
Interstitial Cells of Cajal and Enteric Nervous System in Gastrointestinal and Neurological Pathology, Relation to Oxidative Stress
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
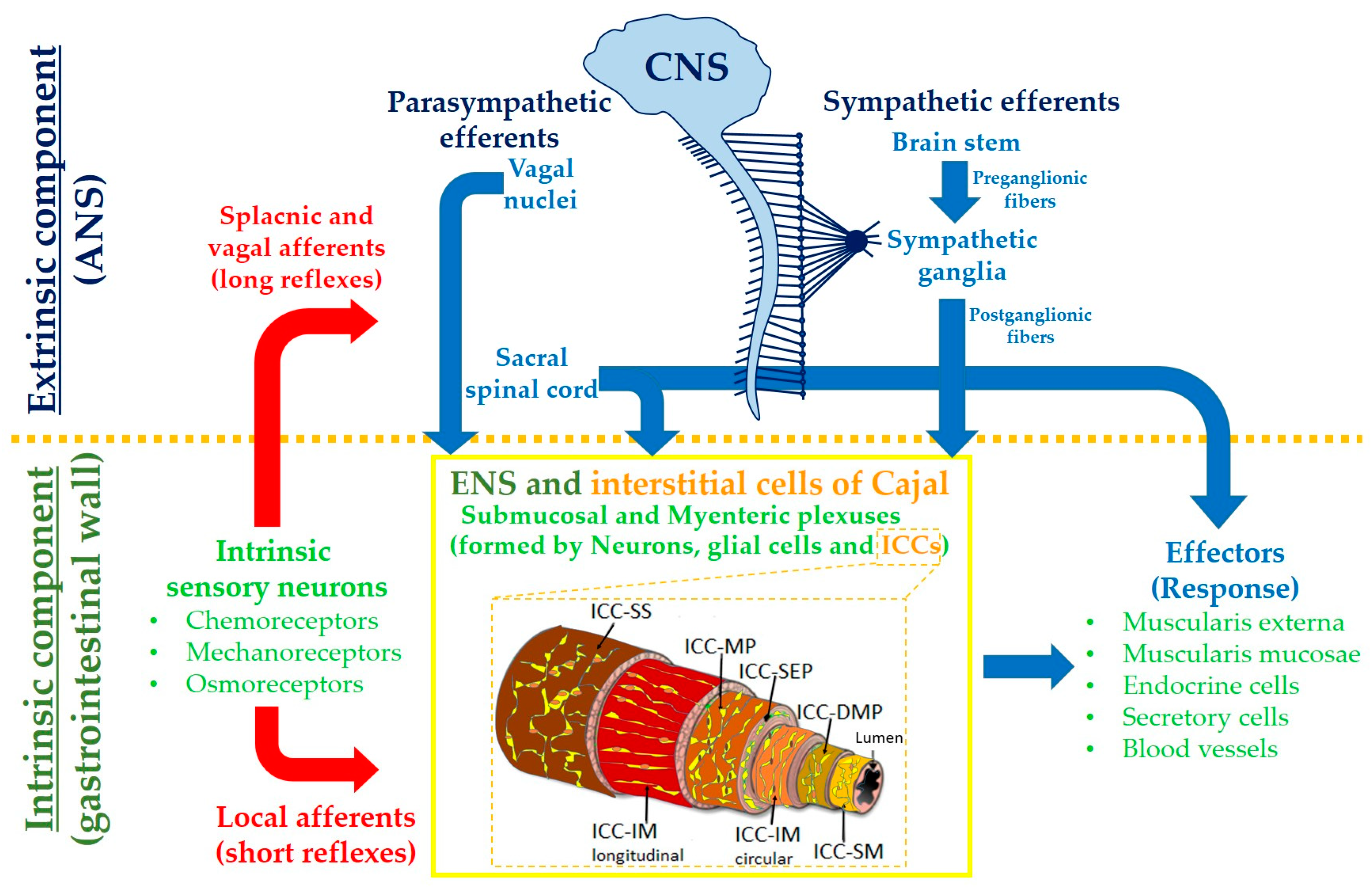
:1. Introduction
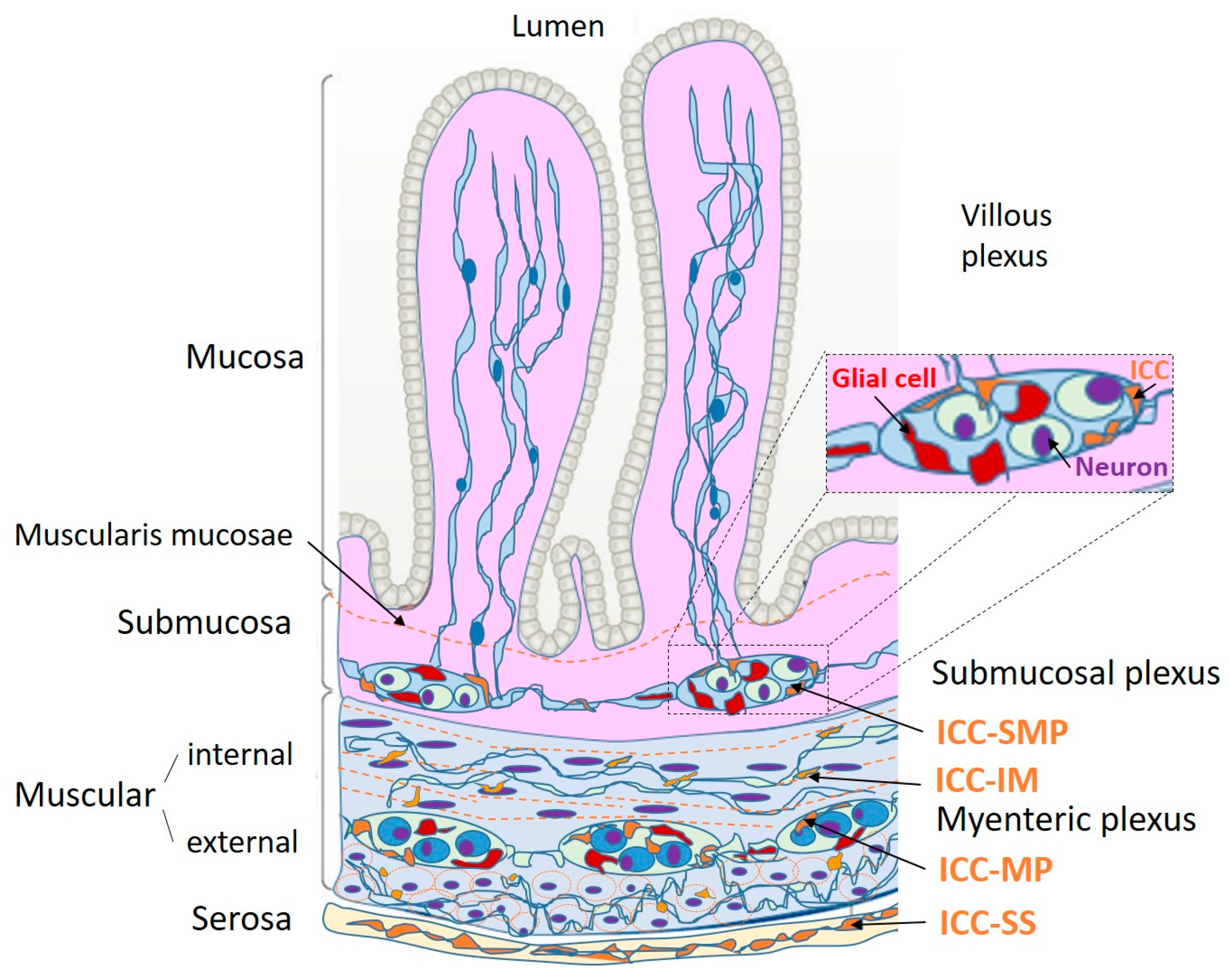
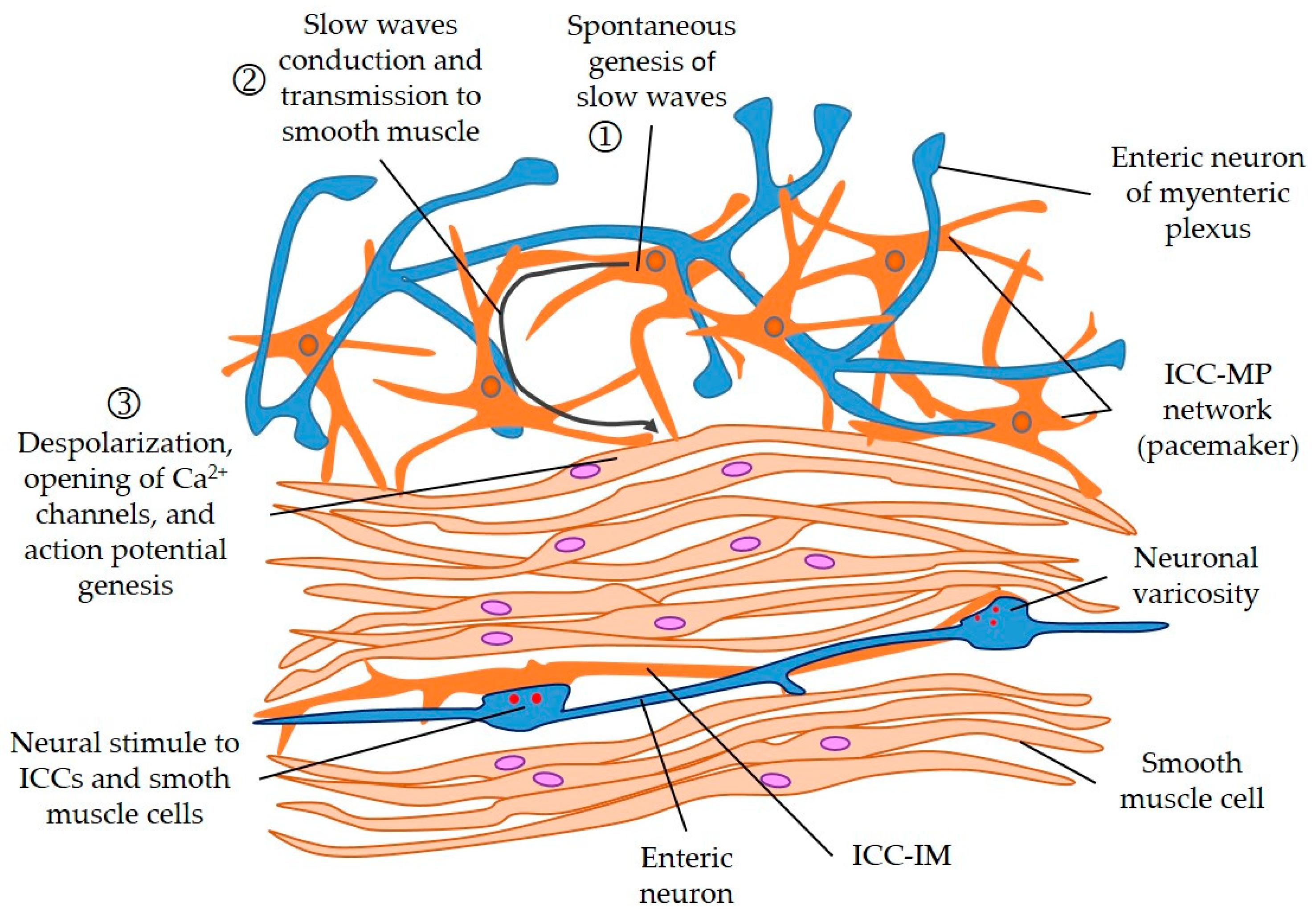
2. The Interstitial Cells of Cajal in the Gut
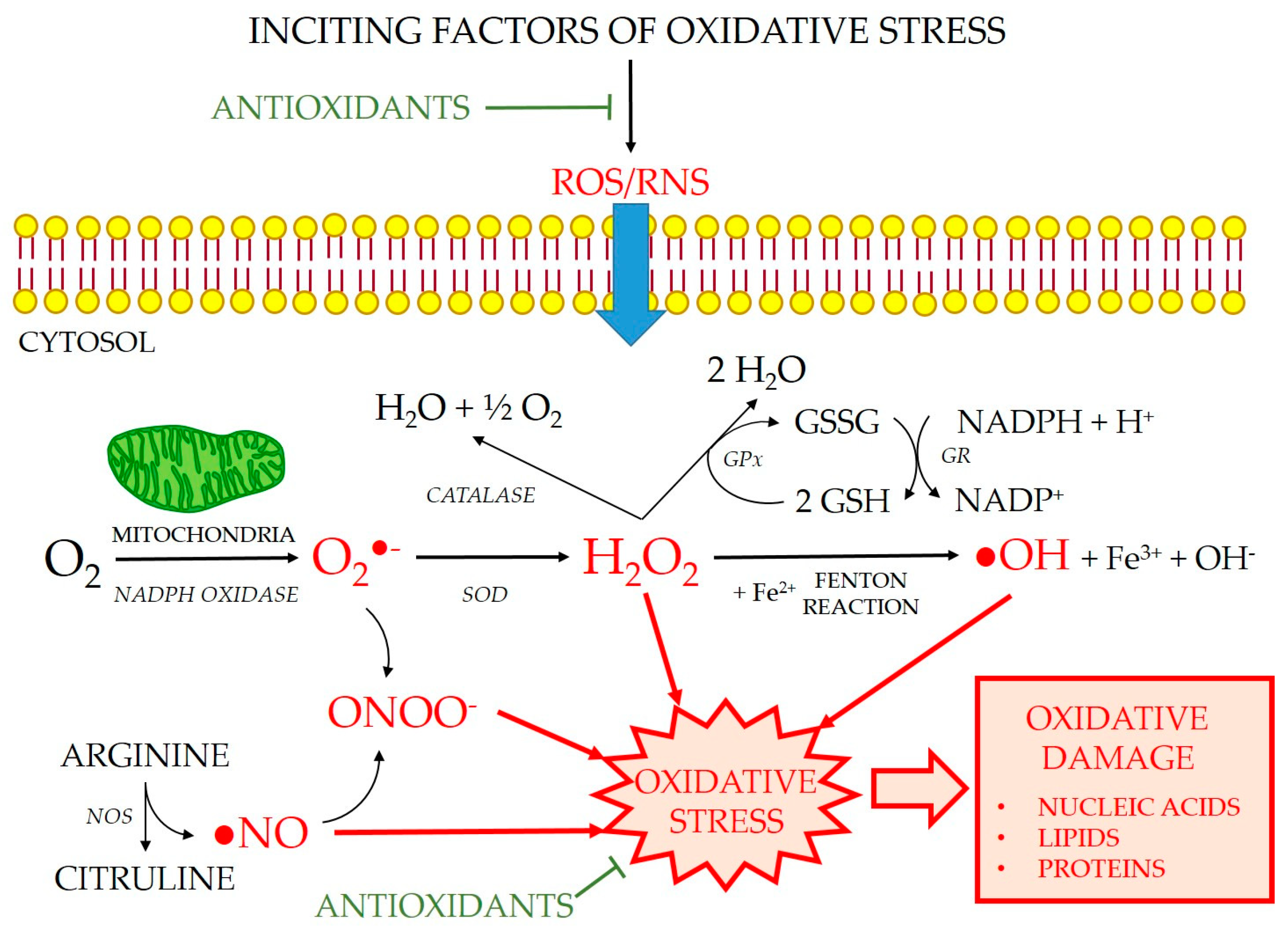
3. Oxidative Stress in the Interstitial Cells of Cajal and the Enteric Nervous System
4. Gastrointestinal Disorders Involving ICCs and the ENS
4.1. Hirschsprung’s Disease
4.2. Achalasia
4.3. Hypertrophic Pyloric Stenosis
4.4. Gastroesophageal Reflux
4.5. Inflammatory Bowel Disease
4.6. Chagas Disease
4.7. Gastrointestinal Stromal Tumors
4.8. Gastroparesia
4.9. Post-Operative Paralytic Ileus
4.10. Constipation
5. Neurodegenerative Diseases with Involvement of the ENS
5.1. Parkinson’s Disease
5.2. Alzheimer’s Disease
5.3. Amyotrophic Lateral Sclerosis
6. Neurodevelopmental Disorders with Involvement of the ENS
6.1. Autism Spectrum Disorders
6.2. Attention-Deficit/Hyperactivity Disorders
6.3. Down Syndrome
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rao, M.; Michael, D. The bowel and beyond: The enteric nervous system in neurological disorders. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 517–528. [Google Scholar] [CrossRef]
- Szylberg, L.; Marszalek, A. Diagnosis of Hirschsprung’s disease with particular emphasis on histopathology. A systematic review of current literature. Gastroenterol. Rev. 2014, 9, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Wright, C.M.; Heuckeroth, R.O. Unexpected Roles for the Second Brain: Enteric Nervous System as Master Regulator of Bowel Function. Annu. Rev. Physiol. 2019, 81, 235–259. [Google Scholar] [CrossRef] [PubMed]
- Avetisyan, M.; Schill, E.M.; Heuckeroth, R.O. Building a second brain in the bowel. J. Clin. Investig. 2015, 125, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Gershon, M.D. Enteric nervous system development: What could possibly go wrong? Nat. Rev. Neurosci. 2018, 19, 552–565. [Google Scholar] [CrossRef]
- Schemann, M.; Frieling, T.; Enck, P. To learn, to remember, to forget—How smart is the gut? Acta Physiol. 2019, 228, e13296. [Google Scholar] [CrossRef] [PubMed]
- Neuhuber, W.; Wörl, J. Monoamines in the enteric nervous system. Histochem. Cell Biol. 2018, 150, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, P.P.; Bertrand, R.L. Serotonin release and uptake in the gastrointestinal tract. Auton. Neurosci. Basic Clin. 2010, 153, 47–57. [Google Scholar] [CrossRef]
- Kaji, N.; Nakayama, S.; Horiguchi, K.; Iino, S.; Ozaki, H.; Hori, M. Disruption of the pacemaker activity of interstitial cells of Cajal via nitric oxide contributes to postoperative ileus. Neurogastroenterol. Motil. 2018, 30, e13334. [Google Scholar] [CrossRef]
- Mikkelsen, H.B. Interstitial cells of Cajal, macrophages and mast cells in the gut musculature: Morphology, distribution, spatial and possible functional interactions. J. Cell Mol. Med. 2010, 14, 818–832. [Google Scholar] [CrossRef]
- Blair, P.J.; Rhee, P.L.; Sanders, K.M.; Ward, S.M. The significance of interstitial cells in neurogastroenterology. J. Neurogastroenterol. Motil 2014, 20, 294–317. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lopez, P.; Garcia-Marin, V.; Freire, M. The histological slides and drawings of Cajal. Front. Neuroanat. 2010, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Radenkovic, G.; Radenkovic, D.; Velickov, A. Development of interstitial cells of Cajal in the human digestive tract as the result of reciprocal induction of mesenchymal and neural crest cells. J. Cell Mol. Med. 2018, 22, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Workman, M.J.; Mahe, M.M.; Trisno, S.; Poling, H.M.; Carey, L.; Watson, C.L.; Sundaram, N.; Chang, C.F.; Schiesser, J.; Aubert, P.; et al. Engineered human pluripotent-stem-cell-derived intestinal tissues with a functional enteric nervous system. Nat. Med. 2017, 23, 49–59. [Google Scholar] [CrossRef]
