
Production of Recombinant Single-Chain Eel Luteinizing Hormone and Follicle-Stimulating Hormone Analogs in Chinese Hamster Ovary Suspension Cell Culture

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
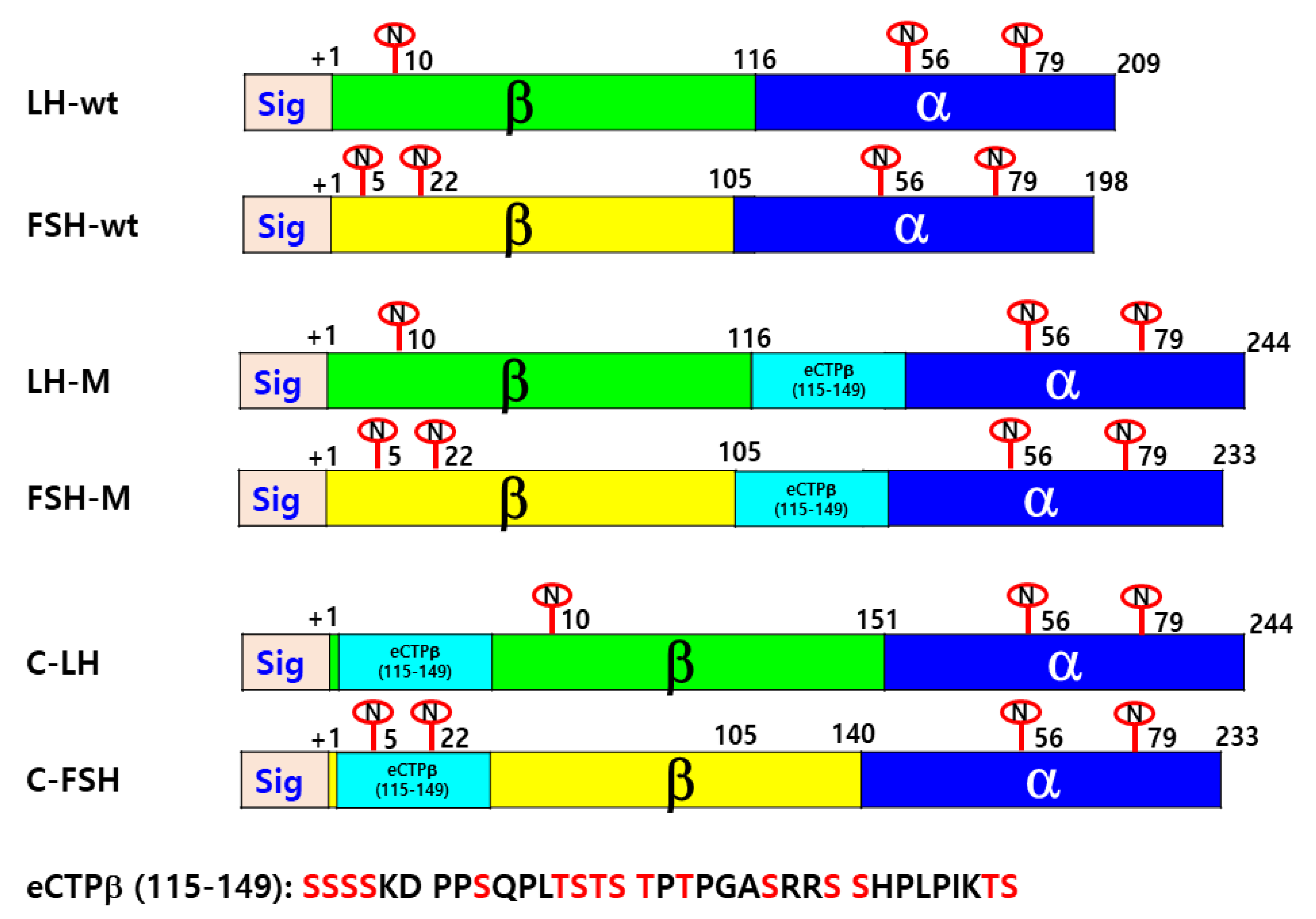
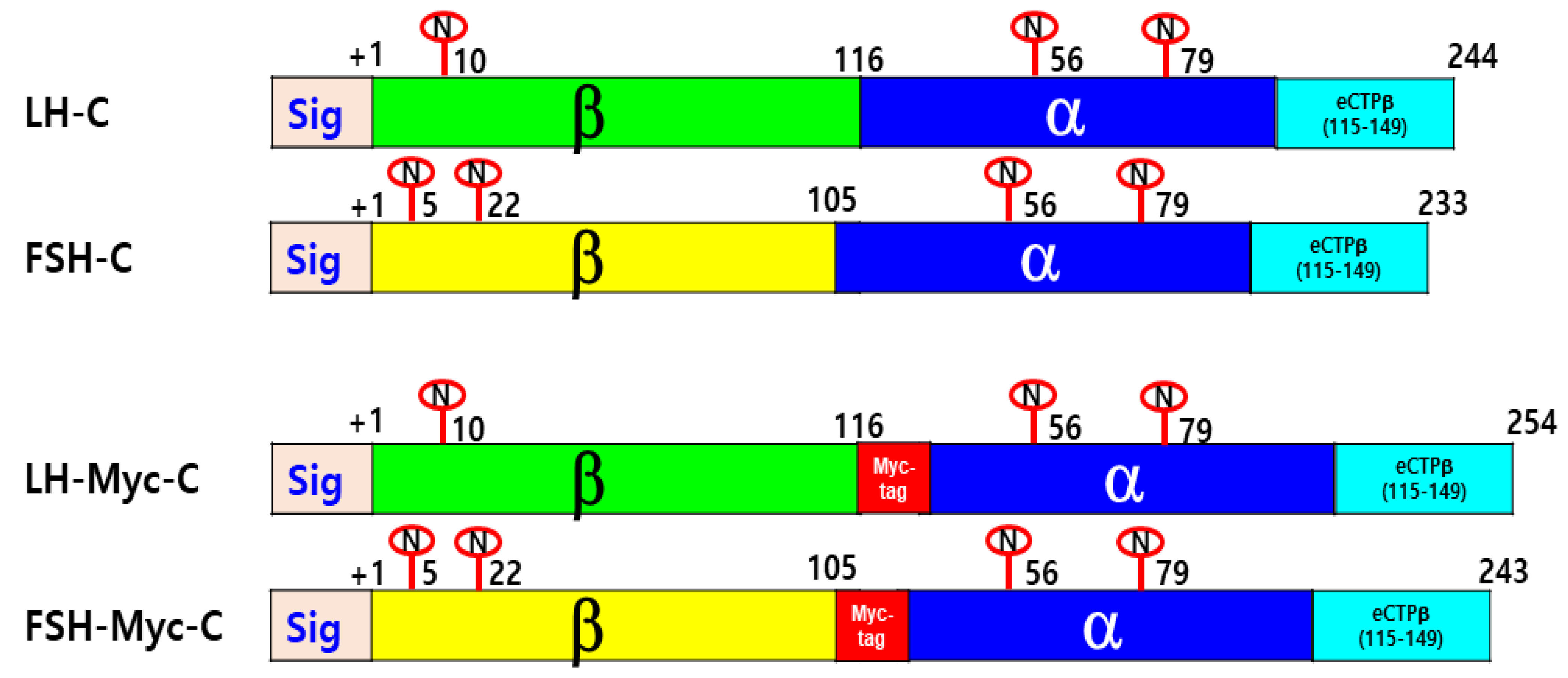
2.2. Construction of Vectors for the Expression of Eel LH-wt, FSH-wt, and Mutant Proteins
2.3. Expression of Rec-Eel LH-wt, Rec-Eel FSH, and Mutant Proteins
2.4. Quantitation of Rec-Eel LH and Eel FSH Proteins Using ELISA
2.5. Western Blotting Analysis of Rec-Proteins
2.6. Enzymatic Release of N-Linked Oligosaccharides
2.7. Analysis of cAMP Levels Using Homogenous Time-Resolved Fluorescence Assay
2.8. Data Analysis
3. Results
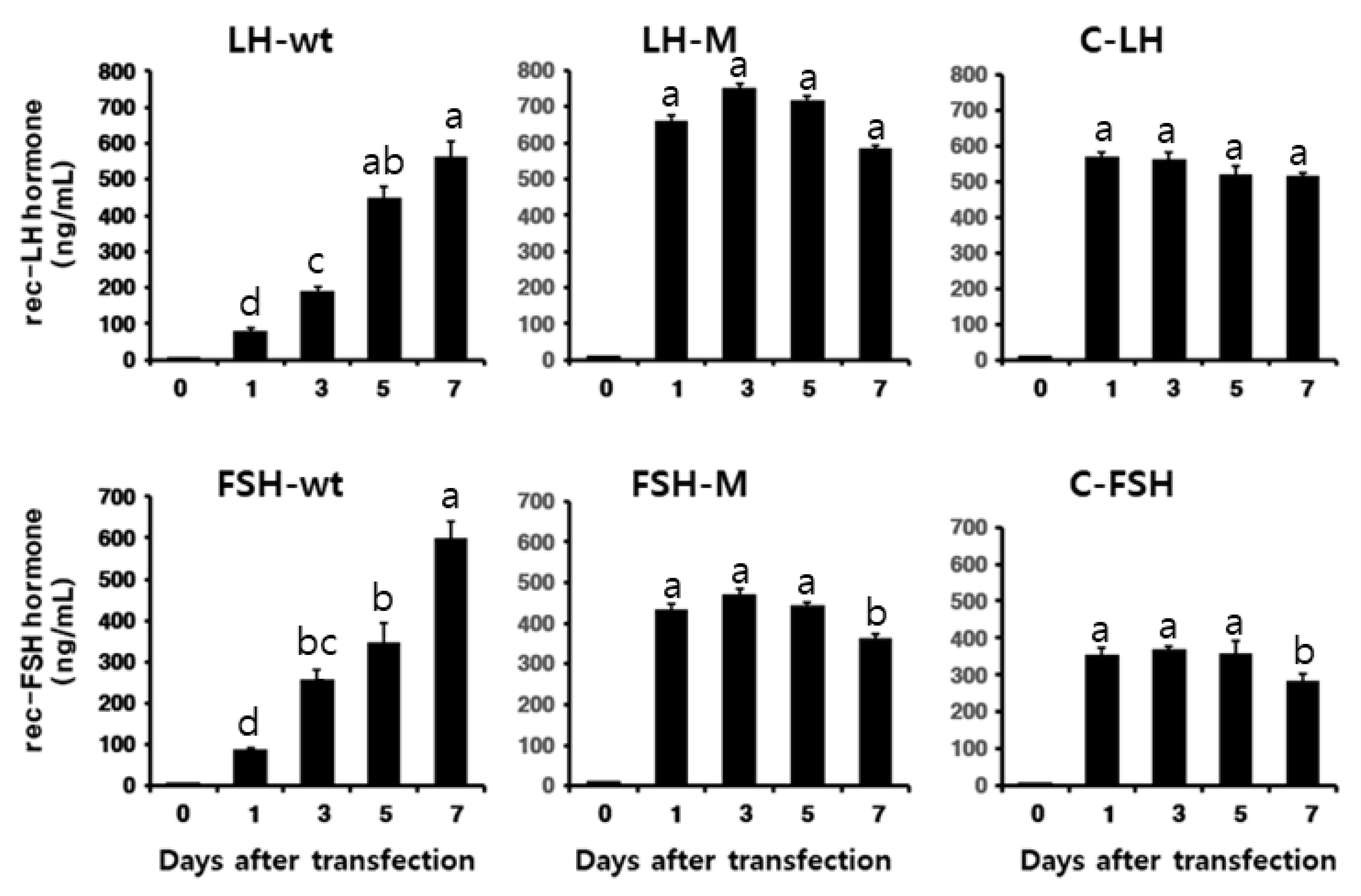
3.1. Secretion of Rec-Eel LHβ/α-wt, Eel FSHβ/α-wt, and Mutant Proteins
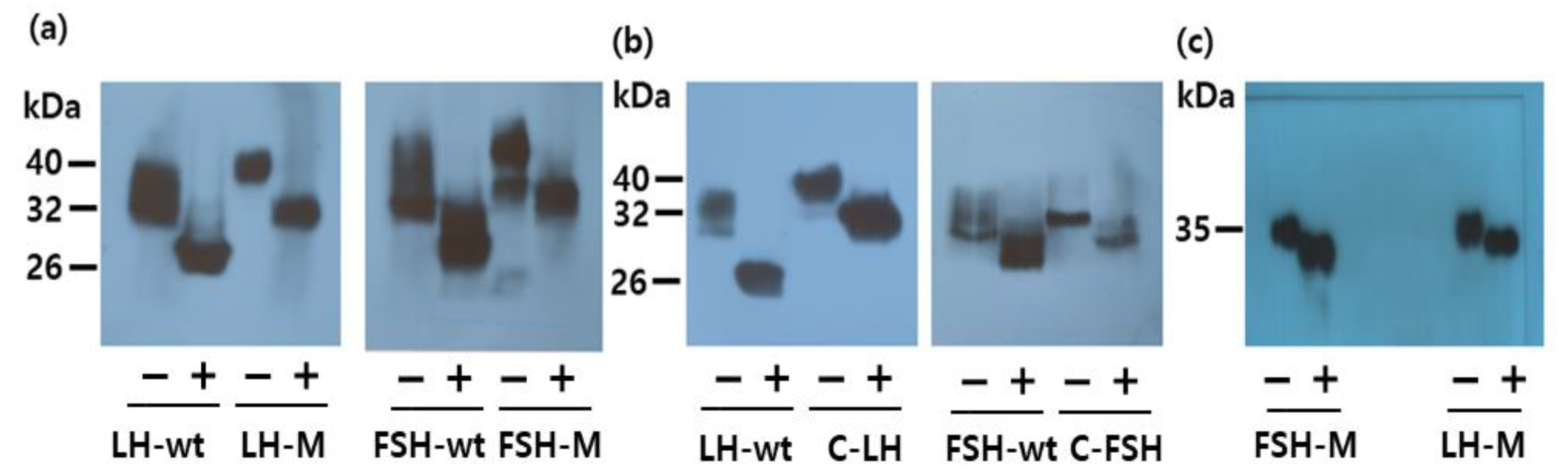
3.2. Deglycosylation of the Rec-Eel LH and FSH Proteins
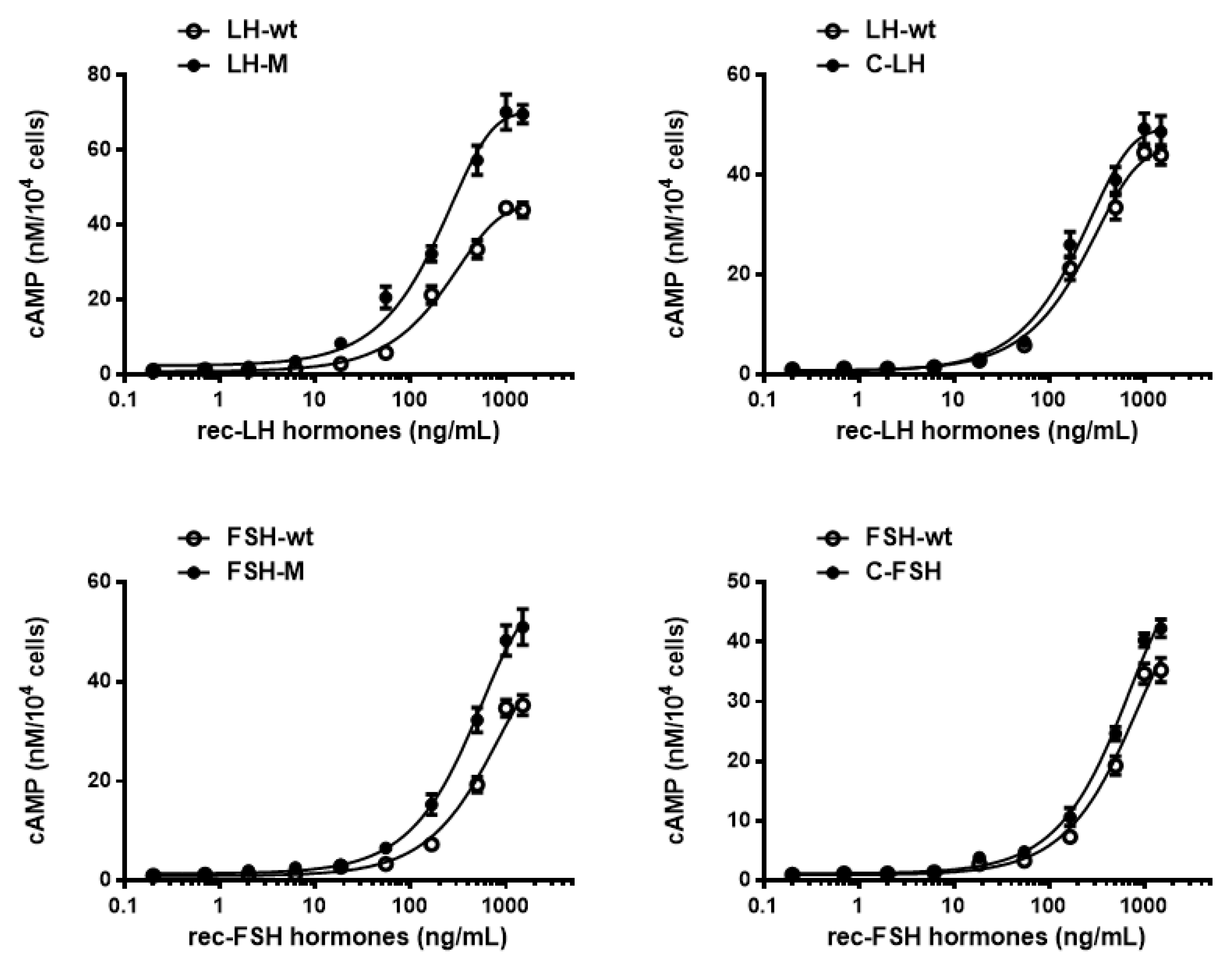
3.3. Biological Activities of the Variants
3.4. Characterization of the Other Mutants with eCTP and Myc-tag
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byambaragchaa, M.; Park, A.; Gil, S.J.; Lee, H.W.; Ko, Y.J.; Choi, S.H.; Kang, M.H.; Min, K.S. Luteinizing hormone-like and follicle-stimulating hormone-like activities of equine chorionic gonadotropin β-subunit mutants in cells expressing rat luteinizing hormone/chorionic gonadotropin receptor and rat follicle-stimulating hormone receptor. Anim. Cells Syst. 2021, 25, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Byambaragchaa, M.; Kang, H.J.; Choi, S.H.; Kang, M.H.; Min, K.S. Specific roles of N- and O-linked oligosaccharide sites on biological activity of equine chorionic gonadotropin (eCG) in cells expressing rat luteinizing hormone/chorionic gonadotropin receptor (LH/CGR) and follicle-stimulating hormone receptor (FSHR). BMC Biotechnol. 2021, 21, 52. [Google Scholar] [CrossRef] [PubMed]
