
Drought Tolerance of Legumes: Physiology and the Role of the Microbiome



Abstract
:1. Introduction
2. Common Drought-Adaptation Mechanisms in Plants
2.1. Role of Phytohormones
2.2. Osmoprotection System
2.3. Reactive Oxygen Species (ROS)
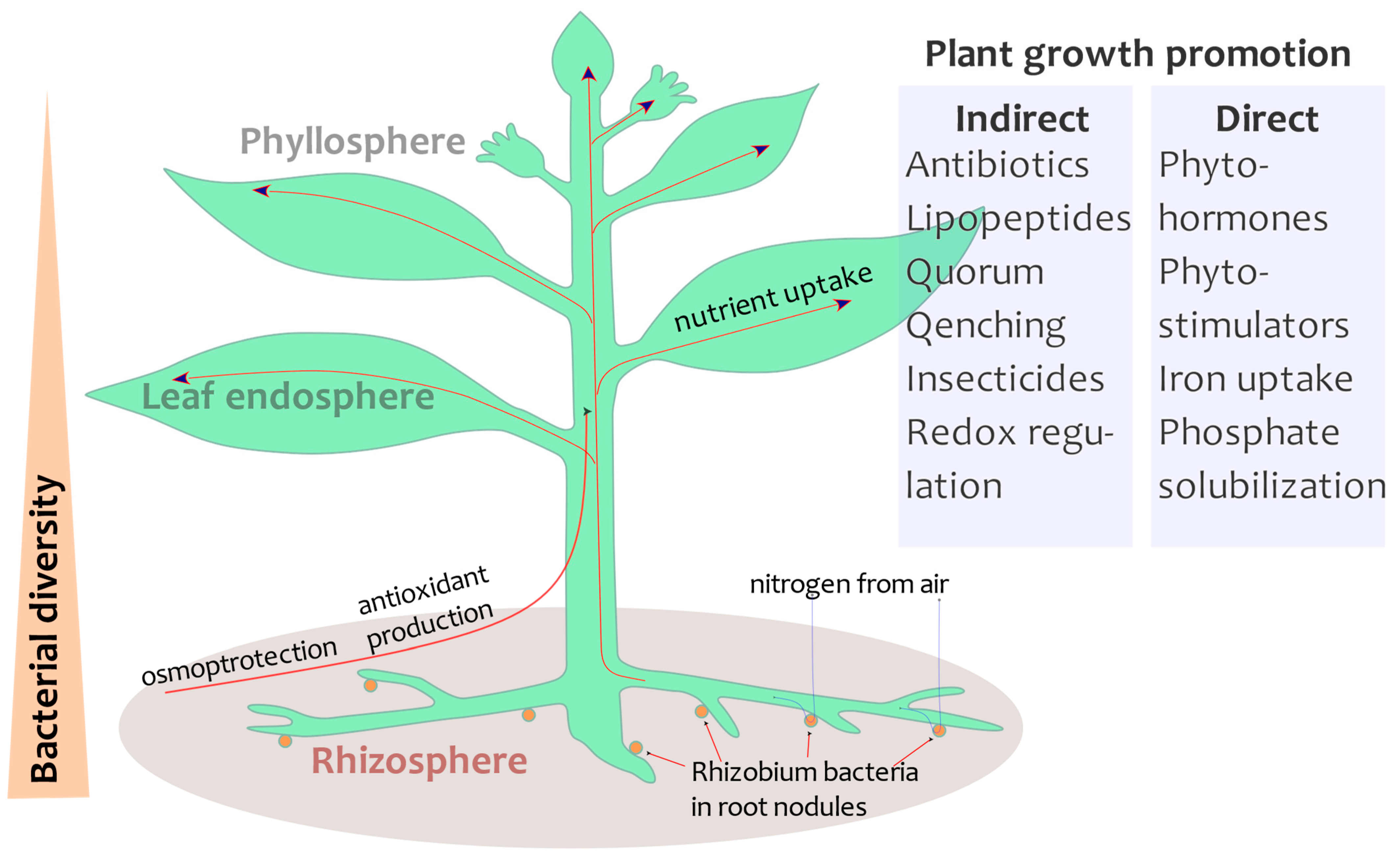
3. The Role of the Beneficial Microbes to Face Drought
4. Microbiome Engineering
5. Microbiome Modulation of Fabaceae Plants

{kind=link}
| Plant Host | Microbial Mixture Components | Reference |
|---|---|---|
| Astragalus adsurgens | A. chlamydospora, S. kiliense, and Monosporascus sp. | [64] |
| Alhagi sparsifolia | Pseudomonas strain LTGT-11-2Z | [63] |
| Vicia faba | Rhizobium leguminosarum (Rl) and Pseudomonas putida (Pp) | [69] |
| Glycine max L. | Azotobacter chroococcum (Az) and Piriformospora indica (Pi) | [70] |
| Glycyrrhiza uralensis | Bacillus amyloliquefaciens strain FZB42 | [72] |
| Chickpea (Cicer arietinum L.) | Mesorhizobium sp. UTADM31, Burkholderia sp. UTADB34 and Pseudomonas sp. UTAD11.3, Funneliformis geosporum and Claroideoglomus claroideum | [71] |
| Vigna radiate | Pseudomonas aeruginosa, the strains MK513745, MK513746, MK513747, MK513748, and MK513749 | [62] |
| Lupinus albus | Bacillus pretiosus SAICEU11T, Pseudomonas agronomica SAICEU22T | [68] |
| Glycine max L. | Bacillus amyloliquefaciens MN592674B, Mycorrhizal spores (Acaulospora laevis, Septoglomus deserticola, Rhizophagus irregularis) | [74] |
| Pisum sativum and Phaseolus vulgaris | Pseudomonas fluorescens DR397 | [65] |
| Pisum sativum | Rhizobium leguminosarum bv. viciae 1066S | [66] |
| Vigna unguiculata | Pseudomonas sp. IESDJP-V1, Pseudomonas sp. IESDJP-V2, Serratia marcescens IESDJP-V3, Bacillus cereus IESDJP-V4, Ochrobactrum sp. IESDJP-V5, Azospirillum brasilense MTCC-4037, Paenibacillus polymyxa BHUPSB17 | [75] |
6. Legume Microbiome for Different Watering Regimes
| Plant Host | Major Taxa | Water Regime/Soil Type | Reference |
| Arachis hypogaea | Acaulospora, Glomus, Gigaspora | Well-watered 45%/30%; medium drought 30%/15% 1 | [81] |
| Cicer arietinum | Azotobacter chroococcum, Bacillus subtilis, Pseudomonas aeruginosa, Bacillus pumilis2 | Drought stress was created by adding 32.6% of polyethylene glycol (PEG 6000) | [77] |
| Vigna mungo, Phaseolus vulgari | Bacillus cereus, Bacillus haynesii2 | In vitro drought tolerance study was conducted using PEG 6000 | [78] |
| Phaseolus vulgaris L. | Pseudomonas, Bacillus, Acinetobacter, Raoultella, Escherichia-Shigella | Ample water (300 mL/day); 66% less water (100 mL/day); Hoagland solution (300 mL/day) | [79] |
| Arachis hypogaea L. | Actinobacteria, Proteobacteria, Saccharibacteria, Chloroflexi, Acidobacteria, and Cyanobacteria | 85% of field capacity (control); 45%–drought | [80] |
| Arachis hypogaea L. | Leptospaerulina, Cladosporium, Apiotrichum; | Field and pot experiment | [82] |
| Arachis hypogaea L. | Actinobacteria, Proteobacteria, Acidobacteria, Chloroflexi; Ascomycota, Basidiomycota, Mortierellomycota | Natural soil | [76] |
| Trigonella foenum-graecum | Ensifer meliloti2 | No data | [83] |
