
Single-Cell RNA Sequencing Reveals Alterations in Patient Immune Cells with Pulmonary Long COVID-19 Complications

 , , and
, , and 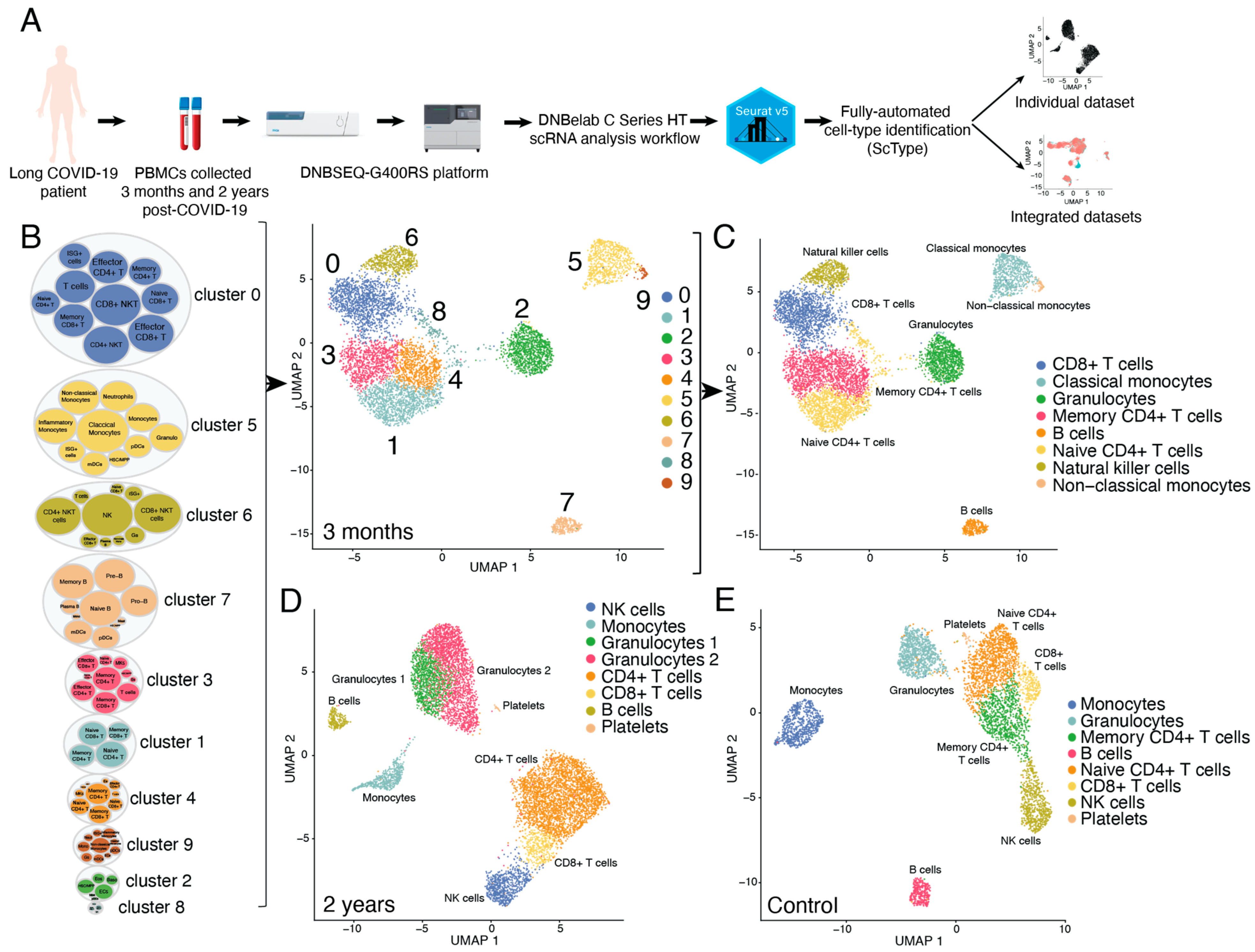{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. RNA Library Preparation
2.3. Sequencing
2.4. Preliminary Data Analysis
2.5. Single-Cell RNA Sequencing Data Analysis Using Seurat v5 R Package
2.6. Cell Type Identification Using ScType R Package
3. Results
3.1. Comprehensive Single-Sell Transcriptomic Analysis and Automated Sell-Type Annotation in Long COVID-19 PBMCs
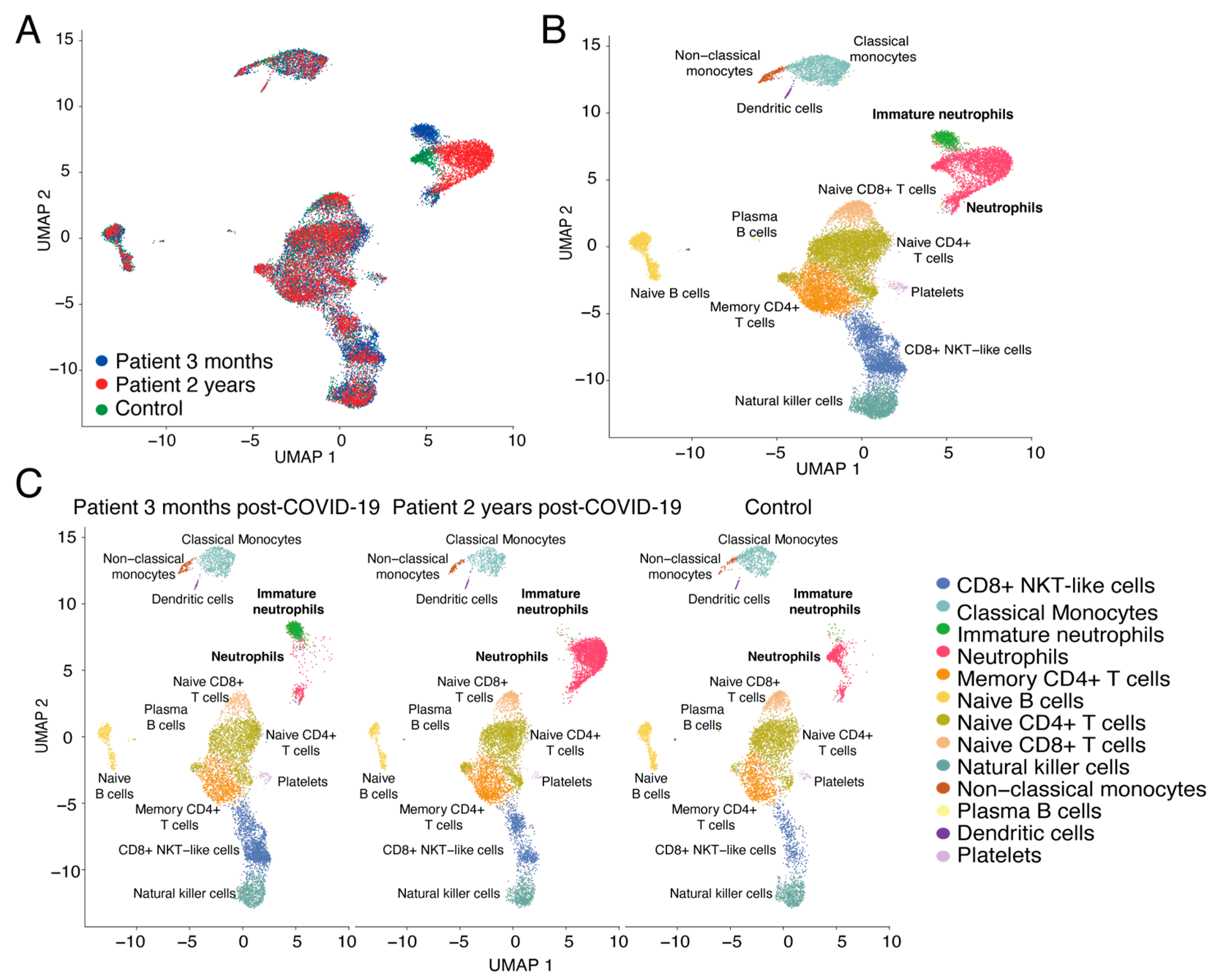
3.2. Dynamics of Neutrophil Populations and CD8+ NKT-Like Cells in Long COVID-19 Patient vs. Control
4. Discussion
Limitations of Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. More than 50 Long-Term Effects of COVID-19: A Systematic Review and Meta-Analysis. Sci. Rep. 2021, 11, 16144. [Google Scholar] [CrossRef] [PubMed]
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-Acute COVID-19 Syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Kang, L.; Guo, L.; Liu, M.; Zhou, X.; et al. 6-Month Consequences of COVID-19 in Patients Discharged from Hospital: A Cohort Study. Lancet 2021, 397, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Al-Aly, Z.; Xie, Y.; Bowe, B. High-Dimensional Characterization of Post-Acute Sequelae of COVID-19. Nature 2021, 594, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Daßler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S.; et al. Targeting Potential Drivers of COVID-19: Neutrophil Extracellular Traps. J. Exp. Med. 2020, 217, e20200652. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil Extracellular Traps in COVID-19. JCI Insight 2020, 5, e138999. [Google Scholar] [CrossRef] [PubMed]
- Zolfaghari Emameh, R.; Nosrati, H.; Eftekhari, M.; Falak, R.; Khoshmirsafa, M. Expansion of Single Cell Transcriptomics Data of SARS-CoV Infection in Human Bronchial Epithelial Cells to COVID-19. Biol. Proced. Online 2020, 22, 16. [Google Scholar] [CrossRef] [PubMed]
