
Development of Digital Droplet PCR Targeting the Influenza H3N2 Oseltamivir-Resistant E119V Mutation and Its Performance through the Use of Reverse Genetics Mutants

 , , and
, , and 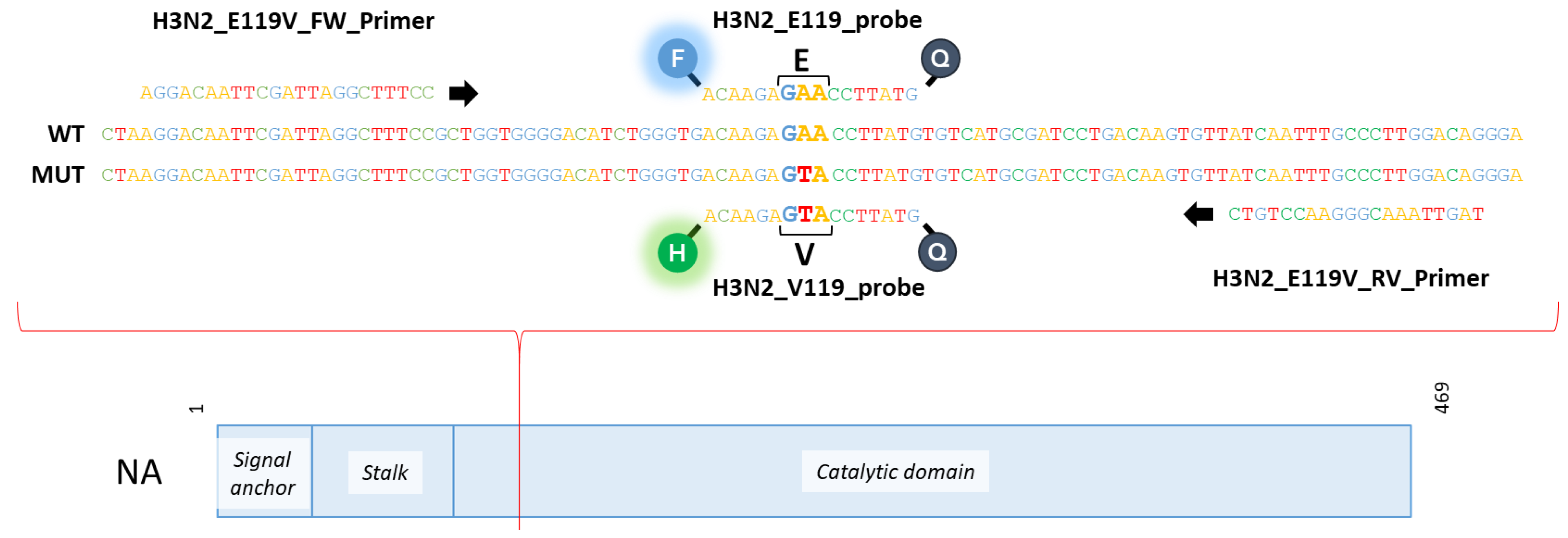{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses, Cells, and Chemicals
2.2. Mutagenesis and Reverse Genetics
2.3. Wild-Type and E119V Mutant Virus Mixes
2.4. NA Activity and Inhibition Assays
2.5. ddPCR Protocol

3. Results
3.1. Generation of the Wild Type and E119V-NA Mutant
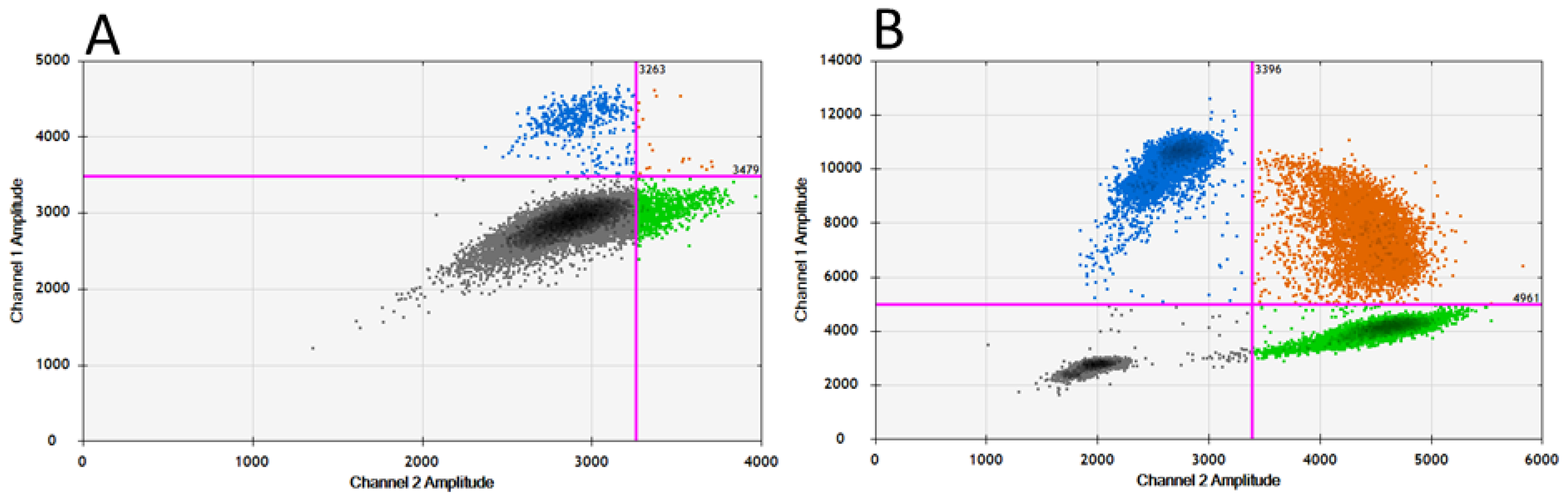
3.2. Adaptation of RT-qPCR to Obtain a Specific RT-ddPCR to Detect the Polymorphism at Position 119-NA of Influenza A(H3N2) Viruses
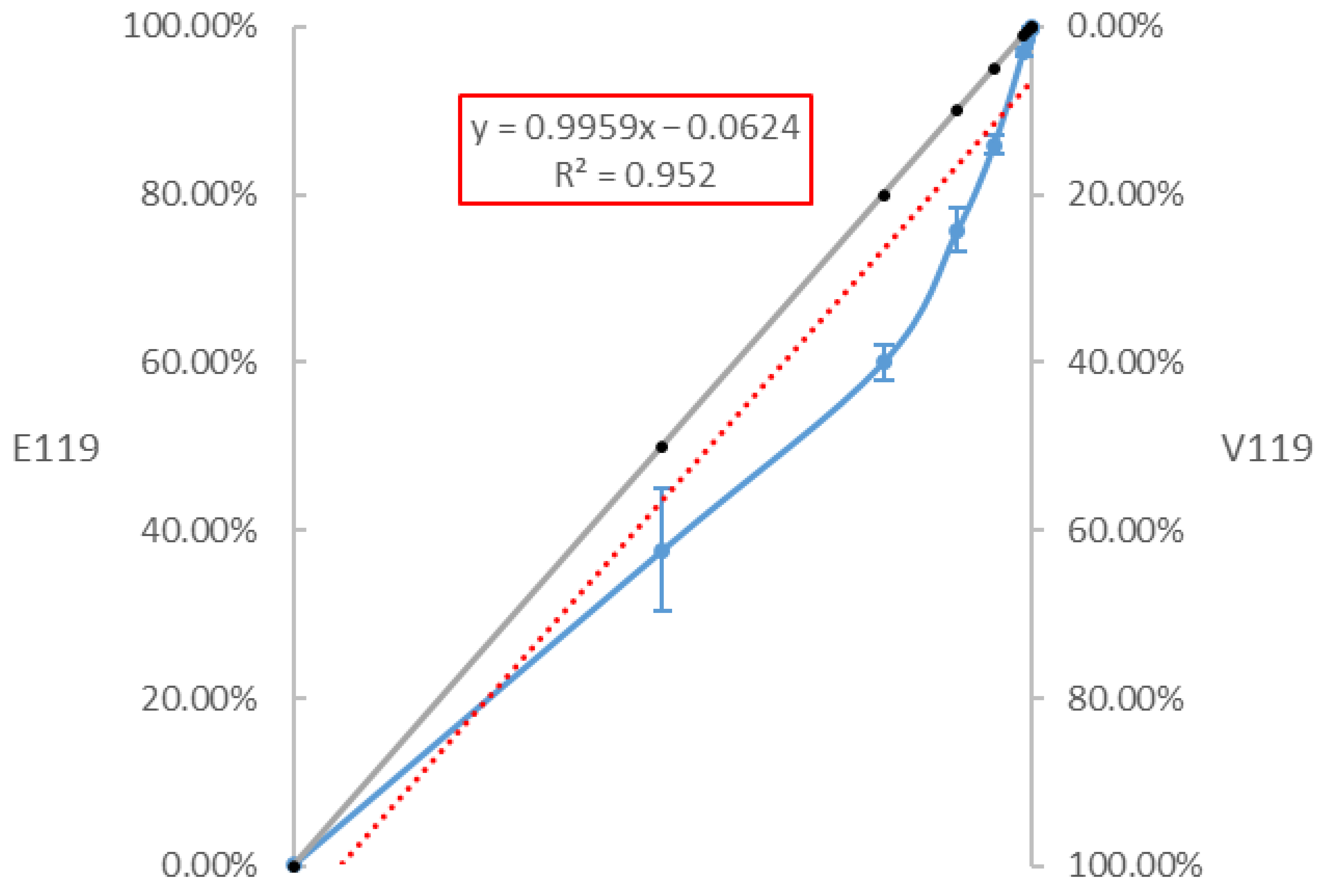
3.3. Sensitivity of the RT-ddPCR to Quantify the Proportion of the E119V-NA Mutation
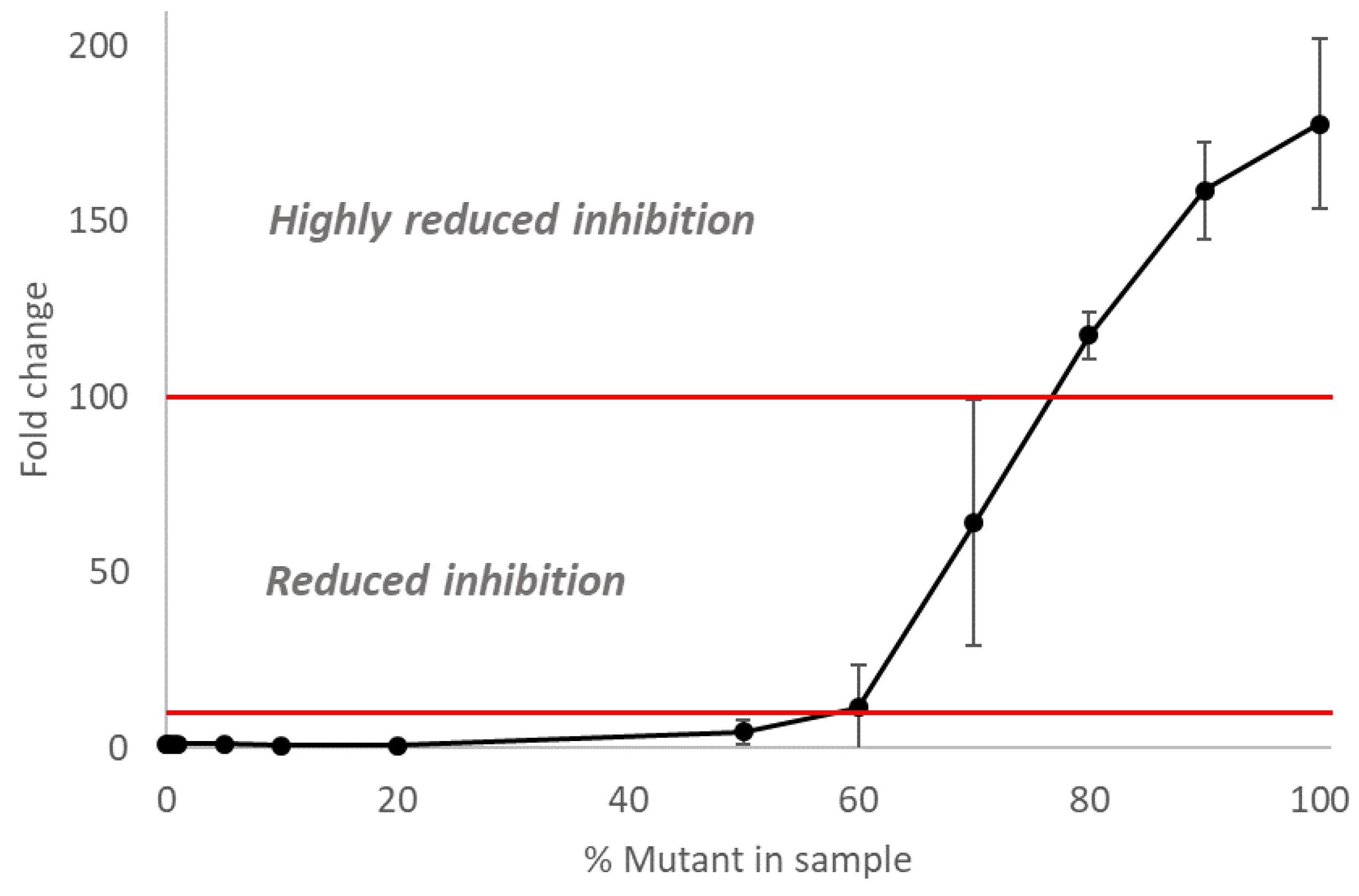
3.4. The Inhibitory Activity of Oseltamivir on Mixed Populations of NAI-Susceptible and -Resistant Influenza A(H3N2) Viruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Vries, E.; Anber, J.; Van Der Linden, A.; Wu, Y.; Maaskant, J.; Stadhouders, R.; Van Beek, R.; Rimmelzwaan, G.; Osterhaus, A.; Boucher, C.; et al. Molecular Assays for Quantitative and Qualitative Detection of Influenza Virus and Oseltamivir Resistance Mutations. J. Mol. Diagn. 2013, 15, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Van der Vries, E.; Schutten, M.; Fraaij, P.; Boucher, C.; Osterhaus, A. Influenza Virus Resistance to Antiviral Therapy. Adv. Pharmacol. 2013, 67, 217–246. [Google Scholar] [PubMed]
- World Health Organization. WHO Global Strategy for Containment of Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- Kosik, I.; Yewdell, J.W. Influenza Hemagglutinin and Neuraminidase: Yin–Yang Proteins Coevolving to Thwart Immunity. Viruses 2019, 11, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M. Evolution and Ecology of Influenza A Viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Mosnier, A.; Caini, S.; Daviaud, I.; Nauleau, E.; Bui, T.T.; Debost, E.; Bedouret, B.; Agius, G.; van der Werf, S.; Lina, B.; et al. Clinical Characteristics Are Similar across Type A and B Influenza Virus Infections. PLoS ONE 2015, 10, e0136186. [Google Scholar] [CrossRef]