- Min, K.S.; Park, J.J.; Lee, S.Y.; Byambragchaa, M.; Kang, M.H. Comparative gene expression profiling of mouse ovaries upon stimulation with natural equine chorionic gonadotropin (N-eCG) and tethered recombinant-eCG (R-eCG). BMC Biotechnol. 2020, 20, 59. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X. Inactivation mutations of G protein-couped receptors and disease: Structure-function insights and therapeutic implications. Pharmacol. Ther. 2006, 111, 949–973. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X.; Abell, A.N.; Liu, X.; Nakamura, K.; Segaloff, D.L. Constitutive activation of G protein-coupled receptors as a result of selective substitution of a conserved leucine residue in transmembrane helix III. Mol. Endocrinol. 2000, 14, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ji, R.L.; Tao, Y.X. Naturally occurring mutations in G protein-coupled receptors associated with obesity and type2 diabetes mellitus. Pharmacol. Ther. 2022, 234, 108044. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Byambaragchaa, M.; Kim, D.J.; Lee, J.H.; Kang, M.H.; Min, K.S. Specific signal transduction of constitutively activating (D576G) and inactivating (R476H) mutants of agonist-stimulated luteinizing hormone receptor in eel. Int. J. Mol. Sci. 2023, 24, 9133. [Google Scholar] [CrossRef]
