
Isodorsmanin A Prevents Inflammatory Response in LPS-Stimulated Macrophages by Inhibiting the JNK and NF-κB Signaling Pathways

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Measurement of Cell Viability
2.3. Measurement of Inflammatory Mediators and Proinflammatory Cytokines
2.4. Nuclear and Cytosolic Extraction
2.5. Western Blotting
2.6. RT-qPCR Analysis
2.7. Statistical Analysis
3. Results
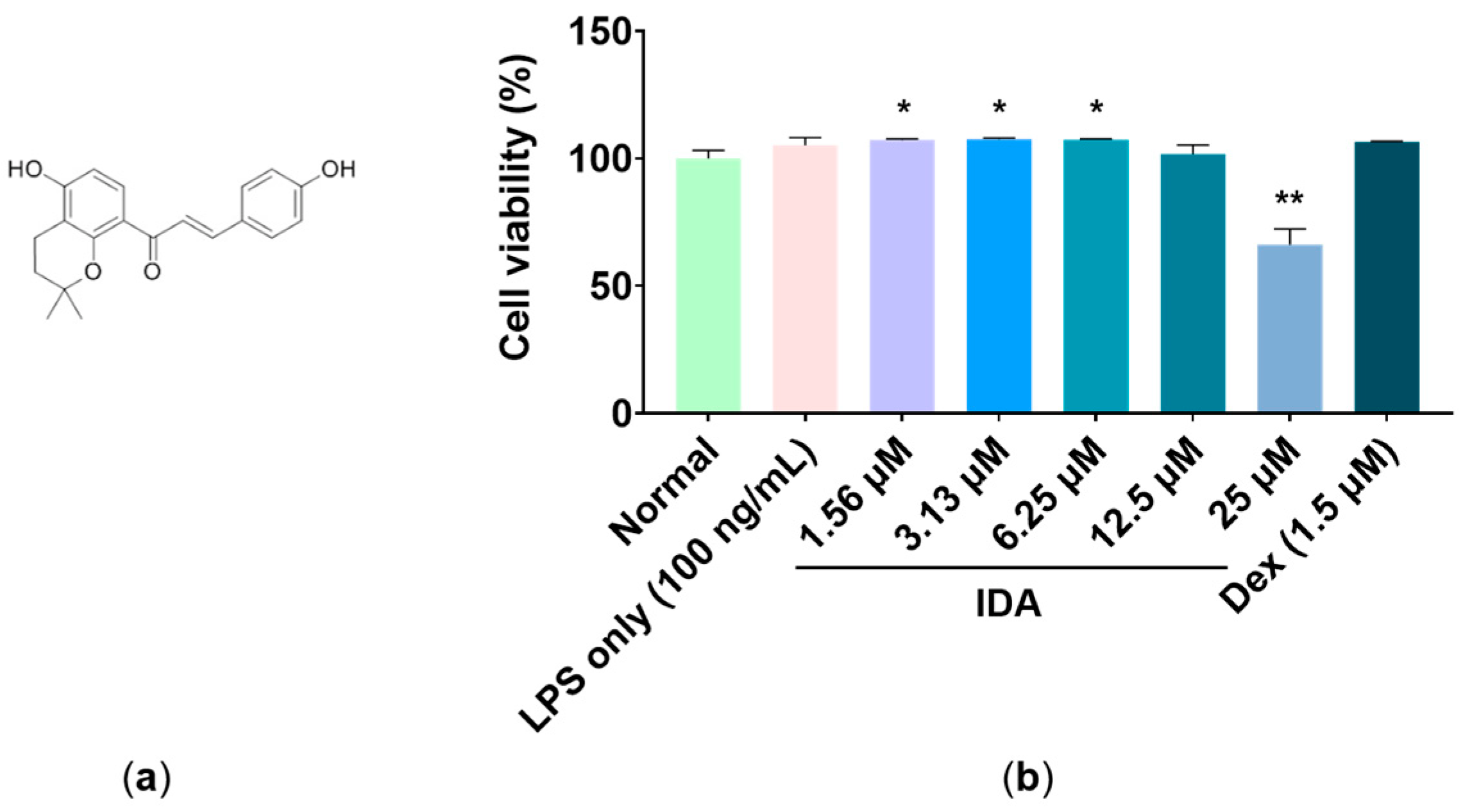
3.1. Effect of IDA on Cell Viability
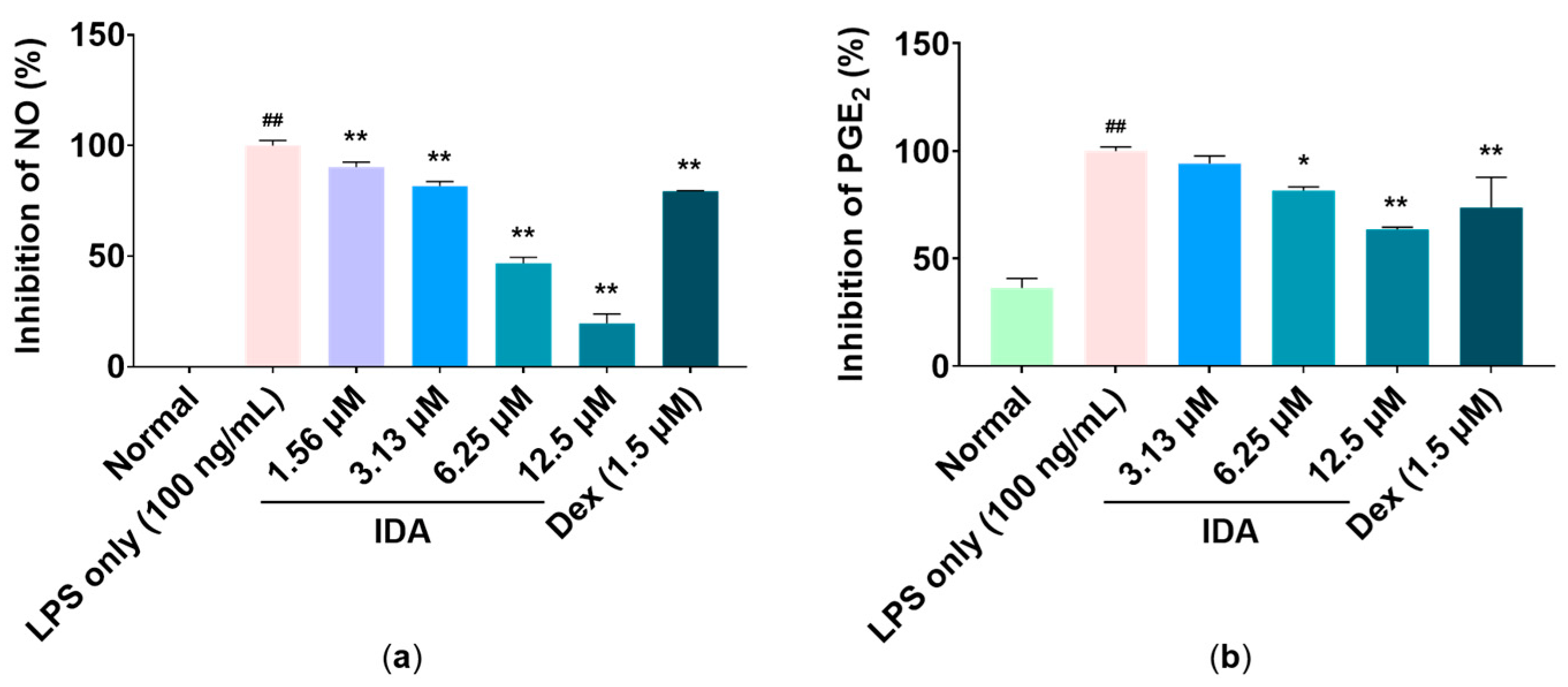
3.2. Effect of IDA on Inflammatory Mediator Production
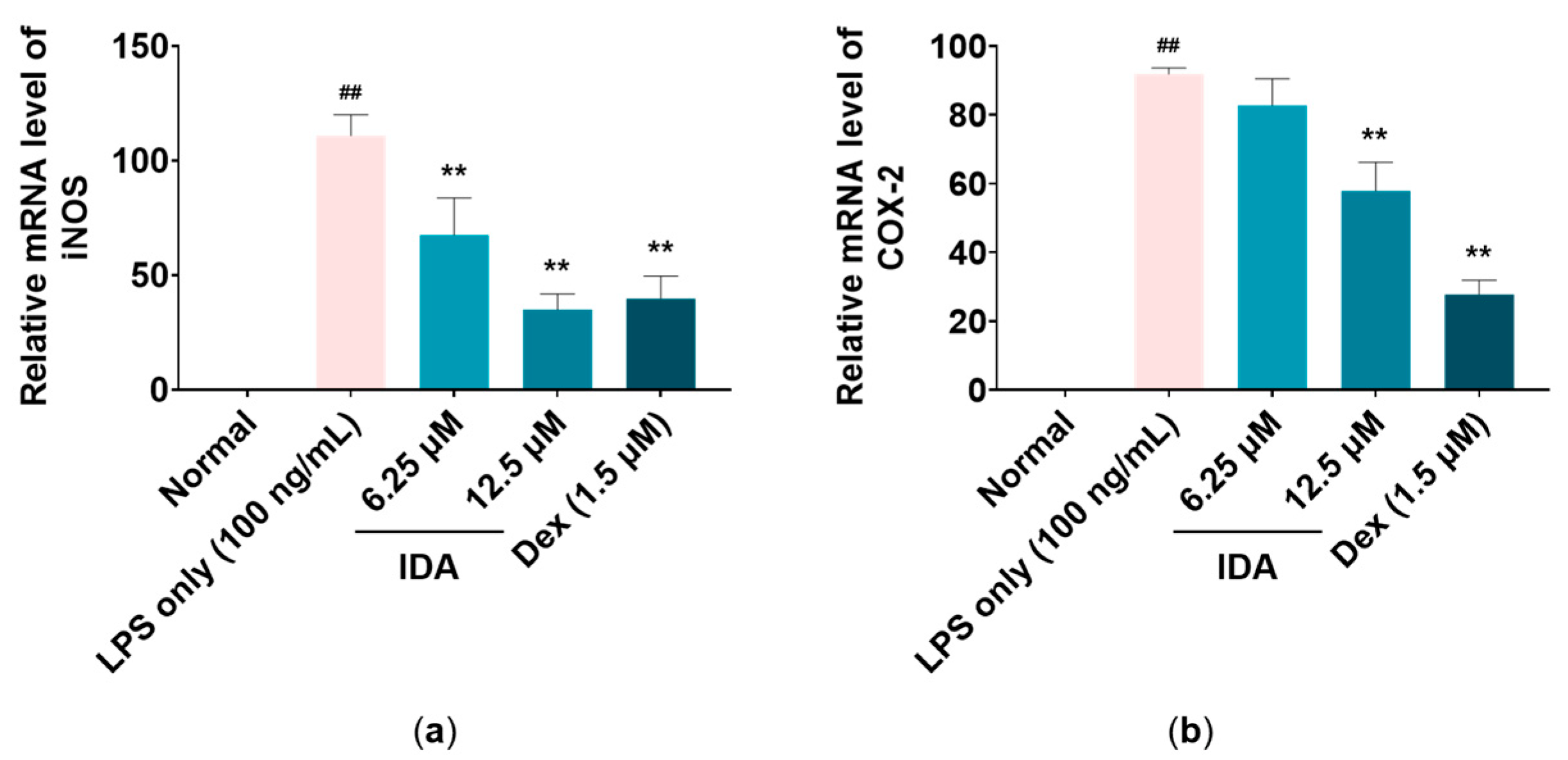
3.3. Effect of IDA on iNOS and COX-2 mRNA Levels
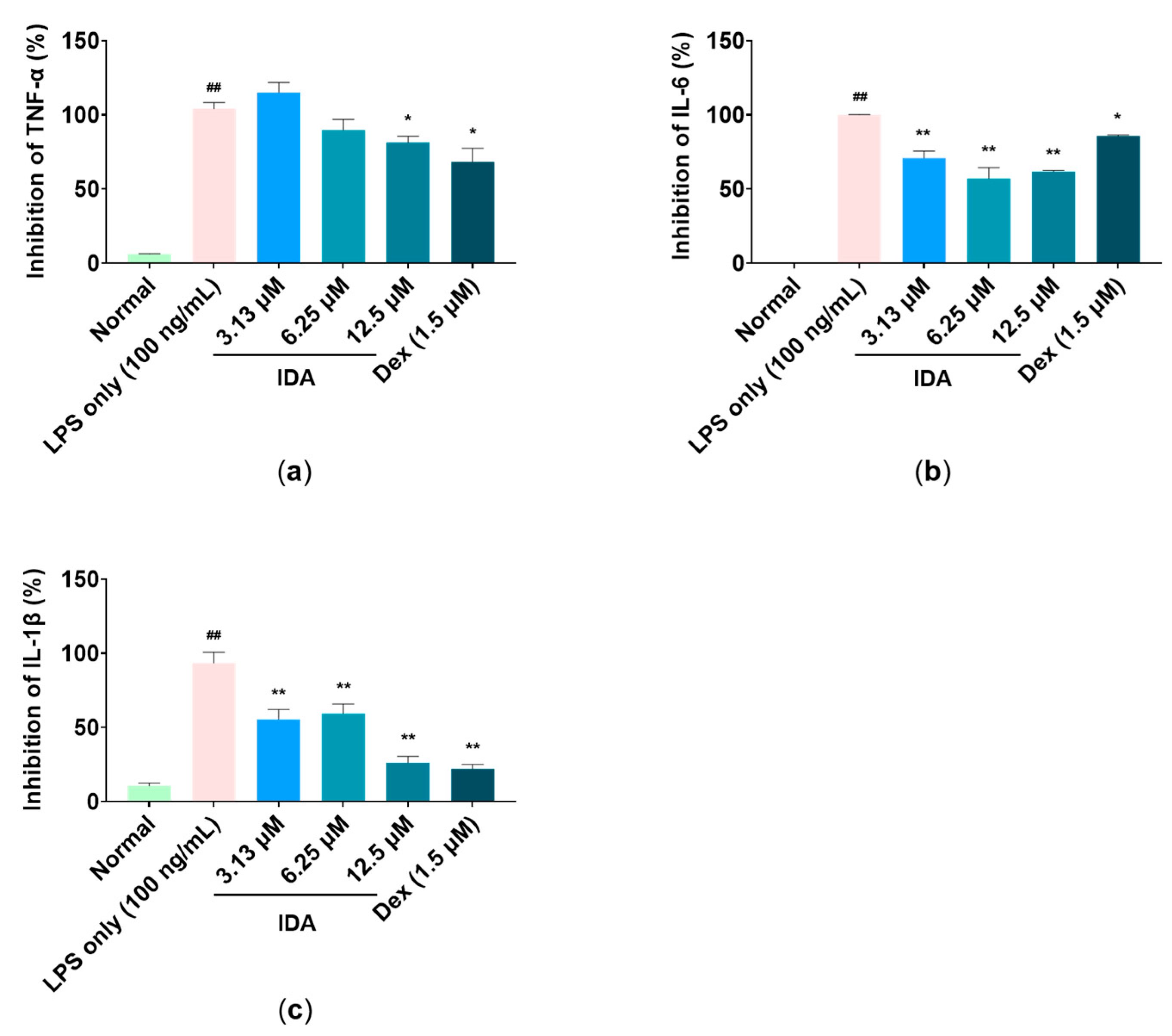
3.4. Effect of IDA on Inflammatory Cytokine Expression
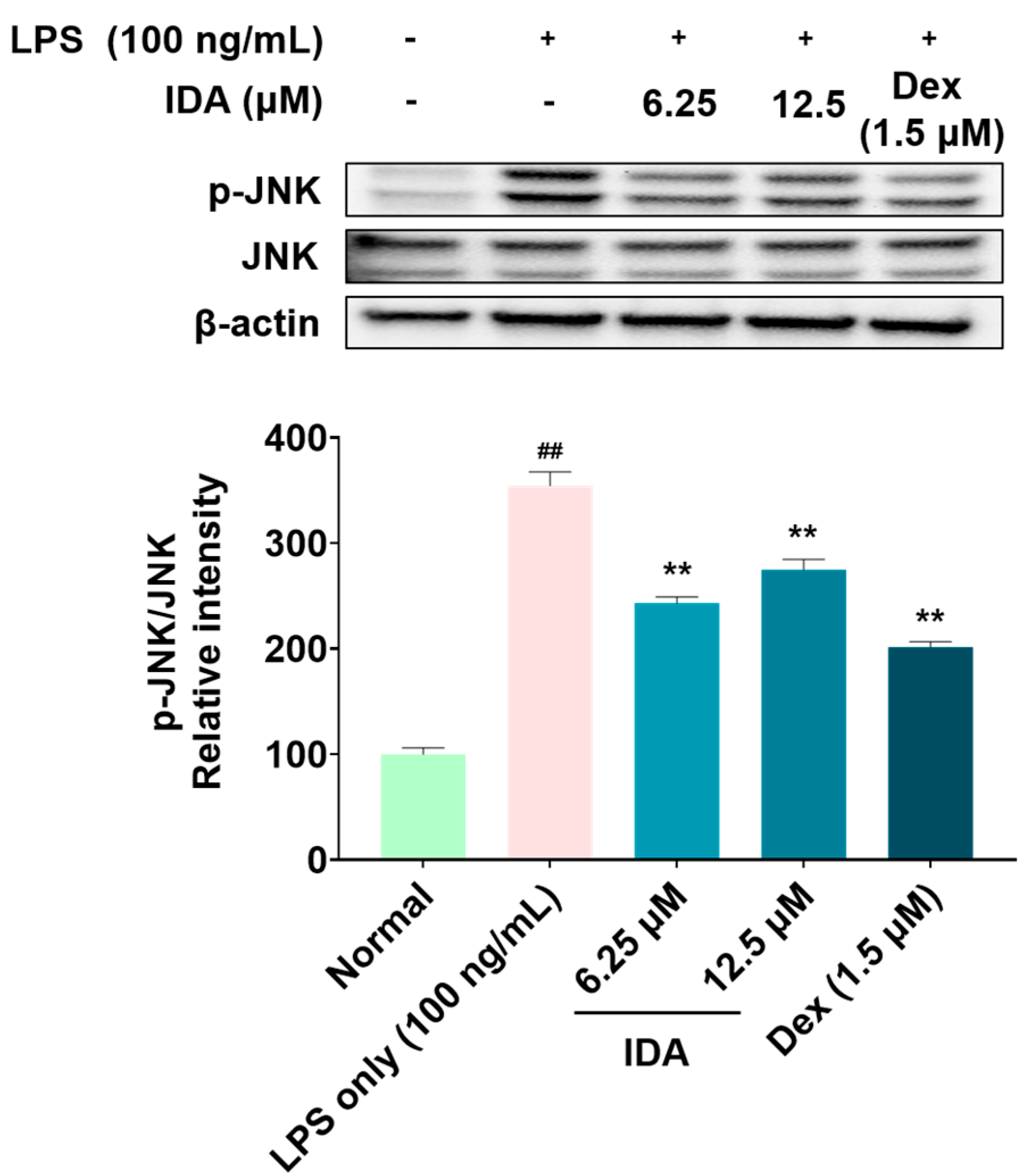
3.5. Effect of IDA on the Phosphorylation of MAPK
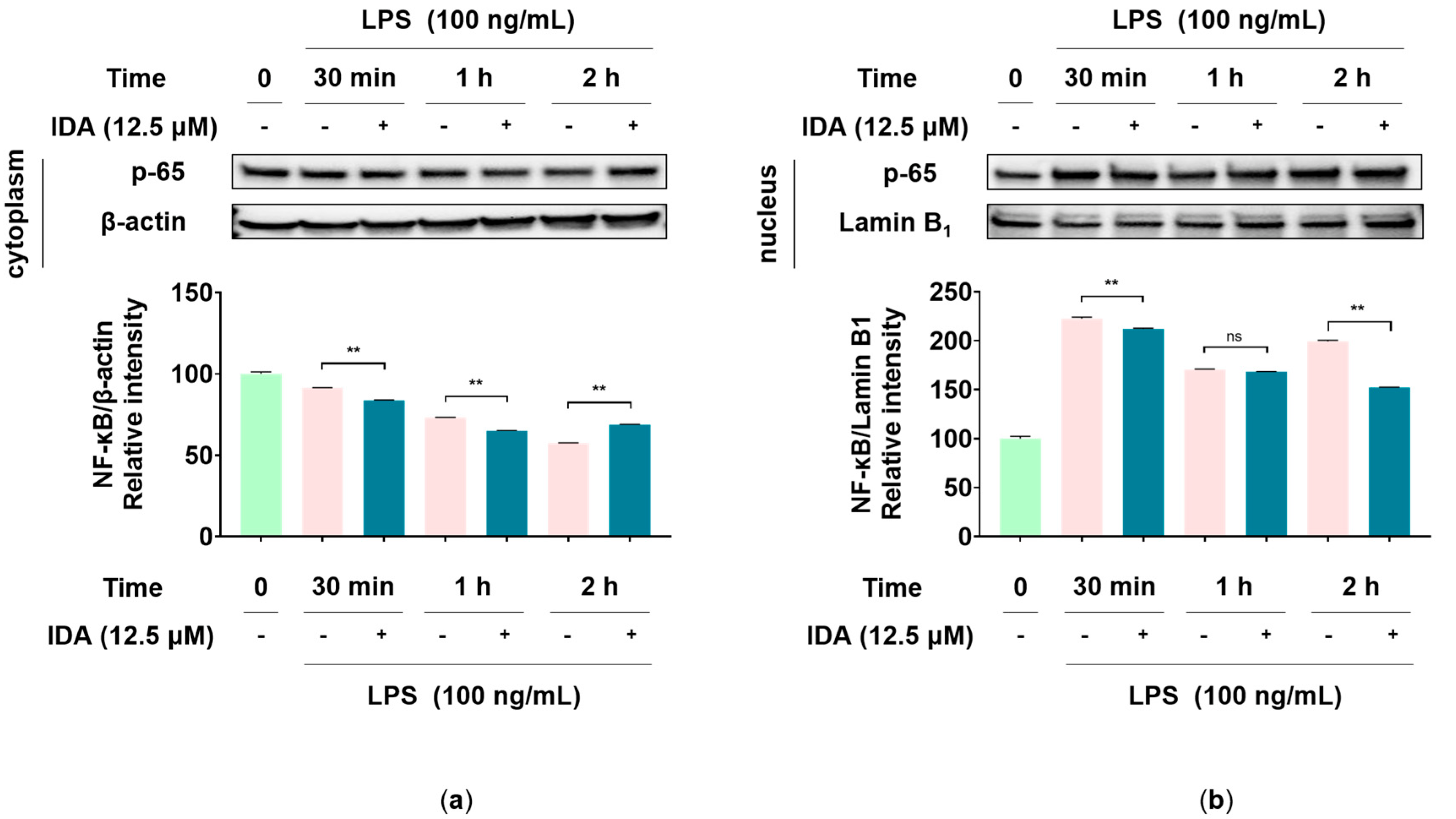
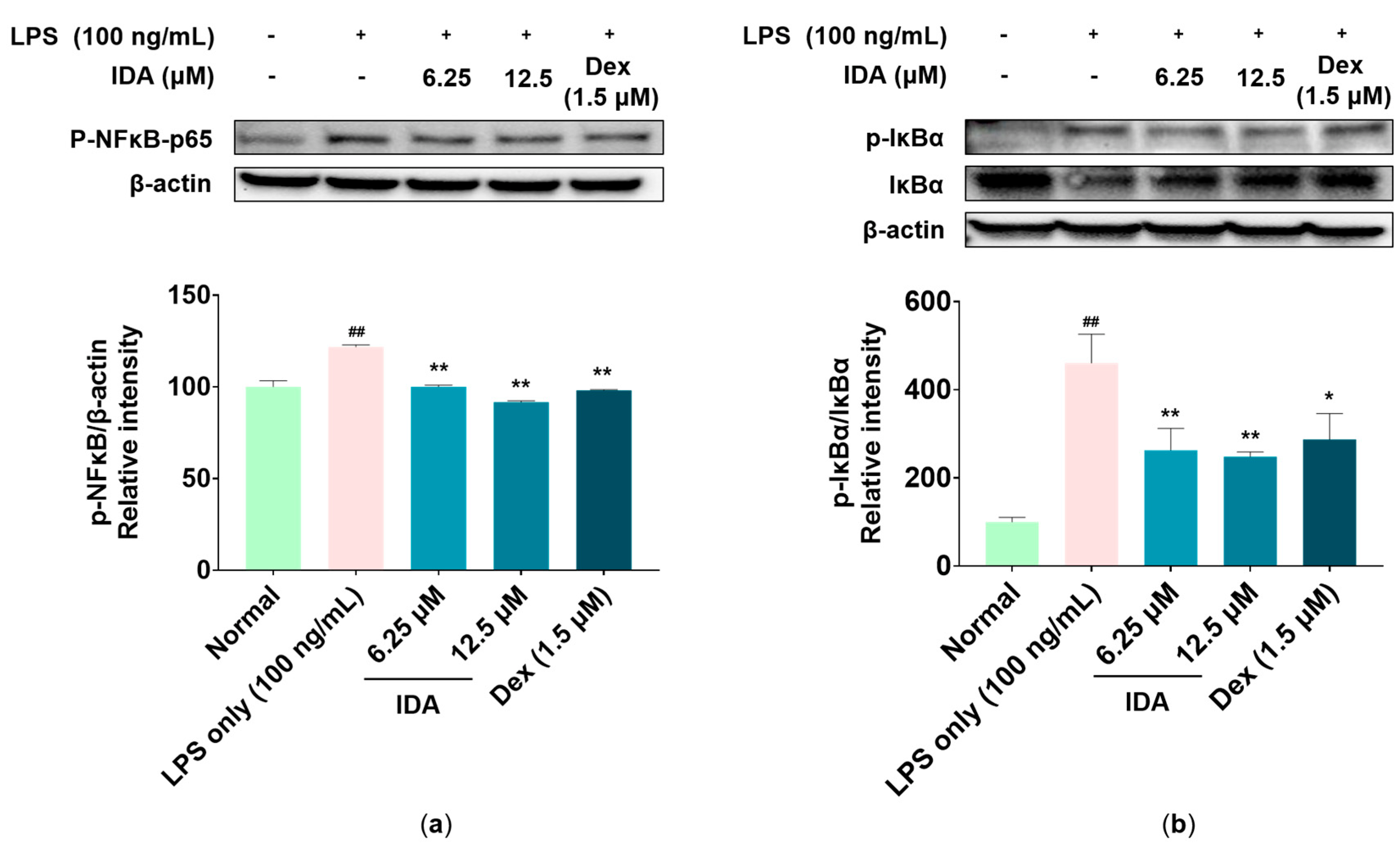
3.6. Effect of IDA on the NF-κB Signaling Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.; Nam, J.W.; Kang, H.J.; Windono, T.; Seo, E.K.; Lee, K.T. Zedoarondiol isolated from the rhizoma of Curcuma heyneana is involved in the inhibition of iNOS, COX-2 and pro-inflammatory cytokines via the downregulation of NF-kappaB pathway in LPS-stimulated murine macrophages. Int. Immunopharmacol. 2009, 9, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Kunnumakkara, A.B.; Sailo, B.L.; Banik, K.; Harsha, C.; Prasad, S.; Gupta, S.C.; Bharti, A.C.; Aggarwal, B.B. Chronic diseases, inflammation, and spices: How are they linked? J. Transl. Med. 2018, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- Nasef, N.A.; Mehta, S.; Ferguson, L.R. Susceptibility to chronic inflammation: An update. Arch. Toxicol. 