| Macrotyloma uniflorum | Proteobacteria, Actinobacteria, Firmicutes, Acidobacteria, Bacteroidetes, Planctomycetes, Gemmatimonadetes | Bulk soil, rhizosphere soil, root nodules and seed samples | [84] |
| Vigna subterranea | Actinobacteria, Proteobacteria, Acidobacteria | Bulk soil | [85] |
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lesk, C.; Anderson, W.; Rigden, A.; Coast, O.; Jägermeyr, J.; McDermid, S.; Davis, K.F.; Konar, M. Compound heat and moisture extreme impacts on global crop yields under climate change. Nat. Rev. Earth Environ. 2022, 3, 872–889. [Google Scholar] [CrossRef]
- Islam, M.S.; Fahad, S.; Hossain, A.; Chowdhury, M.K.; Iqbal, M.A.; Dubey, A.; Kumar, A.; Rajendran, K.; Danish, S.; Rahman, M.H.U.; et al. Legumes under Drought Stress: Plant Responses, Adaptive Mechanisms, and Management Strategies in Relation to Nitrogen Fixation. In Engineering Tolerance in Crop Plants Against Abiotic Stress; CRC Press: Boca Raton, FL, USA, 2021; ISBN 9781000462135. [Google Scholar]
- Zhang, H.; Sun, X.; Dai, M. Improving crop drought resistance with plant growth regulators and rhizobacteria: Mechanisms, applications, and perspectives. Plant Commun. 2022, 3, 100228. [Google Scholar] [CrossRef] [PubMed]
- Dubey, A.; Kumar, A.; Khan, M.L. Role of Biostimulants for Enhancing Abiotic Stress Tolerance in Fabaceae Plants. In The Plant Family Fabaceae; Springer: Singapore, 2020; ISBN 9789811547522. [Google Scholar]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research Progress and Perspective on Drought Stress in Legumes: A Review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatun, M.; Sarkar, S.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Fahad, S.; Datta, R.; Islam, A.K.M.A. Drought stress in grain legumes: Effects, tolerance mechanisms and management. Agronomy 2021, 11, 2374. [Google Scholar] [CrossRef]
- Ullah, A.; Farooq, M. The challenge of drought stress for grain legumes and options for improvement. Arch. Agron. Soil Sci. 2022, 68, 1601–1618. [Google Scholar] [CrossRef]
- Monteoliva, M.I.; Ruiz, O.A.; Li, F. Editorial: Legumes and their microbiome in climate change mitigation. Front. Plant Sci. 2023, 14, 1220535. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, P.; Kumar, R.; Sharma, V.; Bhardwaj, R.; Sharma, I. Role of reactive oxygen species in the regulation of abiotic stress tolerance in legumes. In Abiotic Stress and Legumes: Tolerance and Management; Academic Press: Cambridge, MA, USA, 2021; ISBN 9780128153550. [Google Scholar]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Thakro, V.; Malik, N.; Basu, U.; Srivastava, R.; Narnoliya, L.; Daware, A.; Varshney, N.; Mohanty, J.K.; Bajaj, D.; Dwivedi, V.; et al. A superior gene allele involved in abscisic acid signaling enhances drought tolerance and yield in chickpea. Plant Physiol. 2023, 191, 1884–1912. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Liang, D.W.; Bian, X.H.; Shen, M.; Xiao, J.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Lv, J.; Chen, X.; et al. GmWRKY54 improves drought tolerance through activating genes in abscisic acid and Ca2+ signaling pathways in transgenic soybean. Plant J. 2019, 100, 384–398. [Google Scholar] [CrossRef]