- Kara, E.; Crepieux, P.; Gauthier, C.; Martinat, N.; Piketty, V.; Guillou, F.; Reiter, E. A phosphorylation cluster of five serine and threonine residues in the C-terminus of the follicle-stimulating hormone receptor is important for desensitization but not for β-arrestin-mediated ERK activation. Mol. Endocrinol. 2006, 20, 3014–3026. [Google Scholar] [CrossRef]
- Piketty, V.; Kara, E.; Guillou, F.; Reiter, E.; Crepiux, P. Follicle-stimulating hormone (FSH) activates extracellular signal-regulated kinase phosphorylation independently of beta-arrestin- and dynamin-mediated FSH receptor internalization. RBE 2006, 4, 33. [Google Scholar] [CrossRef]
- Shenoy, S.K.; Barak, L.S.; Xiao, K.; Ahn, S.; Berthouze, M.; Shukla, A.K.; Luttrell, L.M.; Lefkowitz, R.J. Ubiquitination of β-arrestin links seven-transmembrane receptor endocytosis and ERK activation. J. Biol. Chem. 2007, 282, 29549–29562. [Google Scholar] [CrossRef]
- Shenoy, S.K.; Drake, M.T.; Nelson, C.D.; Houtz, D.A.; Xiao, K.; Madabushi, S.; Reiter, E.; Premont, R.T.; Lichtarge, O.; Lefkowitz, R.J. β-arrestin-arrestin-dependent, G protein-independent ERK1/2 activation by the β2 adrenergic receptor. J. Biol. Chem. 2006, 281, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Slosky, L.M.; Bai, Y.; Toth, K.; Ray, C.; Rochelle, L.K.; Badea, A.; Chandrasekhar, R.; Pogorelov, V.M.; Abraham, D.M.; Atluri, N.; et al. β-arrestin-biased allosteric modulated of NTSR1 selectively attenuates addictive behaviors. Cell 2020, 181, 1364–1379. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, M. Potential Leydig cell mitogenic signals generated by the wild-type and constitutively active mutants of the lutropin/choriogonadotropin receptor (LHR). Mol. Cell Endocrinol. 2007, 260, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, K.; Ascoli, M. Lutropin/choriogonadotropin stimulate the proliferation of primary cultures of rat Leydig cells through a pathway that involves activation of the extracellular regulated kinase 1/2 cascade. Endocrinology 2007, 148, 3214–3225. [Google Scholar] [CrossRef] [PubMed]