2017, 91, 1131–1141. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Hong, S.H.; Park, S.D.; Ahn, S.G.; Yoon, J.H.; Kwon, B.M.; Kim, S.A. 2′-Benzoyloxycinnamaldehyde inhibits nitric oxide production in lipopolysaccharide-stimulated RAW 264.7 cells via regulation of AP-1 pathway. Eur. J. Pharm. 2012, 696, 179–186. [Google Scholar] [CrossRef]
- Van den Bossche, J.; O’Neill, L.A.; Menon, D. Macrophage Immunometabolism: Where Are We (Going)? Trends Immunol. 2017, 38, 395–406. [Google Scholar] [CrossRef]
- Vannini, F.; Kashfi, K.; Nath, N. The dual role of iNOS in cancer. Redox Biol. 2015, 6, 334–343. [Google Scholar] [CrossRef]
- Ally, A.; Powell, I.; Ally, M.M.; Chaitoff, K.; Nauli, S.M. Role of neuronal nitric oxide synthase on cardiovascular functions in physiological and pathophysiological states. Nitric Oxide 2020, 102, 52–73. [Google Scholar] [CrossRef]
- Cyr, A.R.; Huckaby, L.V.; Shiva, S.S.; Zuckerbraun, B.S. Nitric Oxide and Endothelial Dysfunction. Crit. Care Clin. 2020, 36, 307–321. [Google Scholar] [CrossRef]
- Farlik, M.; Reutterer, B.; Schindler, C.; Greten, F.; Vogl, C.; Müller, M.; Decker, T. Nonconventional Initiation Complex Assembly by STAT and NF-κB Transcription Factors Regulates Nitric Oxide Synthase Expression. Immunity 2010, 33, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Pautz, A.; Art, J.; Hahn, S.; Nowag, S.; Voss, C.; Kleinert, H. Regulation of the expression of inducible nitric oxide synthase. Nitric Oxide 2010, 23, 75–93. [Google Scholar] [CrossRef]