- Soni, P.; Nair, R.; Jain, S.; Sahu, R.K.; Banjare, K.; Sahu, K. Assessment of Salicylic Acid Impacts on Physiological and Biochemical Characteristics under Water Deficit Stress on Pea (Pisum sativum L. var. Kashi Nandni). Int. J. Environ. Clim. Chang. 2022, 12, 3034–3041. [Google Scholar] [CrossRef]
- Boulahia, K.; Ould Said, C.; Abrous-Belbachir, O. Exogenous Application of Salicylic Acid Improve Growth and Some Physio-Biochemical Parameters in Herbicide Stressed Phaseolus vulgaris L. Gesunde Pflanz. 2023. [Google Scholar] [CrossRef]
- Imran, M.; Latif Khan, A.; Shahzad, R.; Aaqil Khan, M.; Bilal, S.; Khan, A.; Kang, S.M.; Lee, I.J. Exogenous melatonin induces drought stress tolerance by promoting plant growth and antioxidant defence system of soybean plants. AoB Plants 2021, 13, plab026. [Google Scholar] [CrossRef] [PubMed]
- Anitha, K.; Senthil, A.; Kalarani, M.K.; Senthil, N.; Marimuthu, S.; Djanaguiraman, M.; Umapathi, M. Exogenous melatonin improves seed germination and seedling growth in greengram under drought stress. J. Appl. Nat. Sci. 2022, 14, 3818. [Google Scholar] [CrossRef]
- Kuppusamy, A.; Alagarswamy, S.; Karuppusami, K.M.; Maduraimuthu, D.; Natesan, S.; Ramalingam, K.; Muniyappan, U.; Subramanian, M.; Kanagarajan, S. Melatonin Enhances the Photosynthesis and Antioxidant Enzyme Activities of Mung Bean under Drought and High-Temperature Stress Conditions. Plants 2023, 12, 2535. [Google Scholar] [CrossRef] [PubMed]
- Pandiyan, M.; Sivaji, M.; Yuvaraj, M.; Krishnaveni, A.; Sivakumar, C.; Jamuna, E. Molecular and Physiological Approaches for Effective Management of Drought in Black Gram. In Legumes: Physiology and Molecular Biology of Abiotic Stress Tolerance; Springer: Singapore, 2023; ISBN 9789811958175. [Google Scholar]
- Liang, Y.; Wei, G.; Ning, K.; Li, M.; Zhang, G.; Luo, L.; Zhao, G.; Wei, J.; Liu, Y.; Dong, L.; et al. Increase in carbohydrate content and variation in microbiome are related to the drought tolerance of Codonopsis pilosula. Plant Physiol. Biochem. 2021, 165, 19–35. [Google Scholar] [CrossRef]
- Abid, G.; Ouertani, R.N.; Ghouili, E.; Muhovski, Y.; Jebara, S.H.; Abdelkarim, S.; Chaieb, O.; Ben Redjem, Y.; El Ayed, M.; Barhoumi, F.; et al. Exogenous application of spermidine mitigates the adverse effects of drought stress in faba bean (Vicia faba L.). Funct. Plant Biol. 2022, 49, 405–420. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Anandham, R.; Senthilkumar, M.; Venkatramanan, V. Adaptation Mechanism of Methylotrophic Bacteria to Drought Condition and Its Strategies in Mitigating Plant Stress Caused by Climate Change. In Exploring Synergies and Trade-Offs between Climate Change and the Sustainable Development Goals; Springer: Singapore, 2021. [Google Scholar]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M.; Johnson, M.T.J. Assembly and ecological function of the root microbiome across angiosperm plant species. Proc. Natl. Acad. Sci. USA 2018, 115, E1157–E1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganie, S.A.; Bhat, J.A.; Devoto, A. The influence of endophytes on rice fitness under environmental stresses. Plant Mol. Biol. 2022, 109, 447–467. [Google Scholar] [CrossRef]