- Al-Sajee, D.; Huizinga, J.D. Interstitial cells of Cajal: Pathology, injury and repair. Sultan Qaboos Univ. Med. J. 2012, 12, 411–421. [Google Scholar] [CrossRef]
- Gfroerer, S.; Rolle, U. Interstitial cells of Cajal in the normal human gut and in Hirschsprung disease. Pediatr. Surg. Int. 2013, 29, 889–897. [Google Scholar] [CrossRef]
- Garcia-Lopez, P.; Garcia-Marin, V.; Martínez-Murillo, R.; Freire, M. Updating old ideas and recent advances regarding the Interstitial Cells of Cajal. Brain Res. Rev. 2009, 61, 154–169. [Google Scholar] [CrossRef]
- Sohal, G.S.; Ali, M.M.; Farooqui, F.A. A second source of precursor cells for the developing enteric nervous system and interstitial cells of Cajal. Int. J. Dev. Neurosci. 2002, 20, 619–626. [Google Scholar] [CrossRef]
- Al-Shboul, O. The importance of interstitial cells of cajal in the gastrointestinal tract. Saudi J. Gastroenterol. 2013, 19, 3–15. [Google Scholar] [CrossRef]
- Streutker, C.J.; Huizinga, J.D.; Driman, D.K.; Riddell, R.H. Interstitial cells of Cajal in health and disease. Part I: Normal ICC structure and function with associated motility disorders. Histopathology 2007, 50, 176–189. [Google Scholar] [CrossRef]
- Pasternak, A.; Szura, M.; Gil, K.; Matyja, A. Interstitial cells of Cajal-systematic review. Folia Morphol. 2016, 75, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, R.M.; Moustafa, Y.M.; Hamdy, H. Interstitial cells of Cajal, the Maestro in health and disease. World J. Gastroenterol. 2010, 16, 3239–3248. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.I. Human Physiology, 12th ed.; McGraw Hill: New York, NY, USA, 2011. [Google Scholar]
- Komuro, T. Comparative morphology of interstitial cells of Cajal: Ultrastructural characterization. Microsc. Res. Tech. 1999, 47, 267–285. [Google Scholar] [CrossRef]
- Wang, L.; Vargas, H.; French, S.W. Cellular origin of gastrointestinal stromal tumors: A study of 27 cases. Arch. Pathol. Lab. Med. 2000, 124, 1471–1475. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Traini, C. Interstitial cells of Cajal and telocytes in the gut: Twins, related or simply neighbor cells? Biomol. Concepts 2016, 7, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D.; Martz, S.; Gil, V.; Wang, X.Y.; Jimenez, M.; Parsons, S. Two independent networks of interstitial cells of Cajal work cooperatively with the enteric nervous system to create colonic motor patterns. Front. Neurosci. 2011, 5, 93. [Google Scholar] [CrossRef]
- McCann, C.J.; Hwang, S.J.; Bayguinov, Y.; Colletti, E.J.; Sanders, K.M.; Ward, S.M. Establishment of pacemaker activity in tissues allotransplanted with interstitial cells of Cajal. Neurogastroenterol. Motil. 2013, 25, e418–e428. [Google Scholar] [CrossRef]
- Piotrowska, A.P.; Solari, V.; Puri, P. Distribution of heme oxygenase-2 in nerves and interstitial cells of Cajal in the normal pylorus and in infantile hypertrophic pyloric stenosis. Arch. Pathol. Lab. Med. 2003, 127, 1182–1186. [Google Scholar] [CrossRef]
- Kraichely, R.E.; Farrugia, G. Mechanosensitive ion channels in interstitial cells of Cajal and smooth muscle of the gastrointestinal tract. Neurogastroenterol. Motil. 2007, 19, 245–252. [Google Scholar] [CrossRef]