- Bielinska, M.; Matzuk, M.M.; Boime, I. Site-specific processing of the N-linked oligosaccharides of the human chorionic gonadotropin α subunit. J. Biol. Chem. 1989, 264, 17113–17118. [Google Scholar] [CrossRef] [PubMed]
- Bishop, L.A.; Robertson, D.M.; Cahir, N.; Schofield, P.R. Specific roles for the asparagine-linked carbohydrate residues of recombinant human follicle stimulating hormone in receptor binding and signal transduction. Mol. Endocrinol. 1994, 8, 722–731. [Google Scholar]
- Fares, F.A.; Gruener, N.; Kraiem, Z. The role of the asparagine-linked oligosaccharides of the α-subunit in human thyrotropin bioactivity. Endocrinology 1996, 137, 555–560. [Google Scholar] [CrossRef]
- Flack, M.R.; Froehlich, J.; Bennet, A.P.; Anasti, J.; Nisula, B.C. Site-directed mutagenesis defines the individual roles of the glycosylation sites on follicle-stimulating hormone. J. Biol. Chem. 1994, 269, 14015–14020. [Google Scholar] [CrossRef]
- Matzuk, M.M.; Keene, J.L.; Boime, I. Site specificity of the chorionic gonadotropin N-linked oligosaccharides in signal transduction. J. Biol. Chem. 1989, 264, 2409–2414. [Google Scholar] [CrossRef]
- Valove, F.M.; Finch, C.; Anasti, J.N.; Froehlich, J.; Flack, M.R. Receptor binding and signal transduction are dissociable functions requires different sites on follicle-stimulating hormone. Endocrinology 1994, 135, 2657–2661. [Google Scholar] [CrossRef]