- Sessitsch, A.; Pfaffenbichler, N.; Mitter, B. Microbiome Applications from Lab to Field: Facing Complexity. Trends Plant Sci. 2019, 24, 194–198. [Google Scholar] [CrossRef]
- Sarkar, S.; Khatun, M.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Danish, S.; Datta, R.; Islam, A.K.M.A. Abiotic stresses: Alteration of composition and grain quality in food legumes. Agronomy 2021, 11, 2238. [Google Scholar] [CrossRef]
- Vorobyov, N.I.; Provorov, N.A. Simulation of the evolution of the legume-rhizobia symbiosis under the conditions of ecological instability. Russ. J. Genet. Appl. Res. 2015, 5, 91–101. [Google Scholar] [CrossRef]
- Jacques, C.; Salon, C.; Barnard, R.L.; Vernoud, V.; Prudent, M. Drought stress memory at the plant cycle level: A review. Plants 2021, 10, 1873. [Google Scholar] [CrossRef] [PubMed]
- Santos-Medellín, C.; Liechty, Z.; Edwards, J.; Nguyen, B.; Huang, B.; Weimer, B.C.; Sundaresan, V. Prolonged drought imparts lasting compositional changes to the rice root microbiome. Nat. Plants 2021, 7, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Duan, G.-L.; García-Palacios, P.; Yang, G.; Cui, H.-L.; Yan, M.; Yin, Y.; Yi, X.-Y.; Li, L.; Delgado-Baquerizo, M.; et al. Environmental factors and host genotype control foliar epiphytic microbial community of wild soybeans across China. Front. Microbiol. 2023, 14, 1065302. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G. How plants recruit their microbiome? New insights into beneficial interactions. J. Adv. Res. 2022, 40, 45–58. [Google Scholar] [CrossRef]
- Diáz-Cruz, G.A.; Cassone, B.J. Changes in the phyllosphere and rhizosphere microbial communities of soybean in the presence of pathogens. FEMS Microbiol. Ecol. 2022, 98, fiac022. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Rizaludin, M.S.; Stopnisek, N.; Raaijmakers, J.M.; Garbeva, P. The chemistry of stress: Understanding the ‘cry for help’ of plant roots. Metabolites 2021, 11, 357. [Google Scholar] [CrossRef]
- Lyu, D.; Smith, D.L. The root signals in rhizospheric inter-organismal communications. Front. Plant Sci. 2022, 13, 1064058. [Google Scholar] [CrossRef]
- Hao, Z.; Xie, W.; Jiang, X.; Wu, Z.; Zhang, X.; Chen, B. Arbuscular mycorrhizal fungus improves Rhizobium–glycyrrhiza seedling symbiosis under drought stress. Agronomy 2019, 9, 572. [Google Scholar] [CrossRef] [Green Version]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.; Wang, C.; Zhang, F.; Chen, L.; Sun, Z.; Cai, Y.; Luo, Y.; Liao, J.; Wang, Y.; Cha, Y.; et al. A nitrogen fixing symbiosis-specific pathway required for legume flowering. Sci. Adv. 2023, 9, ade1150. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Y.; Pei, K.; Zhou, J.; Peixoto, L.; Gunina, A.; Zeng, Z.; Zang, H.; Rasmussen, J.; Kuzyakov, Y. Nitrogen rhizodeposition by legumes and its fate in agroecosystems: A field study and literature review. Land Degrad. Dev. 2021, 32, 410–419. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.Y.; Wu, H.M.; Zhang, F.F.; Li, C.J.; Li, X.X.; Lambers, H.; Li, L. Root exudates drive interspecific facilitation by enhancing nodulation and N2 fixation. Proc. Natl. Acad. Sci. USA 2016, 113, 6496–6501. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, S.E.; De Beuf, K.; Vekeman, B.; Willems, A. A large diversity of non-rhizobial endophytes found in legume root nodules in Flanders (Belgium). Soil Biol. Biochem. 2015, 83, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Hidalgo, P.; Hirsch, A.M. Erratum: The Nodule Microbiome: N2-Fixing Rhizobia Do Not Live Alone. Phytobiomes J. 2022, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Tsiknia, M.; Tsikou, D.; Papadopoulou, K.K.; Ehaliotis, C. Multi-species relationships in legume roots: From pairwise legume-symbiont interactions to the plant—Microbiome—Soil continuum. FEMS Microbiol. Ecol. 2021, 97, fiaa222. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Tyagi, A.; Park, S.; Mir, R.A.; Mushtaq, M.; Bhat, B.; Mahmoudi, H.; Bae, H. Deciphering the plant microbiome to improve drought tolerance: Mechanisms and perspectives. Environ. Exp. Bot. 2022, 201, 104933. [Google Scholar] [CrossRef]
- Phour, M.; Sindhu, S.S. Mitigating abiotic stress: Microbiome engineering for improving agricultural production and environmental sustainability. Planta 2022, 256, 85. [Google Scholar] [CrossRef] [PubMed]
- Panke-Buisse, K.; Lee, S.; Kao-Kniffin, J. Cultivated Sub-Populations of Soil Microbiomes Retain Early Flowering Plant Trait. Microb. Ecol. 2017, 73, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech 2017, 7, 315. [Google Scholar] [CrossRef]
- Monteoliva, M.I.; Valetti, L.; Taurian, T.; Crociara, S.C.; Guzzo, M.C. Synthetic Communities of Bacterial Endophytes to Improve the Quality and Yield of Legume Crops. In Legumes Research; InTechOpen: London, UK, 2022; Volume 1. [Google Scholar]
- Quides, K.W.; Atamian, H.S. A microbiome engineering framework to evaluate rhizobial symbionts of legumes. Plant Soil 2021, 463, 631–642. [Google Scholar] [CrossRef]
- Jochum, M.D.; McWilliams, K.L.; Pierson, E.A.; Jo, Y.K. Host-mediated microbiome engineering (HMME) of drought tolerance in the wheat rhizosphere. PLoS ONE 2019, 14, e0225933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, B.L.; de Cassia Pessotti, R.; Fischer, M.S.; Collins, A.; El-Hifnawi, L.; Liu, M.D.; Traxler, M.F. Cooperation, competition, and specialized metabolism in a simplified root nodule microbiome. MBio 2020, 11, e01917-20. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Álvarez, C.; Peimbert, M.; Rodríguez-Martin, P.; Trejo-Aguilar, D.; Alcaraz, L.D. A Study of Microbial Diversity in a Biofertilizer Consortium. bioRxiv 2023. [Google Scholar] [CrossRef]