- Powley, T.L.; Wang, X.Y.; Fox, E.A.; Phillips, R.J.; Liu, L.W.C.; Huizinga, J.D. Ultrastructural evidence for communication between intramuscular vagal mechanoreceptors and interstitial cells of Cajal in the rat fundus. Neurogastroenterol. Motil. 2008, 20, 69–79. [Google Scholar] [CrossRef]
- Hassan, H.M.; Fridovich, I. Intracellular production of superoxide radical and of hydrogen peroxide by redox active compounds. Arch. Biochem. Biophys 1979, 196, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I. Antioxidant actions of plant foods: Use of oxidative DNA damage as a tool for studying antioxidant efficacy. Free Radic. Res. 1999, 30, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Young, I.S. Measurement of total antioxidant capacity. J. Clin. Pathol. 2001, 54, 339. [Google Scholar] [CrossRef] [PubMed]
- Miquel, J.; Fleming, J.E. Theoretical and experimental support for an oxygen radical-mitochondrial damage hypothesis of cell aging. In Free Radicals, Aging and Degenerative Diseases; Johnson, J.E., Jr., Harman, D., Walford, R., Miquel, J., Eds.; Alan R. Liss: New York, NY, USA, 1986; pp. 51–74. [Google Scholar]
- Yu, B.P. Cellular defenses against damage from reactive oxygen species. Physiol. Rev. 1994, 74, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Beckman, K.B.; Ames, B.N. The free radical theory of aging matures. Physiol. Rev. 1998, 78, 547–581. [Google Scholar] [CrossRef]
- Mocellin, S.; Bronte, V.; Nitti, D. Nitric oxide, a double edged sword in cancer biology: Searching for therapeutic opportunities. Med. Res. Rev. 2007, 27, 317–352. [Google Scholar] [CrossRef]
- Costa, N.T.; Iriyoda, T.M.V.; Alfieri, D.F.; Simão, A.N.C.; Dichi, I. Influence of disease-modifying antirheumatic drugs on oxidative and nitrosative stress in patients with rheumatoid arthritis. Inflammopharmacology 2018, 26, 1151–1164. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants: A personal view. Nutr. Rev. 1994, 52 Pt 1, 253–265. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef]
- Reiter, R.J. The role of the neurohormone melatonin as a buffer against macromolecular oxidative damage. Neurochem. Int. 1995, 27, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Mehlhorn, R. Mutagenicity of nitroxide-free radicals. Arch. Biochem. Biophys 1986, 251, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A. Oxidative stress, oxidant defense, and dietary constituents. Modern nutrition. In Health and Disease, 8th ed.; Shils, M.E., Olson, J.A., Shike, M., Eds.; Williams and Wilkins: Philadelphia, PA, USA, 1994; pp. 501–512. [Google Scholar]
- Cheeseman, K.H.; Slater, T.F. An introduction to free radical biochemistry. Br. Med. Bull. 1993, 49, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, J.M. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin. Chem. 1995, 41 Pt 2, 1819–1828. [Google Scholar] [CrossRef]
- Gaweł, S.; Wardas, M.; Niedworok, E.; Wardas, P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad Lek. 2004, 57, 453–455. [Google Scholar]
- Dumitrescu, L.; Popescu-Olaru, I.; Cozma, L.; Tulbǎ, D.; Hinescu, M.E.; Ceafalan, L.C.; Gherghiceanu, M.; Popescu, B.O. Oxidative stress and the microbiota-gut-brain axis. Oxid Med. Cell Longev. 2018, 2018, 2406594. [Google Scholar] [CrossRef]
- Takahashi, T. Pathophysiological significance of neuronal nitric oxide synthase in the gastrointestinal tract. J. Gastroenterol. 2003, 38, 421–430. [Google Scholar] [CrossRef]
- Sanders, K.M.; Ward, S.M. Nitric oxide and its role as a non-adrenergic, non-cholinergic inhibitory neurotransmitter in the gastrointestinal tract. Br. J. Pharmacol. 2019, 176, 212–227. [Google Scholar] [CrossRef]