- Monto, A.S.; McKimm-Breschkin, J.L.; Macken, C.; Hampson, A.W.; Hay, A.; Klimov, A.; Tashiro, M.; Webster, R.G.; Aymard, M.; Hayden, F.G.; et al. Detection of Influenza Viruses Resistant to Neuraminidase Inhibitors in Global Surveillance during the First 3 Years of Their Use. Antimicrob. Agents Chemother. 2006, 50, 2395–2402. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Chung, J.; Palese, P. Alterations of the Stalk of the Influenza Virus Neuraminidase: Deletions and Insertions. Virus Res. 1993, 29, 141–153. [Google Scholar] [CrossRef]
- Palese, P.; Tobita, K.; Ueda, M.; Compans, R.W. Characterization of Temperature Sensitive Influenza Virus Mutants Defective in Neuraminidase. Virology 1974, 61, 397–410. [Google Scholar] [CrossRef]
- Demicheli, V.; Jefferson, T.; Pietrantonj, C.; Ferroni, E.; Thorning, R.E.; Rivette, A. Vaccines for Preventing Influenza in the Elderly. Cochrane Database Syst. Rev. 2018, 2. [Google Scholar] [CrossRef] [Green Version]
- Aoki, F.Y.; Boivin, G.; Roberts, N. Influenza Virus Susceptibility and Resistance to Oseltamivir. Antivir. Ther. 2007, 12, 603–616. [Google Scholar] [CrossRef]
- Stephenson, I.; Democratis, J.; Lackenby, A.; McNally, T.; Smith, J.; Pareek, M.; Ellis, J.; Bermingham, A.; Nicholson, K.; Zambon, M. Neuraminidase Inhibitor Resistance after Oseltamivir Treatment of Acute Influenza A and B in Children. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2009, 48, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Kiso, M.; Mitamura, K.; Sakai-Tagawa, Y.; Shiraishi, K.; Kawakami, C.; Kimura, K.; Hayden, F.G.; Sugaya, N.; Kawaoka, Y. Resistant Influenza A Viruses in Children Treated with Oseltarnivir: Descriptive Study. Lancet 2004, 364, 759–765. [Google Scholar] [CrossRef]
- Baz, M.; Abed, Y.; McDonald, J.; Boivin, G. Characterization of Multidrug-Resistant Influenza A/H3N2 Viruses Shed during 1 Year by an Immunocompromised Child. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2006, 43, 1555–1561. [Google Scholar] [CrossRef]