- Pereira, B.P.; do Vale, G.T.; Ceron, C.S. The role of nitric oxide in renovascular hypertension: From the pathophysiology to the treatment. Naunyn Schmiedebergs Arch Pharm. 2022, 395, 121–131. [Google Scholar] [CrossRef]
- Kawahara, K.; Hohjoh, H.; Inazumi, T.; Tsuchiya, S.; Sugimoto, Y. Prostaglandin E2-induced inflammation: Relevance of prostaglandin E receptors. Biochim Biophys Acta 2015, 1851, 414–421. [Google Scholar] [CrossRef]
- Kitanaka, T.; Nakano, R.; Kitanaka, N.; Kimura, T.; Okabayashi, K.; Narita, T.; Sugiya, H. JNK activation is essential for activation of MEK/ERK signaling in IL-1β-induced COX-2 expression in synovial fibroblasts. Sci. Rep. 2017, 7, 39914. [Google Scholar] [CrossRef]
- Fan, H.; Gao, Z.; Ji, K.; Li, X.; Wu, J.; Liu, Y.; Wang, X.; Liang, H.; Liu, Y.; Li, X.; et al. The in vitro and in vivo anti-inflammatory effect of osthole, the major natural coumarin from Cnidium monnieri (L.) Cuss, via the blocking of the activation of the NF-κB and MAPK/p38 pathways. Phytomedicine 2019, 58, 152864. [Google Scholar] [CrossRef]
- Han, J.M.; Lee, E.K.; Gong, S.Y.; Sohng, J.K.; Kang, Y.J.; Jung, H.J. Sparassis crispa exerts anti-inflammatory activity via suppression of TLR-mediated NF-κB and MAPK signaling pathways in LPS-induced RAW264.7 macrophage cells. J. Ethnopharmacol. 2019, 231, 10–18. [Google Scholar] [CrossRef]