- Friebe, A.; Voußen, B.; Groneberg, D. NO-GC in cells off the beaten track. Nitric. Oxide—Biol. Chem. 2018, 77, 12–18. [Google Scholar] [CrossRef]
- Kaji, N.; Horiguchi, K.; Iino, S.; Nakayama, S.; Ohwada, T.; Otani, Y.; Firman, M.; Sanders, K.; Ozaki, H.; Hori, M. Nitric oxide-induced oxidative stress impairs pacemaker function of murine interstitial cells of Cajal during inflammation. Pharmacol. Res. 2016, 111, 838–848. [Google Scholar] [CrossRef]
- Loffredo, L.; Ettorre, E.; Zicari, A.M.; Inghilleri, M.; Nocella, C.; Perri, L.; Spalice, A.; Fossati, C.; De Lucia, M.C.; Pigozzi, F.; et al. Oxidative Stress and Gut-Derived Lipopolysaccharides in Neurodegenerative Disease: Role of NOX2. Oxid Med. Cell Longev. 2020, 3, 8630275. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kong, P.; Chen, C.; Tang, J.; Jin, X.; Yan, J.; Wang, Y. Targeting IL-17A Improves the Dysmotility of the Small Intestine and Alleviates the Injury of the Interstitial Cells of Cajal during Sepsis. Oxid Med. Cell Longev. 2019, 2019, 1475729. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Pollock, J.; Schmidt, H.H.H.W.; Ward, S.M.; Sanders, K.M. Expression of nitric oxide synthase immunoreactivity by interstitial cells of the canine proximal colon. J. Auton. Nerv. Syst. 1994, 49, 1–14. [Google Scholar] [CrossRef]
- Rivera, L.R.; Poole, D.P.; Thacker, M.; Furness, J.B. The involvement of nitric oxide synthase neurons in enteric neuropathies. Neurogastroenterol. Motil. 2011, 23, 980–988. [Google Scholar] [CrossRef]
- Gangula, P.R.R.; Maner, W.L.; Micci, M.A.; Garfield, R.E.; Pasricha, P.J. Diabetes induces sex-dependent changes in neuronal nitric oxide synthase dimerization and function in the rat gastric antrum. Am. J. Physiol.—Gastrointest. Liver Physiol. 2007, 292, G725–G733. [Google Scholar] [CrossRef]
- Bódi, N.; Szalai, Z.; Bagyánszki, M. Nitrergic enteric neurons in health and disease-focus on animal models. Int. J. Mol. Sci. 2019, 20, 2003. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Seidler, B.; Kettenberger, A.; Sibaev, A.; Rohn, M.; Feil, R.; Allescher, H.D.; Vanderwinden, J.M.; Hofmann, F.; Schemann, M.; et al. Interstitial cells of Cajal integrate excitatory and inhibitory neurotransmission with intestinal slow-wave activity. Nat. Commun. 2013, 4, 1630. [Google Scholar] [CrossRef]
- Iino, S.; Horiguchi, K.; Nojyo, Y. Interstitial cells of Cajal are innervated by nitrergic nerves and express nitric oxide-sensitive guanylate cyclase in the guinea-pig gastrointestinal tract. Neuroscience 2008, 152, 437–448. [Google Scholar] [CrossRef]
- Choi, K.M.; Gibbons, S.J.; Nguyen, T.V.; Stoltz, G.J.; Lurken, M.S.; Ordog, T.; Szurszewski, J.H.; Farrugia, G. Heme oxygenase-1 protects interstitial cells of Cajal from oxidative stress and reverses diabetic gastroparesis. Gastroenterology 2008, 135, 2055–2064.e2. [Google Scholar] [CrossRef]
- Orozco-Ibarra, M.; Pedraza-Chaverrí, J. Hemo oxigenasa: Aspectos básicos y su importancia en el sistema nervioso central. Arch. Neurociencias 2010, 15, 47–55. [Google Scholar]
- Doré, S.; Takahashi, M.; Ferris, C.D.; Hester, L.D.; Guastella, D.; Snyder, S.H. Bilirubin, formed by activation of heme oxygenase-2, protects neurons against oxidative stress injury. Proc. Natl. Acad. Sci. USA 1999, 96, 2445–2450. [Google Scholar] [CrossRef] [PubMed]
