piddock, LJV; Webber, MA. Inhibition of multidrug efflux as a strategy to prevent biofilm formation. J Antimicrob Chemother 2014, 69, 673–81. [Google Scholar] [CrossRef] [PubMed]
- Midgley, M. The phosphonium ion efflux system of Escherichia coli: relationship to the ethidium efflux system and energetic studies. J Gen Microbiol 1986, 132, 3187–93. [Google Scholar] [CrossRef] [PubMed]
- Gros, P; Talbot, F; Tang-Wai, D; Bibi, E; Kaback, HR. Lipophilic cations: a group of model substrates for the multidrug-resistance transporter. Biochemistry 1992, 31, 1992–8. [Google Scholar] [CrossRef] [PubMed]
- Paixão, L; Rodrigues, L; Couto, I; Martins, M; Fernandes, P; de Carvalho, CC; et al. Fluorometric determination of ethidium bromide efflux kinetics in Escherichia coli. J Biol Eng 2009, 3, 18. [Google Scholar]
- Daugelavicius, R; Buivydas, A; Sencilo, A; Bamford, DH. Assessment of the activity of RND-type multidrug efflux pumps in Pseudomonas aeruginosa using tetraphenylphosphonium ions. Int J Antimicrob Agents 2010, 36, 234–8. [Google Scholar] [CrossRef] [PubMed]
- Ye, C; Wang, Z; Lu, W; Zhong, M; Chai, Q; Wei, Y. Correlation between AcrB trimer association affinity and efflux activity. Biochemistry 2014, 53, 3738–46. [Google Scholar] [CrossRef] [PubMed]
- Severini, A; Morgan, AR. An assay for proteinases and their inhibitors based on DNA/ethidium bromide fluorescence. Anal Biochem 1991, 193, 83–9. [Google Scholar] [CrossRef]
- Hockings, P; Rogers, PJ. The measurement of transmembrane electrical potential with lipophilic cations. Biochim Biophys Acta 1996, 1282, 101–6. [Google Scholar] [CrossRef]
- Babayan, A; Nikaido, H. In Pseudomonas aeruginosa ethidium bromide does not induce its own degradation or the assembly of pumps involved in its efflux. Biochem Biophys Res Commun 2004, 324, 1065–8. [Google Scholar] [CrossRef] [PubMed]
- Martins, A; Spengler, G; Martins, M; Rodrigues, L; Viveiros, M; Davin-Regli, A; et al. Physiological characterisation of the efflux pump system of antibiotic-susceptible and multidrug-resistant Enterobacter aerogenes. Int J Antimicrob Agents 2010, 36, 313–8. [Google Scholar] [CrossRef] [PubMed]
- Baylay, A; Piddock, LJ. Clinically relevant fluoroquinolone resistance due to constitutive overexpression of the PatAB ABC transporter in Streptococcus pneumoniae is conferred by disruption of a transcriptional attenuator. J Antimicrob Chemother 2015, 70, 670–9. [Google Scholar] [CrossRef] [PubMed]
- Blair, J; Piddock, LJ. How to measure export via bacterial multidrug resistance efflux pumps. MBio 2016. pii:e00840-00816. [Google Scholar] [CrossRef] [PubMed]
- Cerca, P; Martins, A; Cuoto, I; Viveiros, M; Amaral, L. Competition between substrates of the efflux pump system of Salmonella enteritidis. In Vivo 2011, 25, 597–602. [Google Scholar] [PubMed]
- Sutkuvienė, S; Mikalayeva, V; Pavan, S; Berti, F; Daugelavicius, R. Evaluation of the efficiency of synthesized efflux pump inhibitors on Salmonella enterica ser. typhimurium cells. Chem Biol Drug Des 2013, 82, 438–45. [Google Scholar]
- Cammann, K. Working with ion-selective electrodes; Springer-Verlag: Berlin, 1979. [Google Scholar]
- Li, W; Nordenskiold, L; Mu, Y. Sequence-specific Mg2+–DNA interactions: a molecular dynamics simulation study. J Phys Chem B 2011, 115, 14713–20. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M; Harada, Y. Direct observation of the reversible unwinding of a single DNA molecule caused by the intercalation of ethidium bromide. Nucleic Acids Res 2007, 35, e125. [Google Scholar]
- Nicholls, D; Ferguson, SJ. Bioenergetics, 4th ed.; Academic Press: San Diego, 2013. [Google Scholar]
- Martins, A; Spengler, G; Rodrigues, L; Viveiros, M; Ramos, J; Martins, M; et al. pH modulation of efflux pump activity of multi-drug resistant Escherichia coli: protection during its passage and eventual colonization of the colon. PLoS One 2009, 4, e6656. [Google Scholar]
- Amaral, L; Fanning, S; Pagès, JM. Efflux pumps of gram-negative bacteria: genetic responses to stress and the modulation of their activity by pH, inhibitors, and phenothiazines. Adv Enzymol Relat Areas Mol Biol 2011, 61–108. [Google Scholar]