- Hong, S.M.; Choi, J.H.; Jo, S.J.; Min, K.S.; Kim, D.J.; Lee, J.M.; Kusakabe, T. Heterologous production and glycosylation of Japanese eel follitropin using silkworm. Biotechnol. Bioprocess Eng. 2019, 24, 745–753. [Google Scholar] [CrossRef]
- Kamei, H.; Kaneko, T.; Aida, K. Steroidogenic activities of follicle-stimulating hormone in the ovary of Japanese eel, Anguilla japonica. Gen. Comp. Endocrinol. 2006, 146, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Kazeto, Y.; Kohara, M.; Miura, T.; Miura, C.; Yamaguchi, S.; Trant, J.M.; Adachi, S.; Yamauchi, K. Japanese eel follicle-stimulating hormone (fsh) and luteinizing hormone (lh) by Drosophila S2 cells and their differential actions on the reproductive biology. Biol. Reprod. 2008, 79, 938–946. [Google Scholar] [CrossRef]
- Kobayashi, M.; Hayakawa, Y.; Park, W.; Banba, A.; Yoshizaki, G.; Kumamaru, K.; Kagawa, H.; Nagaya, H.; Sohn, Y.C. Production of recombinant Japanese eel gonadotropins by baculovirus in silkworm larvae. Gen. Comp. Endocrinol. 2010, 167, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Fares, F. The role of O-linked and N-linked oligosaccharides on the structure-function of glycoprotein hormones: Development of agonists and antagonists. Biochim. Biophys. Acta 2006, 1760, 560–567. [Google Scholar] [CrossRef]
- Ben-Menahem, D.; Jablonka-Shariff, A.; Hyde, R.K.; Pixley, M.R.; Srivastava, S.; Berger, P.; Boime, I. The position of the alpha and beta subunits in a single chain variant of human chorionic gonadotropin affects the heterodimeric interaction of the subunits and receptor-binding epitopes. J. Biol. Chem. 2001, 276, 29871–29879. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Park, C.W.; Kim, D.W.; Park, H.K.; Byambaragchaa, M.; Lee, N.S.; Hong, S.M.; Seo, M.Y.; Kang, M.H.; Min, K.S. Production and characterization of monoclonal antibodies against recombinant tethered follicle-stimulating hormone from Japanese eel Anguilla japonica. Gen. Comp. Endocrinol. 2016, 233, 8–15. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structure proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Sugino, H.; Bousfield, G.R.; Moore, W.T.J.; Ward, D.N. Structural studies on equine glycoprotein hormones: Amino acid sequence of equine chorionic gonadotropin β-subunit. J. Biol. Chem. 1987, 262, 8603–8609. [Google Scholar] [CrossRef]
- Bousfield, G.R.; Butney, V.Y. Identification of twelve O-glycosylation sites in equine chorionic gonadotropin β and equine luteinizing hormone β by solid-phase Edman degradation. Biol. Reprod. 2001, 64, 136–147. [Google Scholar] [CrossRef]
- Sherman, G.B.; Wolfe, M.W.; Farmerie, T.A.; Clay, C.M.; Threadgill, D.S.; Sharp, D.C.; Nilson, J.H. A single gene encodes the beta-subunit of equine luteinizing hormone and chorionic gonadotropin. Mol. Endocrinol. 1992, 6, 951–959. [Google Scholar] [PubMed]
- Garcia-Campayo, V.; Sato, A.; Hirsch, B.; Sugahara, T.; Muyan, M.; Hsueh, A.J.; Boime, I. Design of stable biological active recombinant lutropin analogos. Nat. Biotechnol. 1997, 15, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Fares, F.A.; Suganuma, N.; Nishimori, K.; LaPolt, P.S.; Hsueh, A.J.W.; Boime, I. Design of a long-acting follitropin agonist by fusing the C-terminal sequence of the chorionic gonadotropin β subunit to the follitropin β subunit. Proc. Natl. Acad. Sci. USA 1992, 89, 4304–4308. [Google Scholar] [CrossRef] [PubMed]