- Colpaert, E.E.; Timmermans, J.P.; Lefebvre, R.A. Immunohistochemical localization of the antioxidant enzymes biliverdin reductase and heme oxygenase-2 in human and pig gastric fundus. Free Radic. Biol. Med. 2002, 32, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Zhang, Z.; Miao, Y.; Xu, Y.; Zhu, J.; Wan, Q. Physiological serum total bilirubin concentrations were inversely associated with diabetic peripheral neuropathy in Chinese patients with type 2 diabetes: A cross-sectional study. Diabetol. Metab. Syndr. 2019, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.; Di Domenico, F.; Mancuso, C.; Butterfield, D.A. The Janus Face of the Heme Oxygenase/Biliverdin Reductase System in Alzheimer Disease: It’s Time for Reconciliation. Neurobiol. Dis. 2014, 41, 2971–3010. [Google Scholar] [CrossRef] [PubMed]
- Nitti, M.; Piras, S.; Brondolo, L.; Marinari, U.M.; Pronzato, M.A.; Furfaro, A.L. Heme oxygenase 1 in the nervous system: Does it favor neuronal cell survival or induce neurodegeneration? Int. J. Mol. Sci. 2018, 19, 2260. [Google Scholar] [CrossRef]
- Schipper, H.M.; Song, W.; Tavitian, A.; Cressatti, M. The sinister face of heme oxygenase-1 in brain aging and disease. Prog. Neurobiol. 2019, 172, 40–70. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide dismutases: Role in redox signaling, vascular function, and diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- Patlevič, P.; Vašková, J.; Švorc, P.; Vaško, L.; Švorc, P. Reactive oxygen species and antioxidant defense in human gastrointestinal diseases. Integr. Med. Res. 2016, 5, 250–258. [Google Scholar] [CrossRef]
- Ishihara, T.; Tanaka, K.I.; Tasaka, Y.; Namba, T.; Suzuki, J.; Ishihara, T.; Okamoto, S.; Hibi, T.; Takenaga, M.; Igarashi, R.; et al. Therapeutic effect of lecithinized superoxide dismutase against colitis. J. Pharmacol. Exp. Ther. 2009, 328, 152–164. [Google Scholar] [CrossRef]
- Horspool, A.M.; Chang, H.C. Neuron-specific regulation of superoxide dismutase amid pathogen-induced gut dysbiosis. Redox Biol. 2018, 17, 377–385. [Google Scholar] [CrossRef]
- Fang, S.; Christensen, J. Manganese superoxide dismutase and reduced nicotinamide adenine dinucleotide diaphorase colocalize in the rat gut. Gastroenterology 1995, 109, 1429–1436. [Google Scholar] [CrossRef]
- Puri, P.; Ohshiro, K.; Wester, T. Hirschsprung’s disease: A search for etiology. Pediatr. Surg. 1998, 7, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Matsuura, K.; Matsuoka, T.; Nakatani, H.; Nakano, T.; Furuya, Y.; Sugimoto, T.; Kobayashi, M.; Araki, K. A morphological study of the pacemaker cells of the aganglionic intestine in Hirschsprung’s disease utilizing ls/ls model mice. Med. Mol. Morphol. 2005, 38, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, L.; Maddur, S.; Puri, P. Immunolocalization of the Gap junction protein Connexin43 in the interstitial cells of Cajal in the normal and Hirschsprung’s disease bowel. J. Pediatr. Surg. 2000, 35, 823–828. [Google Scholar] [CrossRef]
- Furness, J.B.; Callaghan, B.P.; Rivera, L.R.; Cho, H.J. The Enteric Nervous System and Gastrointestinal Innervation: Integrated Local and Central Control. Adv. Exp. Med. Biol. 2014, 817, 115–133. [Google Scholar] [CrossRef]
- Margolis, K.G.; Karamooz, N.; Stevanovic, K.; Li, Z.S.; Ahuja, A.; D’Autréaux, F.; Saurman, V.; Chalazonitis, A.; Gershon, M.D. Enteric Neuronal Density Contributes to the Severity of Intestinal Inflammation. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [PubMed]
- De Lima, M.A.; Cabrine-Santos, M.; Tavares, M.G.; Gerolin, G.P.; Lages-Silva, E.; Ramirez, L.E. Interstitial cells of Cajal in chagasic megaesophagus. Ann. Diagn Pathol. 2008, 12, 271–274. [Google Scholar] [CrossRef]
- Freitas, M.A.R.; Segatto, N.; Tischler, N.; de Oliveira, E.C.; Brehmer, A.; da Silveira, A.B.M. Relation between mast cells concentration and serotonin expression in chagasic megacolon development. Parasite Immunol. 2017, 39, 19–22. [Google Scholar] [CrossRef]