- Hu, T.Y.; Ju, J.M.; Mo, L.H.; Ma, L.; Hu, W.H.; You, R.R.; Chen, X.Q.; Chen, Y.Y.; Liu, Z.Q.; Qiu, S.Q.; et al. Anti-inflammation action of xanthones from Swertia chirayita by regulating COX-2/NF-κB/MAPKs/Akt signaling pathways in RAW 264.7 macrophage cells. Phytomedicine 2019, 55, 214–221. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Moens, U.; Kostenko, S.; Sveinbjørnsson, B. The Role of Mitogen-Activated Protein Kinase-Activated Protein Kinases (MAPKAPKs) in Inflammation. Genes 2013, 4, 101–133. [Google Scholar] [CrossRef]
- Prashar, H.; Chawla, A.; Sharma, A.; Kharb, R. Chalcone as a versatile moiety for diverse pharmacological activities. Int. J. Pharm. Sci. Res. 2012, 3, 1913–1927. [Google Scholar]
- Kostanecki, S.V.; Tambor, J. Ueber die sechs isomeren Monooxybenzalacetophenone (Monooxychalkone). Ber. Der Dtsch. Chem. Ges. 1899, 32, 1921–1926. [Google Scholar] [CrossRef]
- Díaz-Tielas, C.; Graña, E.; Reigosa, M.J.; Sánchez-Moreira, S.M. Biological Activities and Novel Applications of Chalcones 1. Planta Daninha 2016, 34, 607–616. [Google Scholar] [CrossRef]
- Singhal, M.; Paul, A.; Singh, H. Synthesis and characterisation of some novel chalcone derivatives: An intermediate for various hetreocyclics compounds. Int. J. Pharm. Innov. 2011, 1, 1–7. [Google Scholar]
- Lee, A.; Yang, H.; Kim, T.; Ha, H.; Hwang, Y.-H. Identification and pharmacokinetics of bioavailable anti-resorptive phytochemicals after oral administration of Psoralea corylifolia L. Biomed. Pharmacother. 2021, 144, 112300. [Google Scholar] [CrossRef] [PubMed]
- Ali, J.; Akhtar, N.; Sultana, Y.; Baboota, S.; Ahuja, A. Antipsoriatic microemulsion gel formulations for topical drug delivery of babchi oil (Psoralea corylifolia). Methods Find. Exp. Clin. Pharmacol. 2008, 30, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Khatune, N.A.; Islam, M.E.; Haque, M.E.; Khondkar, P.; Rahman, M.M. Antibacterial compounds from the seeds of Psoralea corylifolia. Fitoterapia 2004, 75, 228–230. [Google Scholar] [CrossRef]