- Aghamohamadi, N.; Shahba, F.; Zarezadeh Mehrabadi, A.; Khorramdelazad, H.; Karimi, M.; Falak, R.; Emameh, R.Z. Age-Dependent Immune Responses in COVID-19-Mediated Liver Injury: Focus on Cytokines. Front. Endocrinol. 2023, 14, 1139692. [Google Scholar] [CrossRef] [PubMed]
- Jouan, Y.; Guillon, A.; Gonzalez, L.; Perez, Y.; Boisseau, C.; Ehrmann, S.; Ferreira, M.; Daix, T.; Jeannet, R.; François, B.; et al. Phenotypical and Functional Alteration of Unconventional T Cells in Severe COVID-19 Patients. J. Exp. Med. 2020, 217, e20200872. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yang, P.; Zhao, Y.; Zhuang, Z.; Wang, Z.; Song, R.; Zhang, J.; Liu, C.; Gao, Q.; Xu, Q.; et al. Single-Cell Sequencing of Peripheral Mononuclear Cells Reveals Distinct Immune Response Landscapes of COVID-19 and Influenza Patients. Immunity 2020, 53, 685–696.e3. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Panzer, U.; Bonn, S.; Krebs, C.F. Single-Cell Biology to Decode the Immune Cellular Composition of Kidney Inflammation. Cell Tissue Res. 2021, 385, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Qiu, Y.; Tu, G.; Yang, C. Single-Cell RNA Sequencing in Immunology. Curr. Genom. 2020, 21, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhou, X. Impact of Single-Cell RNA Sequencing on Understanding Immune Regulation. J. Cell. Mol. Med. 2022, 26, 4645–4657. [Google Scholar] [CrossRef] [PubMed]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martínez-Colón, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T.; et al. A Single-Cell Atlas of the Peripheral Immune Response in Patients with Severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Y.; Wang, X.-M.; Xing, X.; Xu, Z.; Zhang, C.; Song, J.-W.; Fan, X.; Xia, P.; Fu, J.-L.; Wang, S.-Y.; et al. Single-Cell Landscape of Immunological Responses in Patients with COVID-19. Nat. Immunol. 