- Nakajima, T.; Miwa, S.; Ando, T.; Fujinami, H.; Kajiura, S.; Hosokawa, A.; Takano, Y.; Sugiyama, T. Interstitial cells of Cajal do not harbor c-kit or PDGFRA gene mutations in patients with sporadic gastrointestinal stromal tumors. J. Gastroenterol. 2009, 44, 426–431. [Google Scholar] [CrossRef]
- The, F.O.; Cailotto, C.; Van Der Vliet, J.; De Jonge, W.J.; Bennink, R.J.; Buijs, R.M.; Boeckxstaens, G.E. Central activation of the cholinergic anti-inflammatory pathway reduces surgical inflammation in experimental post-operative ileus. Br. J. Pharmacol. 2011, 163, 1007–1016. [Google Scholar] [CrossRef]
- Camilleri, M.; Cowen, T.; Koch, T.R. Enteric neurodegeneration in ageing. Neurogastroenterol. Motil 2008, 20, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Bhutto, A.; Morley, J.E. The clinical significance of gastrointestinal changes with aging. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pinilla, P.J.; Gibbons, S.J.; Sarr, M.G.; Kendrick, M.L.; Shen, K.R.; Cima, R.R.; Dozois, E.J.; Larson, D.W.; Ordog, T.; Pozo, M.J.; et al. Changes in interstitial cells of cajal with age in the human stomach and colon. Neurogastroenterol. Motil. 2011, 23, 36–44. [Google Scholar] [CrossRef]
- Farrugia, G. Interstitial cells of Cajal in health and disease. Neurogastroenterol. Motil. 2008, 20, 54–63. [Google Scholar] [CrossRef]
- Currò, D.; Ipavec, V.; Preziosi, P. Neurotransmitters of the non-adrenergic non-cholinergic relaxation of proximal stomach. Eur. Rev. Med. Pharmacol. Sci. 2008, 12, 53–62. [Google Scholar] [PubMed]
- Leclair-Visonneau, L.; Neunlist, M.; Derkinderen, P.; Lebouvier, T. The gut in Parkinson’s disease: Bottom-up, top-down, or neither? Neurogastroenterol. Motil. 2020, 32, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging. Cell 2019, 18, e13031. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging. 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Rietdijk, C.D.; Perez-Pardo, P.; Garssen, J.; van Wezel, R.J.A.; Kraneveld, A.D. Exploring Braak’s hypothesis of parkinson’s disease. Front. Neurol. 2017, 8, 37. [Google Scholar] [CrossRef]
- Lionnet, A.; Leclair-Visonneau, L.; Neunlist, M.; Murayama, S.; Takao, M.; Adler, C.H.; Derkinderen, P.; Beach, T.G. Does Parkinson’s disease start in the gut? Acta Neuropathol. 2017, 135, 1469–1480. [Google Scholar] [CrossRef]
- Svensson, E.; Horváth-Puhó, E.; Thomsen, R.W.; Djurhuus, J.C.; Pedersen, L.; Borghammer, P.; Sørensen, H.T. Vagotomy and subsequent risk of Parkinson’s disease. Ann. Neurol. 2015, 78, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kurek, J.; Morgan, J.C.; Wakade, C.; Rao, S.S.C. Constipation in Parkinson’s Disease: A Nuisance or Nuanced Answer to the Pathophysiological Puzzle? Curr. Gastroenterol. Rep. 2018, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Colucci, R.; Antonioli, L.; Barocelli, E.; Ballabeni, V.; Bernardini, N.; Blandizzi, C.; De Jonge, W.J.; Fornai, M. Intestinal dysfunction in Parkinson’s disease: Lessons learned from translational studies and experimental models. Neurogastroenterol. Motil. 2016, 28, 1781–1791. [Google Scholar] [CrossRef]
- Colucci, M.; Cervio, M.; Faniglione, M.; De Angelis, S.; Pajoro, M.; Levandis, G.; Tassorelli, C.; Blandini, F.; Feletti, F.; De Giorgio, R.; et al. Intestinal dysmotility and enteric neurochemical changes in a Parkinson’s disease rat model. Auton. Neurosci. Basic. Clin. 2012, 169, 77–86. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K. Neuropathological Staging of Brain Pathology in Sporadic Parkinson’s disease: Separating the Wheat from the Chaff. J. Park. Dis. 2017, 7, S73–S87. [Google Scholar] [CrossRef]