- Lee, S.W.; Yun, B.R.; Kim, M.H.; Park, C.S.; Lee, W.S.; Oh, H.M.; Rho, M.C. Phenolic compounds isolated from Psoralea corylifolia inhibit IL-6-induced STAT3 activation. Planta Med. 2012, 78, 903–906. [Google Scholar] [CrossRef]
- Potapenko, A.; Sukhorukov, V.L.; Davidov, B.V. A comparison between skin-photosensitizing (344 nm) activities of 8-methoxypsoralen and angelicin. Experientia 1984, 40, 264–265. [Google Scholar] [CrossRef]
- Song, P.; Yang, X.Z.; Yuan, J.Q. Cytotoxic constituents from Psoralea corylifolia. J. Asian Nat. Prod. Res. 2013, 15, 624–630. [Google Scholar] [CrossRef]
- Tsai, W.J.; Hsin, W.C.; Chen, C.C. Antiplatelet flavonoids from seeds of Psoralea corylifolia. J. Nat. Prod. 1996, 59, 671–672. [Google Scholar] [CrossRef]
- Xiao, G.; Li, G.; Chen, L.; Zhang, Z.; Yin, J.J.; Wu, T.; Cheng, Z.; Wei, X.; Wang, Z. Isolation of antioxidants from Psoralea corylifolia fruits using high-speed counter-current chromatography guided by thin layer chromatography-antioxidant autographic assay. J. Chromatography. A 2010, 1217, 5470–5476. [Google Scholar] [CrossRef]
- Seo, E.; Truong, C.-S.; Jun, H.-S. Psoralea corylifolia L. seed extract attenuates dexamethasone-induced muscle atrophy in mice by inhibition of oxidative stress and inflammation. J. Ethnopharmacol. 2022, 296, 115490. [Google Scholar] [CrossRef]