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Molina-Romero, D.; Baez, A.; Quintero-Hernández, V.; Castañeda-Lucio, M.; Fuentes-Ramírez, L.E.; del Rocío Bustillos-Cristales, M.; Rodríguez-Andrade, O.; Morales-García, Y.E.; Munive, A.; Muñoz-Rojas, J. Compatible bacterial mixture, tolerant to desiccation, improves maize plant growth. PLoS ONE 2017, 12, e0187913. [Google Scholar] [CrossRef]
- Chang, C.Y.; Vila, J.C.C.; Bender, M.; Li, R.; Mankowski, M.C.; Bassette, M.; Borden, J.; Golfier, S.; Sanchez, P.G.L.; Waymack, R.; et al. Engineering complex communities by directed evolution. Nat. Ecol. Evol. 2021, 5, 1011–1023. [Google Scholar] [CrossRef]
- Mueller, U.G.; Juenger, T.E.; Kardish, M.R.; Carlson, A.L.; Burns, K.M.; Edwards, J.A.; Smith, C.C.; Fang, C.-C.; Des Marais, D.L. Artificial Selection on Microbiomes To Breed Microbiomes That Confer Salt Tolerance to Plants. mSystems 2021, 6, e01125-21. [Google Scholar] [CrossRef]
- Jacquiod, S.; Spor, A.; Wei, S.; Munkager, V.; Bru, D.; Sørensen, S.J.; Salon, C.; Philippot, L.; Blouin, M. Artificial selection of stable rhizosphere microbiota leads to heritable plant phenotype changes. Ecol. Lett. 2022, 25, 189–201. [Google Scholar] [CrossRef]
- Xu, L.; Naylor, D.; Dong, Z.; Simmons, T.; Pierroz, G.; Hixson, K.K.; Kim, Y.M.; Zink, E.M.; Engbrecht, K.M.; Wang, Y.; et al. Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E4284–E4293. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Coleman-Derr, D. Causes and consequences of a conserved bacterial root microbiome response to drought stress. Curr. Opin. Microbiol. 2019, 49, 1–6. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Chiniquy, D.; Pierroz, G.; Deng, S.; Gao, C.; Diamond, S.; Simmons, T.; Wipf, H.M.L.; Caddell, D.; et al. Genome-resolved metagenomics reveals role of iron metabolism in drought-induced rhizosphere microbiome dynamics. Nat. Commun. 2021, 12, 3209. [Google Scholar] [CrossRef] [PubMed]
- Inbaraj, M.P. Plant-Microbe Interactions in Alleviating Abiotic Stress—A Mini Review. Front. Agron. 2021, 3, 667903. [Google Scholar] [CrossRef]
- Uzma, M.; Iqbal, A.; Hasnain, S. Drought tolerance induction and growth promotion by indole acetic acid producing Pseudomonas aeruginosa in Vigna radiata. PLoS ONE 2022, 17, e0262932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, W.; Li, Q.; Cui, R.; Wang, Z.; Wang, Y.; Zhang, Y.Z.; Ding, W.; Shen, X. Deciphering the root endosphere microbiome of the desert plant Alhagi sparsifolia for drought resistance-promoting bacteria. Appl. Environ. Microbiol. 2020, 86, e02863-19. [Google Scholar] [CrossRef]
- Zuo, Y.; Hu, Q.; Liu, J.; He, X. Relationship of root dark septate endophytes and soil factors to plant species and seasonal variation in extremely arid desert in Northwest China. Appl. Soil Ecol. 2022, 175, 104454. [Google Scholar] [CrossRef]
- Nishu, S.D.; No, J.H.; Lee, T.K. Transcriptional Response and Plant Growth Promoting Activity of Pseudomonas fluorescens DR397 under Drought Stress Conditions. Microbiol. Spectr. 2022, 10, e00979-22. [Google Scholar] [CrossRef]