- Kamei, H.; Kawazoe, I.; Kanekko, T.; Aida, K. Purification of follicle-stimulating hormone from immature Japanese eel Anguilla japonica, and its biochemical properties and steroidogenic activities. Gen. Comp. Endocrinol. 2005, 143, 256–257. [Google Scholar] [CrossRef] [PubMed]
- Kamei, H.; Ohira, T.; Yoshiura, Y.; Uchida, N.; Nagasawa, H.; Aida, K. Expression of biologically active recombinant follicle stimulating hormone of Japanese eel Anguilla japonica using methylotropic yeast. Gen. Comp. Endocrinol. 2003, 134, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Fares, F.A.; Yamabe, S.; Ben-Menahem, D.; Pixley, M.; Hsueh, A.J.W.; Boime, I. Conversion of thyrotropin heterodimer to a biological active single-chain. Endocrinology 1998, 139, 2459–2464. [Google Scholar] [CrossRef]
- Bhaskaran, R.S.; Ascoli, M. The post-endocytotic fate of the gonadotropin receptors is an important determinant of the desensitization of gonadotropin responses. J. Mol. Endocrinol. 2005, 34, 447–457. [Google Scholar] [CrossRef]
- Sugahara, T.; Grootenhuis, P.D.J.; Sato, A.; Kudo, M.; Ben-Menahem, D.; Pixley, M.R.; Hsueh, A.J.W.; Boime, I. Expression of biological active fusion genes encoding the common α subunit and either the CGβ or FSHβ subunits: Role of a linker sequence. Mol. Cell Endocrinol. 1996, 125, 71–77. [Google Scholar] [CrossRef]
- Sugahara, T.; Sato, A.; Kudo, M.; Ben-Menahem, D.; Pixley, M.R.; Hsueh, A.J.W.; Boime, I. Expression of biological active fusion genes encoding the common α subunit and the follicle-stimulating hormone β subunits. J. Biol. Chem. 1996, 271, 10445–10448. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rec-LH Hormones | cAMP Response | ||
|---|---|---|---|