- Tamura, D.; Sugaya, N.; Ozawa, M.; Takano, R.; Ichikawa, M.; Yamazaki, M.; Kawakami, C.; Shimizu, H.; Uehara, R.; Kiso, M.; et al. Frequency of Drug-Resistant Viruses and Virus Shedding in Pediatric Influenza Patients Treated with Neuraminidase Inhibitors. Clin. Infect. Dis. 2011, 52, 432–437. [Google Scholar] [CrossRef]
- Ison, M.G.; Gubareva, L.V.; Atmar, R.L.; Treanor, J.; Hayden, F.G. Recovery of Drug-Resistant Influenza Virus from Immunocompromised Patients: A Case Series. J. Infect. Dis. 2006, 193, 760–764. [Google Scholar] [CrossRef]
- Escuret, V.; Ferraris, O.; Lina, B. The Antiviral Resistance of Influenza Virus. Therapy 2011, 8, 741–762. [Google Scholar] [CrossRef]
- Van Poelvoorde, L.A.E.; Saelens, X.; Thomas, I.; Roosens, N.H. Next-Generation Sequencing: An Eye-Opener for the Surveillance of Antiviral Resistance in Influenza. Trends Biotechnol. 2019, 38, 360–367. [Google Scholar] [CrossRef] [Green Version]
- WHO Laboratory Methodologies for Testing the Antiviral Susceptibility of Influenza Viruses: Neuraminidase Inhibitor (NAI). Available online: https://www.who.int/influenza/gisrs_laboratory/antiviral_susceptibility/nai_overview/en/ (accessed on 1 February 2021).
- Meijer, A.; Rebelo-De-Andrade, H.; Correia, V.; Besselaar, T.; Drager-Dayal, R.; Fry, A.; Gregory, V.; Gubareva, L.; Kageyama, T.; Lackenby, A.; et al. Global Update on the Susceptibility of Human Influenza Viruses to Neuraminidase Inhibitors, 2012–2013. Antivir. Res. 2014, 110, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.I.; Lackenby, A.; Daniels, R.S.; McCauley, J.W.; Pereyaslov, D.; Broberg, E.K.; Meijer, A.; Zambon, M.C. Evaluation of Influenza Virus Antiviral Susceptibility Testing in Europe: Results from the First External Quality Assessment Exercise. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2013, 56, 212–218. [Google Scholar] [CrossRef]
- Okomo-Adhiambo, M.; Mishin, V.P.; Sleeman, K.; Saguar, E.; Guevara, H.; Reisdorf, E.; Griesser, R.H.; Spackman, K.J.; Mendenhall, M.; Carlos, M.P.; et al. Standardizing the Influenza Neuraminidase Inhibition Assay among United States Public Health Laboratories Conducting Virological Surveillance. Antivir. Res. 2016, 128, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Van der Vries, E.; Stittelaar, K.J.; van Amerongen, G.; Veldhuis Kroeze, E.J.B.; de Waal, L.; Fraaij, P.L.A.; Meesters, R.J.; Luider, T.M.; van der Nagel, B.; Koch, B.; et al. Prolonged Influenza Virus Shedding and Emergence of Antiviral Resistance in Immunocompromised Patients and Ferrets. PLoS Pathog. 