- Liu, F.; Sun, G.Q.; Gao, H.Y.; Li, R.S.; Soromou, L.W.; Chen, N.; Deng, Y.H.; Feng, H.H. Angelicin regulates LPS-induced inflammation via inhibiting MAPK/NF-κB pathways. J. Surg. Res. 2013, 185, 300–309. [Google Scholar] [CrossRef]
- Li, H.; Xu, J.; Li, X.; Hu, Y.; Liao, Y.; Zhou, W.; Song, Z. Anti-inflammatory activity of psoralen in human periodontal ligament cells via estrogen receptor signaling pathway. Sci. Rep. 2021, 11, 8754. [Google Scholar] [CrossRef]
- Xu, Q.X.; Hu, Y.; Li, G.Y.; Xu, W.; Zhang, Y.T.; Yang, X.W. Multi-Target Anti-Alzheimer Activities of Four Prenylated Compounds from Psoralea Fructus. Molecules 2018, 23, 614. [Google Scholar] [CrossRef]
- Hung, Y.L.; Wang, S.C.; Suzuki, K.; Fang, S.H.; Chen, C.S.; Cheng, W.C.; Su, C.C.; Yeh, H.C.; Tu, H.P.; Liu, P.L.; et al. Bavachin attenuates LPS-induced inflammatory response and inhibits the activation of NLRP3 inflammasome in macrophages. Phytomedicine 2019, 59, 152785. [Google Scholar] [CrossRef]
- Pae, H.-O.; Cho, H.; Oh, G.-S.; Kim, N.-Y.; Song, E.-K.; Kim, Y.-C.; Yun, Y.-G.; Kang, C.-L.; Kim, J.-D.; Kim, J.-M.; et al. Bakuchiol from Psoralea corylifolia inhibits the expression of inducible nitric oxide synthase gene via the inactivation of nuclear transcription factor-κB in RAW 264.7 macrophages. Int. Immunopharmacol. 2001, 1, 1849–1855. [Google Scholar] [CrossRef]
- Rao, K.M.K. MAP kinase activation in macrophages. J. Leukoc. Biol. 2001, 69, 3–10. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- Arango Duque, G.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Gandhi, J.; Khera, L.; Gaur, N.; Paul, C.; Kaul, R. Role of Modulator of Inflammation Cyclooxygenase-2 in Gammaherpesvirus Mediated Tumorigenesis. Front. Microbiol. 