- Belimov, A.A.; Zinovkina, N.Y.; Safronova, V.I.; Litvinsky, V.A.; Nosikov, V.V.; Zavalin, A.A.; Tikhonovich, I.A. Rhizobial ACC deaminase contributes to efficient symbiosis with pea (Pisum sativum L.) under single and combined cadmium and water deficit stress. Environ. Exp. Bot. 2019, 167, 103859. [Google Scholar] [CrossRef]
- Kichko, A.A.; Gladkov, G.V.; Ulianich, P.S.; Safronova, V.I.; Pinaev, A.G.; Sekste, E.A.; Belimov, A.A.; Andronov, E.E. Water Stress, Cadmium, and Plant Genotype Modulate the Rhizosphere Microbiome of Pisum sativum L. Plants 2022, 11, 3013. [Google Scholar] [CrossRef]
- Robas Mora, M.; Fernández Pastrana, V.M.; Oliva, L.L.G.; Lobo, A.P.; Jiménez Gómez, P.A. Plant growth promotion of the forage plant Lupinus albus Var. Orden Dorado using Pseudomonas agronomica sp. nov. and Bacillus pretiosus sp. nov. added over a valorized agricultural biowaste. Front. Microbiol. 2023, 13, 1046201. [Google Scholar] [CrossRef]
- Mansour, E.; Mahgoub, H.A.M.; Mahgoub, S.A.; El-Sobky, E.S.E.A.; Abdul-Hamid, M.I.; Kamara, M.M.; AbuQamar, S.F.; El-Tarabily, K.A.; Desoky, E.S.M. Enhancement of drought tolerance in diverse Vicia faba cultivars by inoculation with plant growth-promoting rhizobacteria under newly reclaimed soil conditions. Sci. Rep. 2021, 11, 24142. [Google Scholar] [CrossRef]
- Yaghoubian, I.; Modarres-Sanavy, S.A.M.; Smith, D.L. Plant growth promoting microorganisms (PGPM) as an eco-friendly option to mitigate water deficit in soybean (Glycine max L.): Growth, physio-biochemical properties and oil content. Plant Physiol. Biochem. 2022, 191, 55–66. [Google Scholar] [CrossRef]
- Laranjeira, S.; Fernandes-Silva, A.; Reis, S.; Torcato, C.; Raimundo, F.; Ferreira, L.; Carnide, V.; Marques, G. Inoculation of plant growth promoting bacteria and arbuscular mycorrhizal fungi improve chickpea performance under water deficit conditions. Appl. Soil Ecol. 2021, 164, 103927. [Google Scholar] [CrossRef]
- Yue, L.; Uwaremwe, C.; Tian, Y.; Liu, Y.; Zhao, X.; Zhou, Q.; Wang, Y.; Zhang, Y.; Liu, B.; Cui, Z.; et al. Bacillus amyloliquefaciens Rescues Glycyrrhizic Acid Loss Under Drought Stress in Glycyrrhiza uralensis by Activating the Jasmonic Acid Pathway. Front. Microbiol. 2022, 12, 4211. [Google Scholar] [CrossRef]
- He, C.; Liu, C.; Liu, H.; Wang, W.; Hou, J.; Li, X. Dual inoculation of dark septate endophytes and Trichoderma viride drives plant performance and rhizosphere microbiome adaptations of Astragalus mongholicus to drought. Environ. Microbiol. 2022, 24, 324–340. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Alhaj Hamoud, Y.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef]
- Verma, J.P.; Jaiswal, D.K.; Gaurav, A.K.; Mukherjee, A.; Krishna, R.; Prudêncio de Araujo Pereira, A. Harnessing bacterial strain from rhizosphere to develop indigenous PGPR consortium for enhancing lobia (Vigna unguiculata) production. Heliyon 2023, 9, e13804. [Google Scholar] [CrossRef]
- Li, M.; Wei, Y.; Yin, Y.; Ding, H.; Zhu, W.; Zhou, Y. The Effect of Intercropping Mulberry (Morus alba L.) with Peanut (Arachis hypogaea L.), on the Soil Rhizosphere Microbial Community. Forests 2022, 13, 1757. [Google Scholar] [CrossRef]