2013, 9, e1003343. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Honzumi, K.; Sato, T.; Hashimoto, K.; Watanabe, M.; Miyazaki, K.; Kawasaki, Y.; Hosoya, M. Quantitative Analysis of Influenza A (H3N2) E119V and R292K Variants in Clinical Specimens by Real-Time Reverse Transcription Polymerase Chain Reaction. J. Clin. Virol. 2015, 68, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Hindson, B.J.; Ness, K.D.; Masquelier, D.A.; Belgrader, P.; Heredia, N.J.; Makarewicz, A.J.; Bright, I.J.; Lucero, M.Y.; Hiddessen, A.L.; Legler, T.C.; et al. High-Throughput Droplet Digital PCR System for Absolute Quantitation of DNA Copy Number. Anal. Chem. 2011, 83, 8604–8610. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.; Huggett, J.F.; Bushell, C.A.; Cowen, S.; Scott, D.J.; Foy, C.A. Evaluation of Digital PCR for Absolute DNA Quantification. Anal. Chem. 2011, 83, 6474–6484. [Google Scholar] [CrossRef]
- Whale, A.S.; Huggett, J.F.; Cowen, S.; Speirs, V.; Shaw, J.; Ellison, S.; Foy, C.A.; Scott, D.J. Comparison of Microfluidic Digital PCR and Conventional Quantitative PCR for Measuring Copy Number Variation. Nucleic Acids Res. 2012, 40, e82. [Google Scholar] [CrossRef]
- Sanders, R.; Mason, D.J.; Foy, C.A.; Huggett, J.F. Evaluation of Digital PCR for Absolute RNA Quantification. PLoS ONE 2013, 8, e75296. [Google Scholar] [CrossRef]
- Hindson, C.M.; Chevillet, J.R.; Briggs, H.A.; Gallichotte, E.N.; Ruf, I.K.; Hindson, B.J.; Vessella, R.L.; Tewari, M. Absolute Quantification by Droplet Digital PCR versus Analog Real-Time PCR. Nat. Methods 2013, 10, 1003–1005. [Google Scholar] [CrossRef]
- Demeke, T.; Dobnik, D. Critical Assessment of Digital PCR for the Detection and Quantification of Genetically Modified Organisms. Anal. Bioanal. Chem. 2018, 410, 4039–4050. [Google Scholar] [CrossRef] [Green Version]