- Bohnert, J; Karamian, B; Nikaido, H. Optimized Nile Red efflux assay of AcrAB-TolC multidrug efflux system shows competition between substrates. Antimicrob Agents Chemother 2010, 54, 3770–5. [Google Scholar]
- Iyer, R; Erwin, AL. Direct measurement of efflux in Pseudomonas aeruginosa using an environment-sensitive fluorescent dye. Res Microbiol 2015, 166, 516–24. [Google Scholar]
- Daugelavičius, R; Bakiene, E; Bamford, DH. Stages of polymyxin B interaction with the Escherichia coli cell envelope. Antimicrob Agents Chemother 2000, 44, 2969–78. [Google Scholar]
- Masuda, N; Sakagawa, E; Ohya, S; Gotoh, N; Tsujimoto, H; Nishino, T. Substrate specificities of MexAB-OprM, MexCD-OprJ, and MexXY-oprM efflux pumps in Pseudomonas aeruginosa. Antimicrob Agents Chemother 2000, 44, 3322–7. [Google Scholar]
- Spengler, G; Rodrigues, L; Martins, A; Martins, M; McCusker, M; Cerca, P; et al. Genetic response of Salmonella enterica serotype Enteritidis to thioridazine rendering the organism resistant to the agent. Int J Antimicrob Agents 2012, 39, 16–21. [Google Scholar]
- Kong, L; Liu, Z; Hu, X; Liu, S. Interaction of polymyxin B with ds-DNA, and determination of DNA or polymyxin B via resonance Rayleigh scattering and resonance non-linear scattering spectra. Microchim Acta 2011, 173, 207–13. [Google Scholar]
- de Mendoza, D. Temperature sensing by membranes. Annu Rev Microbiol 2014, 68, 101–16. [Google Scholar] [CrossRef] [PubMed]
- Katsu, T; Yoshimura, S; Tsuchiya, T; Fujita, Y. Temperature dependence of action of polymyxin B on Escherichia coli. J Biochem 1984, 95, 1645–53. [Google Scholar]
- Bailey, A; Paulsen, IT; Piddock, LJ. RamA confers multidrug resistance in Salmonella enterica via increased expression of acrB, which is inhibited by chlorpromazine. Antimicrob Agents Chemother 2008, 52, 3604–11. [Google Scholar]
- Lomovskaya, O; Warren, MS; Lee, A; Galazzo, J; Fronko, R; Lee, M; et al. Identification and characterization of inhibitors of multidrug resistance efflux pumps in Pseudomonas aeruginosa: novel agents for combination therapy. Antimicrob Agents Chemother 2001, 45, 105–16. [Google Scholar]
- Iino, R; Nishino, K; Noji, H; Yamaguchi, A; Matsumoto, Y. A microfluidic device for simple and rapid evaluation of multidrug efflux pump inhibitors. Front Microbiol 2010, 3, 40. [Google Scholar]
- Kinana, A; Vargiu, AV; May, T; Nikaido, H. Aminoacyl b-naphthylamides as substrates and modulators of AcrB multidrug efflux pump. Proc Natl Acad Sci U S A 2016, 113, 1405–10. [Google Scholar]
- Lamers, R; Cavallari, JF; Burrows, LL. The efflux inhibitor phenylalanine–arginine beta naphthylamide (PAβN) permeabilizes the outer membrane of gram-negative bacteria. PLOS ONE 2013, 8, e60666. [Google Scholar]
- Misra, R; Morrison, KD; Cho, HJ; Khuu, T. Importance of realtime assays to distinguish multidrug efflux pumpinhibiting and outer membrane-destabilizing activities in Escherichia coli. J Bacteriol 2015, 197, 2479–88. [Google Scholar]
- Hunter, R; Phoenix, VR; Saxena, A; Beveridge, TJ. Impact of growth environment and physiological state on metal immobilization by Pseudomonas aeruginosa PAO1. Can J Microbiol 2010, 56, 527–38. [Google Scholar]





© 2017 The Lithuanian University of Health Sciences. Production and hosting by Elsevier Sp. z o.o. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
Share and Cite
Mikalayeva, V.; Sakalauskaitė, S.; Daugelavičius, R. Interaction of ethidium and tetraphenylphosphonium cations with Salmonella enterica cells. Medicina 2017, 53, 122-130. https://doi.org/10.1016/j.medici.2017.04.001
Mikalayeva V, Sakalauskaitė S, Daugelavičius R. Interaction of ethidium and tetraphenylphosphonium cations with Salmonella enterica cells. Medicina. 2017; 53(2):122-130. https://doi.org/10.1016/j.medici.2017.04.001
Chicago/Turabian StyleMikalayeva, Valeryia, Sandra Sakalauskaitė, and Rimantas Daugelavičius. 2017. "Interaction of ethidium and tetraphenylphosphonium cations with Salmonella enterica cells" Medicina 53, no. 2: 122-130. https://doi.org/10.1016/j.medici.2017.04.001