| Basal | Log (EC50) | Rmax | |
| (nmoles/104 Cells) | (ng/mL) | (nmoles/104 Cells) | |
| LH-wt | 0.8 ± 0.4 | 222.2 | 44.9 ± 0.9 |
| (194.5–259.1) | |||
| LH-M | 1.2 ± 0.7 | 184.7 | 70.1 ± 0.7 |
| (160.5–217.5) | |||
| C-LH | 0.6 ± 0.5 | 274.5 | 49.2 ± 1.1 |
| (163.7–227.1) | |||
| rec-FSH Hormones | cAMP Response | ||
|---|---|---|---|
| Basal | Log (EC50) | Rmax | |
| (nmoles/104 Cells) | (ng/mL) | (nmoles/104 Cells) | |
| FSH-wt | 0.7 ± 0.2 | 730.2 | 52.9 ± 1.8 |
| (512.6–1269) | |||
| FSH-M | 1.2 ± 0.3 | 425.1 | 59.6 ± 1.7 |
| (371.1–497.5) | |||
| C-FSH | 0.9 ± 0.5 | 567.6 | 49.9 ± 0.6 |
| (403.6–956.2) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byambaragchaa, M.; Kim, S.-G.; Park, S.H.; Shin, M.G.; Kim, S.-K.; Kang, M.-H.; Min, K.-S. Production of Recombinant Single-Chain Eel Luteinizing Hormone and Follicle-Stimulating Hormone Analogs in Chinese Hamster Ovary Suspension Cell Culture. Curr. Issues Mol. Biol. 2024, 46, 542-556. https://doi.org/10.3390/cimb46010035
Byambaragchaa M, Kim S-G, Park SH, Shin MG, Kim S-K, Kang M-H, Min K-S. Production of Recombinant Single-Chain Eel Luteinizing Hormone and Follicle-Stimulating Hormone Analogs in Chinese Hamster Ovary Suspension Cell Culture. Current Issues in Molecular Biology. 2024; 46(1):542-556. https://doi.org/10.3390/cimb46010035
Chicago/Turabian StyleByambaragchaa, Munkhzaya, Sang-Gwon Kim, Sei Hyun Park, Min Gyu Shin, Shin-Kwon Kim, Myung-Hwa Kang, and Kwan-Sik Min. 2024. "Production of Recombinant Single-Chain Eel Luteinizing Hormone and Follicle-Stimulating Hormone Analogs in Chinese Hamster Ovary Suspension Cell Culture" Current Issues in Molecular Biology 46, no. 1: 542-556. https://doi.org/10.3390/cimb46010035