- Van Poelvoorde, L.A.E.; Gand, M.; Fraiture, M.-A.; De Keersmaecker, S.C.J.; Verhaegen, B.; Van Hoorde, K.; Cay, A.B.; Balmelle, N.; Herman, P.; Roosens, N. Strategy to Develop and Evaluate a Multiplex RT-DdPCR in Response to SARS-CoV-2 Genomic Evolution. CIMB 2021, 43, 134. [Google Scholar] [CrossRef]
- Devonshire, A.S.; O’Sullivan, D.M.; Honeyborne, I.; Jones, G.; Karczmarczyk, M.; Pavšič, J.; Gutteridge, A.; Milavec, M.; Mendoza, P.; Schimmel, H.; et al. The Use of Digital PCR to Improve the Application of Quantitative Molecular Diagnostic Methods for Tuberculosis. BMC Infect. Dis. 2016, 16, 366. [Google Scholar] [CrossRef]
- McDermott, G.P.; Do, D.; Litterst, C.M.; Maar, D.; Hindson, C.M.; Steenblock, E.R.; Legler, T.C.; Jouvenot, Y.; Marrs, S.H.; Bemis, A.; et al. Multiplexed Target Detection Using DNA-Binding Dye Chemistry in Droplet Digital PCR. Anal. Chem. 2013, 85, 11619–11627. [Google Scholar] [CrossRef]
- Taylor, S.C.; Carbonneau, J.; Shelton, D.N.; Boivin, G. Optimization of Droplet Digital PCR from RNA and DNA Extracts with Direct Comparison to RT-QPCR: Clinical Implications for Quantification of Oseltamivir-Resistant Subpopulations. J. Virol. Methods 2015, 224, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Suo, T.; Liu, X.; Feng, J.; Guo, M.; Hu, W.; Guo, D.; Ullah, H.; Yang, Y.; Zhang, Q.; Wang, X.; et al. DdPCR: A More Accurate Tool for SARS-CoV-2 Detection in Low Viral Load Specimens. Emerg. Microbes Infect. 2020, 9, 1259–1268. [Google Scholar] [CrossRef]
- Whale, A.S.; Bushell, C.A.; Grant, P.R.; Cowen, S.; Gutierrez-Aguirre, I.; O’Sullivan, D.M.; Žel, J.; Milavec, M.; Foy, C.A.; Nastouli, E.; et al. Detection of Rare Drug Resistance Mutations by Digital PCR in a Human Influenza A Virus Model System and Clinical Samples. J. Clin. Microbiol. 2016, 54, 392–400. [Google Scholar] [CrossRef] [Green Version]
- Pichon, M.; Gaymard, A.; Josset, L.; Valette, M.; Millat, G.; Lina, B.; Escuret, V. Characterization of Oseltamivir-Resistant Influenza Virus Populations in Immunosuppressed Patients Using Digital-Droplet PCR: Comparison with QPCR and next Generation Sequencing Analysis. Antivir. Res. 2017, 145, 160–167. [Google Scholar] [CrossRef]