- Annerino, D.M.; Arshad, S.; Taylor, G.M.; Adler, C.H.; Beach, T.G. Parkinson’s disease is not associated with gastrointestinal myenteric ganglion neuron loss. Acta Neuropathol. 2012, 124, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Corbillé, A.G.; Coron, E.; Neunlist, M.; Derkinderen, P.; Lebouvier, T. Appraisal of the dopaminergic and noradrenergic innervation of the submucosal plexus in PD. J. Park. Dis. 2014, 4, 571–576. [Google Scholar] [CrossRef]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef]
- Semar, S.; Klotz, M.; Letiembre, M.; Van Ginneken, C.; Braun, A.; Jost, V.; Bischof, M.; Lammers, W.J.; Liu, Y.; Fassbender, K.; et al. Changes of the enteric nervous system in amyloid-β protein precursor transgenic mice correlate with disease progression. J. Alzheimer’s Dis. 2013, 36, 7–20. [Google Scholar] [CrossRef]
- Van Ginneken, C.; Schäfer, K.H.; Van Dam, D.; Huygelen, V.; De Deyn, P.P. Morphological changes in the enteric nervous system of aging and APP23 transgenic mice. Brain Res. 2011, 1378, 43–53. [Google Scholar] [CrossRef]
- Puig, K.L.; Lutz, B.M.; Urquhart, S.A.; Rebel, A.; Zhou, X.; Manocha, G.D.; Sens, M.A.; Tuteja, A.K.; Foster, N.L.; Coombs, C.K. Overexpression of Mutant Amyloid-β Protein Precursor and Presenilin 1 Modulates Enteric Nervous System. J. Alzheimer’s Dis. 2015, 44, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Ragagnin, A.M.G.; Shadfar, S.; Vidal, M.; Jamali, M.S.; Atkin, J.D. Motor neuron susceptibility in ALS/FTD. Front. Neurosci. 2019, 13, 532. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Antonioli, L.; Colucci, R.; Blandizzi, C.; Fornai, M. Interplay among gut microbiota, intestinal mucosal barrier and enteric neuro-immune system: A common path to neurodegenerative diseases? Acta Neuropathol. 2018, 136, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Wegorzewska, I.; Bell, S.; Cairns, N.J.; Miller, T.M.; Baloh, R.H. TDP-43 mutant transgenic mice develop features of ALS and frontotemporal lobar degeneration. Proc. Natl. Acad. Sci. USA 2009, 106, 18809–18814. [Google Scholar] [CrossRef]
- Jo, M.; Lee, S.; Jeon, Y.M.; Kim, S.; Kwon, Y.; Kim, H.J. The role of TDP-43 propagation in neurodegenerative diseases: Integrating insights from clinical and experimental studies. Exp. Mol. Med. 2020, 52, 1652–1662. [Google Scholar] [CrossRef]
- Natale, G.; Pasquali, L.; Paparelli, A.; Fornai, F. Parallel manifestations of neuropathologies in the enteric and central nervous systems. Neurogastroenterol. Motil. 2011, 23, 1056–1065. [Google Scholar] [CrossRef]
- Ekhtiari-Bidhendi, E.; Bergh, J.; Zetterström, P.; Forsberg, K.; Pakkenberg, B.; Andersen, P.M.; Marklund, S.L.; Brännström, T. Mutant superoxide dismutase aggregates from human spinal cord transmit amyotrophic lateral sclerosis. Acta Neuropathol. 2018, 136, 939–953. [Google Scholar] [CrossRef]
- Carrera-Juliá, S.; Moreno, M.L.; Barrios, C.; de la Rubia Ortí, J.E.; Drehmer, E. Antioxidant Alternatives in the Treatment of Amyotrophic Lateral Sclerosis: A Comprehensive Review. Front. Physiol. 2020, 11, 63. [Google Scholar] [CrossRef]
- Fuentealba, R.A.; Udan, M.; Bell, S.; Wegorzewska, I.; Shao, J.; Diamond, M.I.; Weihl, C.C.; Baloh, R.H. Interaction with polyglutamine aggregates reveals a Q/N-rich domain in TDP-43. J. Biol. Chem. 2010, 285, 26304–26314. [Google Scholar] [CrossRef]
- Marrie, R.A.; Reider, N.; Stuve, O.; Trojano, M.; Sorensen, P.S.; Cutter, G.R.; Reingold, S.C.; Cohen, J. The incidence and prevalence of comorbid gastrointestinal, musculoskeletal, ocular, pulmonary, and renal disorders in multiple sclerosis: A systematic review. Mult. Scler. 2015, 21, 332–341. [Google Scholar] [CrossRef]