- Pandey, S.; Gupta, S.; Ramawat, N. Unravelling the potential of microbes isolated from rhizospheric soil of chickpea (Cicer arietinum) as plant growth promoter. 3 Biotech 2019, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Andy, A.K.; Rajput, V.D.; Burachevskaya, M.; Gour, V.S. Exploring the Identity and Properties of Two Bacilli Strains and their Potential to Alleviate Drought and Heavy Metal Stress. Horticulturae 2023, 9, 46. [Google Scholar] [CrossRef]
- Bintarti, A.F.; Kearns, P.J.; Sulesky-Grieb, A.; Shade, A. Abiotic Treatment to Common Bean Plants Results in an Altered Endophytic Seed Microbiome. Microbiol. Spectr. 2022, 10, e0021021. [Google Scholar] [CrossRef]
- Dai, L.; Zhang, G.; Yu, Z.; Ding, H.; Xu, Y.; Zhang, Z. Effect of drought stress and developmental stages on microbial community structure and diversity in peanut rhizosphere soil. Int. J. Mol. Sci. 2019, 20, 2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.J.; Song, S.L.; Ma, C.Y.; Zhang, W.; Sun, K.; Tang, M.J.; Xie, X.G.; Fan, K.K.; Dai, C.C. Endophytic fungus improves peanut drought resistance by reassembling the root-dwelling community of arbuscular mycorrhizal fungi. Fungal Ecol. 2020, 48, 100993. [Google Scholar] [CrossRef]
- Luo, X.; Bai, Y.N.; Sun, K.; Zhang, W.; Dai, C.C. Reduced pollen activity in peanut (Arachis hypogaea L.) by long-term monocropping is linked to flower water deficit. Plant Soil 2023, 482, 427–450. [Google Scholar] [CrossRef]
- Khairnar, M.; Hagir, A.; Parmar, K.; Sayyed, R.Z.; James, E.K.; Rahi, P. Phylogenetic diversity and plant growth-promoting activities of rhizobia nodulating fenugreek (Trigonella foenum-graecum Linn.) cultivated in different agroclimatic regions of India. FEMS Microbiol. Ecol. 2022, 98, fiac014. [Google Scholar] [CrossRef]
- Evangilene, S.; Uthandi, S. A 16S rRNA amplicon approach to the structural and functional diversity of bacterial communities associated with horse gram crop for drought mitigation and sustainable productivity. J. Appl. Microbiol. 2022, 133, 3777–3789. [Google Scholar] [CrossRef]
- Ajilogba, C.F.; Olanrewaju, O.S.; Babalola, O.O. Plant Growth Stage Drives the Temporal and Spatial Dynamics of the Bacterial Microbiome in the Rhizosphere of Vigna subterranea. Front. Microbiol. 2022, 13, 106. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrushin, I.S.; Vasilev, I.A.; Markova, Y.A. Drought Tolerance of Legumes: Physiology and the Role of the Microbiome. Curr. Issues Mol. Biol. 2023, 45, 6311-6324. https://doi.org/10.3390/cimb45080398
Petrushin IS, Vasilev IA, Markova YA. Drought Tolerance of Legumes: Physiology and the Role of the Microbiome. Current Issues in Molecular Biology. 2023; 45(8):6311-6324. https://doi.org/10.3390/cimb45080398
Chicago/Turabian StylePetrushin, Ivan S., Ilia A. Vasilev, and Yulia A. Markova. 2023. "Drought Tolerance of Legumes: Physiology and the Role of the Microbiome" Current Issues in Molecular Biology 45, no. 8: 6311-6324. https://doi.org/10.3390/cimb45080398