- World Health Organization. CDC Protocol of Realtime RTPCR for Influenza A(H1N1); World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Da Silva, D.V.; Nordholm, J.; Madjo, U.; Pfeiffer, A.; Daniels, R. Assembly of Subtype 1 Influenza Neuraminidase Is Driven by Both the Transmembrane and Head Domains. J. Biol. Chem. 2013, 288, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Dohna, H.Z.; Cardona, C.J.; Miller, J.; Carpenter, T.E. Emergence and Genetic Variation of Neuraminidase Stalk Deletions in Avian Influenza Viruses. PLoS ONE 2011, 6, e14722. [Google Scholar] [CrossRef]
- Claas, E.C.; Osterhaus, A.D.; van Beek, R.; De Jong, J.C.; Rimmelzwaan, G.F.; Senne, D.A.; Krauss, S.; Shortridge, K.F.; Webster, R.G. Human Influenza A H5N1 Virus Related to a Highly Pathogenic Avian Influenza Virus. Lancet 1998, 351, 472–477. [Google Scholar] [CrossRef]
- Castrucci, M.R.; Kawaoka, Y. Biologic Importance of Neuraminidase Stalk Length in Influenza A Virus. J. Virol. 1993, 67, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Matrosovich, M.; Zhou, N.; Kawaoka, Y.; Webster, R. The Surface Glycoproteins of H5 Influenza Viruses Isolated from Humans, Chickens, and Wild Aquatic Birds Have Distinguishable Properties. J. Virol. 1999, 73, 1146–1155. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, T.W.; Munier, S.; Larcher, T.; Soubieux, D.; Ledevin, M.; Esnault, E.; Tourdes, A.; Croville, G.; Guerin, J.-L.; Quere, P.; et al. Length Variations in the NA Stalk of an H7N1 Influenza Virus Have Opposite Effects on Viral Excretion in Chickens and Ducks. J. Virol. 2012, 86, 584–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhirnov, O.P.; Vorobjeva, I.V.; Saphonova, O.A.; Poyarkov, S.V.; Ovcharenko, A.V.; Anhlan, D.; Malyshev, N.A. Structural and Evolutionary Characteristics of HA, NA, NS and M Genes of Clinical Influenza A/H3N2 Viruses Passaged in Human and Canine Cells. J. Clin. Virol. 2009, 45, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, E.M.; Song, H.; Pena, L.; Perez, D.R. A 27-Amino-Acid Deletion in the Neuraminidase Stalk Supports Replication of an Avian H2N2 Influenza A Virus in the Respiratory Tract of Chickens. J. Virol. 2010, 84, 11831–11840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirst, G.K. Adsorption of Influenza Hemagglutinins and Virus by Red Blood Cells. J. Exp. Med. 1942, 76, 195–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, A. Neuraminidase: The Specific Enzyme of Influenza Virus and Vibrio Cholerae. Biochim. Biophys. Acta 1957, 23, 645–646. [Google Scholar] [CrossRef]