2020, 21, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419–1440.e23. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Giri, A.K.; Aittokallio, T. Fully-Automated and Ultra-Fast Cell-Type Identification Using Specific Marker Combinations from Single-Cell Transcriptomic Data. Nat. Commun. 2022, 13, 1246. [Google Scholar] [CrossRef] [PubMed]
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil Kinetics in Health and Disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Ciarambino, T.; Para, O.; Giordano, M. Immune System and COVID-19 by Sex Differences and Age. Women’s Health 2021, 17, 17455065211022262. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Ngwa, C.; Morales Scheihing, D.A.; Al Mamun, A.; Ahnstedt, H.W.; Finger, C.E.; Colpo, G.D.; Sharmeen, R.; Kim, Y.; Choi, H.A.; et al. Sex Differences in the Immune Response to Acute COVID-19 Respiratory Tract Infection. Biol. Sex Differ. 2021, 12, 66. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaivode, K.; Saksis, R.; Litvina, H.D.; Niedra, H.; Spriņģe, M.L.; Krūmiņa, U.; Kloviņš, J.; Rovite, V. Single-Cell RNA Sequencing Reveals Alterations in Patient Immune Cells with Pulmonary Long COVID-19 Complications. Curr. Issues Mol. Biol. 2024, 46, 461-468. https://doi.org/10.3390/cimb46010029
Vaivode K, Saksis R, Litvina HD, Niedra H, Spriņģe ML, Krūmiņa U, Kloviņš J, Rovite V. Single-Cell RNA Sequencing Reveals Alterations in Patient Immune Cells with Pulmonary Long COVID-19 Complications. Current Issues in Molecular Biology. 2024; 46(1):461-468. https://doi.org/10.3390/cimb46010029
Chicago/Turabian StyleVaivode, Kristīne, Rihards Saksis, Helēna Daiga Litvina, Helvijs Niedra, Marta Līva Spriņģe, Una Krūmiņa, Jānis Kloviņš, and Vita Rovite. 2024. "Single-Cell RNA Sequencing Reveals Alterations in Patient Immune Cells with Pulmonary Long COVID-19 Complications" Current Issues in Molecular Biology 46, no. 1: 461-468. https://doi.org/10.3390/cimb46010029