- McElhanon, B.O.; McCracken, C.; Karpen, S.; Sharp, W.G. Gastrointestinal symptoms in autism spectrum disorder: A meta-analysis. Pediatrics 2014, 133, 872–883. [Google Scholar] [CrossRef] [PubMed]
- Marler, S.; Ferguson, B.J.; Lee, E.B.; Peters, B.; Williams, K.C.; Mcdonnell, E.; Macklin, E.A.; Levitt, P.; Gillespie, C.H.; Anderson, G.M.; et al. Brief Report: Whole Blood Serotonin Levels and Gastrointestinal Symptoms in Autism Spectrum Disorder. J. Autism. Dev. Disord. 2016, 46, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Hosie, S.; Ellis, M.; Swaminathan, M.; Ramalhosa, F.; Seger, G.O.; Balasuriya, G.K.; Gillberg, C.; Rastam, M.; Churilov, L.; McKeown, S.J.; et al. Gastrointestinal dysfunction in patients and mice expressing the autism-associated R451C mutation in neuroligin-3. Autism. Res. 2019, 12, 1043–1056. [Google Scholar] [CrossRef]
- Sharna, S.S.; Balasuriya, G.K.; Hosie, S.; Nithianantharajah, J.; Franks, A.E.; Hill-Yardin, E.L. Altered Caecal Neuroimmune Interactions in the Neuroligin-3R451C Mouse Model of Autism. Front. Cell Neurosci. 2020, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Sandgren, A.M.; Brummer, R.J.M. ADHD-originating in the gut? The emergence of a new explanatory model. Med. Hypotheses 2018, 120, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Eltokhi, A.; Janmaat, I.E.; Genedi, M.; Haarman, B.C.M.; Sommer, I.E.C. Dysregulation of synaptic pruning as a possible link between intestinal microbiota dysbiosis and neuropsychiatric disorders. J. Neurosci. Res. 2020, 98, 1335–1369. [Google Scholar] [CrossRef]
- Moore, S.W. Hirschsprung’s disease and the brain. Pediatr. Surg. Int. 2011, 27, 347–352. [Google Scholar] [CrossRef]
- Moore, S.W. Advances in understanding the association between Down syndrome and Hirschsprung disease (DS-HSCR). Pediatr. Surg. Int. 2018, 34, 1127–1137. [Google Scholar] [CrossRef]
- Becker, L.E.; Armstrong, D.L.; Chan, F. Dendritic atrophy in children with Down’s syndrome. Ann. Neurol. 1986, 20, 520–526. [Google Scholar] [CrossRef]
- Nakazato, Y.; Landing, B.H. Reduced number of neurons in esophageal plexus ganglia in Down syndrome: Additional evidence for reduced cell number as a basic feature of the disorder. Pediatr. Pathol. 1986, 5, 55–63. [Google Scholar] [CrossRef]
- Schill, E.M.; Wright, C.M.; Jamil, A.; LaCombe, J.M.; Roper, R.J.; Heuckeroth, R.O. Down syndrome mouse models have an abnormal enteric nervous system. JCI Insight. 2019, 5, e124510. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Pingarrón, L.; Almeida, H.; Soria-Aznar, M.; Reyes-Gonzales, M.C.; Rodríguez-Moratinos, A.B.; Muñoz-Hoyos, A.; García, J.J. Interstitial Cells of Cajal and Enteric Nervous System in Gastrointestinal and Neurological Pathology, Relation to Oxidative Stress. Curr. Issues Mol. Biol. 2023, 45, 3552-3572. https://doi.org/10.3390/cimb45040232
López-Pingarrón L, Almeida H, Soria-Aznar M, Reyes-Gonzales MC, Rodríguez-Moratinos AB, Muñoz-Hoyos A, García JJ. Interstitial Cells of Cajal and Enteric Nervous System in Gastrointestinal and Neurological Pathology, Relation to Oxidative Stress. Current Issues in Molecular Biology. 2023; 45(4):3552-3572. https://doi.org/10.3390/cimb45040232
Chicago/Turabian StyleLópez-Pingarrón, Laura, Henrique Almeida, Marisol Soria-Aznar, Marcos C. Reyes-Gonzales, Ana B. Rodríguez-Moratinos, Antonio Muñoz-Hoyos, and Joaquín J. García. 2023. "Interstitial Cells of Cajal and Enteric Nervous System in Gastrointestinal and Neurological Pathology, Relation to Oxidative Stress" Current Issues in Molecular Biology 45, no. 4: 3552-3572. https://doi.org/10.3390/cimb45040232