- Palese, P.; Compans, R.W. Inhibition of Influenza Virus Replication in Tissue Culture by 2-Deoxy-2,3-Dehydro-N-Trifluoroacetylneuraminic Acid (FANA): Mechanism of Action. J. Gen. Virol. 1976, 33, 159–163. [Google Scholar] [CrossRef]
- Russell, R.J.; Haire, L.F.; Stevens, D.J.; Collins, P.J.; Lin, Y.P.; Blackburn, G.M.; Hay, A.J.; Gamblin, S.J.; Skehel, J.J. The Structure of H5N1 Avian Influenza Neuraminidase Suggests New Opportunities for Drug Design. Nature 2006, 443, 45–49. [Google Scholar] [CrossRef]
- Webster, R.G.; Laver, W.G. Preparation and Properties of Antibody Directed Specifically against the Neuraminidase of Influenza Virus. J. Immunol. 1967, 99, 49–55. [Google Scholar] [CrossRef]
- Varghese, J.N.; Laver, W.G.; Colman, P.M. Structure of the Influenza Virus Glycoprotein Antigen Neuraminidase at 2.9 Å Resolution. Nature 1983, 303, 35–40. [Google Scholar] [CrossRef]
- Ugbaja, S.C.; Mtambo, S.E.; Mushebenge, A.G.; Appiah-Kubi, P.; Abubakar, B.H.; Ntuli, M.L.; Kumalo, H.M. Structural Investigations and Binding Mechanisms of Oseltamivir Drug Resistance Conferred by the E119V Mutation in Influenza H7N9 Virus. Molecules 2022, 27, 4376. [Google Scholar] [CrossRef]
- Kutyavin, I.V.; Afonina, I.A.; Mills, A.; Gorn, V.V.; Lukhtanov, E.A.; Belousov, E.S.; Singer, M.J.; Walburger, D.K.; Lokhov, S.G.; Gall, A.A.; et al. 3’-Minor Groove Binder-DNA Probes Increase Sequence Specificity at PCR Extension Temperatures. Nucleic Acids Res. 2000, 28, 655–661. [Google Scholar] [CrossRef]
- Lampejo, T. Influenza and Antiviral Resistance: An Overview. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1201–1208. [Google Scholar] [CrossRef]
- Deng, Y.M.; Caldwell, N.; Hurt, A.; Shaw, T.; Kelso, A.; Chidlow, G.; Williams, S.; Smith, D.; Barr, I. A Comparison of Pyrosequencing and Neuraminidase Inhibition Assays for the Detection of Oseltamivir-Resistant Pandemic Influenza A(H1N1) 2009 Viruses. Antivir. Res. 2011, 90, 87–91. [Google Scholar] [CrossRef]
- Burnham, A.J.; Armstrong, J.; Lowen, A.C.; Webster, R.G.; Govorkova, E.A. Competitive Fitness of Influenza B Viruses with Neuraminidase Inhibitor-Resistant Substitutions in a Coinfection Model of the Human Airway Epithelium. J. Virol. 2015, 89, 4575–4587. [Google Scholar] [CrossRef] [Green Version]
- Heijnen, L.; Elsinga, G.; de Graaf, M.; Molenkamp, R.; Koopmans, M.P.G.; Medema, G. Droplet Digital RT-PCR to Detect SARS-CoV-2 Variants of Concern in Wastewater. medRxiv 2021. [Google Scholar] [CrossRef]
- Xagoraraki, I.; O’Brien, E. Wastewater-Based Epidemiology for Early Detection of Viral Outbreaks. In Women in Engineering and Science; O’Bannon, D.J., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 75–97. ISBN 978-3-030-17818-5. [Google Scholar]
- Howard, W.; Hayman, A.; Lackenby, A.; Whiteley, A.; Londt, B.; Banks, J.; McCauley, J.; Barclay, W. The Development of a Reverse Genetics System Enabling the Rescue of Recombinant Avian Influenza Virus, A/Turkey/England/50-92/91 (H5N1). Avian Dis. 2007, 2, 393–395. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Poelvoorde, L.A.E.; Dufrasne, F.E.; Van Gucht, S.; Saelens, X.; Roosens, N.H.C. Development of Digital Droplet PCR Targeting the Influenza H3N2 Oseltamivir-Resistant E119V Mutation and Its Performance through the Use of Reverse Genetics Mutants. Curr. Issues Mol. Biol. 2023, 45, 2521-2532. https://doi.org/10.3390/cimb45030165
Van Poelvoorde LAE, Dufrasne FE, Van Gucht S, Saelens X, Roosens NHC. Development of Digital Droplet PCR Targeting the Influenza H3N2 Oseltamivir-Resistant E119V Mutation and Its Performance through the Use of Reverse Genetics Mutants. Current Issues in Molecular Biology. 2023; 45(3):2521-2532. https://doi.org/10.3390/cimb45030165
Chicago/Turabian StyleVan Poelvoorde, Laura A. E., François E. Dufrasne, Steven Van Gucht, Xavier Saelens, and Nancy H. C. Roosens. 2023. "Development of Digital Droplet PCR Targeting the Influenza H3N2 Oseltamivir-Resistant E119V Mutation and Its Performance through the Use of Reverse Genetics Mutants" Current Issues in Molecular Biology 45, no. 3: 2521-2532. https://doi.org/10.3390/cimb45030165