2017, 8, 538. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.; Domínguez, J.N.; Charris, J.E.; Lobo, G.; Payá, M.; Ferrándiz, M.L. Synthesis and inhibitory activity of dimethylamino-chalcone derivatives on the induction of nitric oxide synthase. Eur. J. Med. Chem. 2002, 37, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.; Payá, M.; Devesa, I.; Dominguez, J.N.; Ferrándiz, M.L. Therapeutic administration of 3,4,5-trimethoxy-4′-fluorochalcone, a selective inhibitor of iNOS expression, attenuates the development of adjuvant-induced arthritis in rats. Naunyn Schmiedebergs Arch Pharm. 2003, 368, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.; Payá, M.; Domínguez, J.N.; Ferrándiz, M.L. ttCH, a selective inhibitor of inducible nitric oxide synthase expression with antiarthritic properties. Eur. J. Pharm. 2003, 465, 183–189. [Google Scholar] [CrossRef]
- Zhao, F.; Nozawa, H.; Daikonnya, A.; Kondo, K.; Kitanaka, S. Inhibitors of nitric oxide production from hops (Humulus lupulus L.). Biol Pharm Bull 2003, 26, 61–65. [Google Scholar] [CrossRef]
- Emam, S.H.; Sonousi, A.; Osman, E.O.; Hwang, D.; Kim, G.-D.; Hassan, R.A. Design and synthesis of methoxyphenyl- and coumarin-based chalcone derivatives as anti-inflammatory agents by inhibition of NO production and down-regulation of NF-κB in LPS-induced RAW264.7 macrophage cells. Bioorganic Chem. 2021, 107, 104630. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesth. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Kim, K.N.; Ko, Y.J.; Kang, M.C.; Yang, H.M.; Roh, S.W.; Oda, T.; Jeon, Y.J.; Jung, W.K.; Heo, S.J.; Yoon, W.J.; et al. Anti-inflammatory effects of trans-1,3-diphenyl-2,3-epoxypropane-1-one mediated by suppression of inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. Food Chem. Toxicol. 2013, 53, 371–375. [Google Scholar] [CrossRef]
- Arbabi, S.; Maier, R.V. Mitogen-activated protein kinases. Crit. Care Med. 2002, 30, S74–S79. [Google Scholar] [CrossRef]
- Shakoory, B.; Fitzgerald, S.M.; Lee, S.A.; Chi, D.S.; Krishnaswamy, G. The role of human mast cell-derived cytokines in eosinophil biology. J. Interferon. Cytokine Res. 2004, 24, 271–281. [Google Scholar] [CrossRef]
- Kriete, A.; Mayo, K.L. Atypical pathways of NF-kappaB activation and aging. Exp. Gerontol. 2009, 44, 250–255. [Google Scholar] [CrossRef]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell. Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Ban, H.S.; Suzuki, K.; Lim, S.S.; Jung, S.H.; Lee, S.; Ji, J.; Lee, H.S.; Lee, Y.S.; Shin, K.H.; Ohuchi, K. Inhibition of lipopolysaccharide-induced expression of inducible nitric oxide synthase and tumor necrosis factor-alpha by 2′-hydroxychalcone derivatives in RAW 264.7 cells. Biochem. Pharm. 2004, 67, 1549–1557. [Google Scholar] [CrossRef]
- Dhar, R.; Kimseng, R.; Chokchaisiri, R.; Hiransai, P.; Utaipan, T.; Suksamrarn, A.; Chunglok, W. 2′,4-Dihydroxy-3′,4′,6′-trimethoxychalcone from Chromolaena odorata possesses anti-inflammatory effects via inhibition of NF-κB and p38 MAPK in lipopolysaccharide-activated RAW 264.7 macrophages. Immunopharmacol. Immunotoxicol. 2018, 40, 43–51. [Google Scholar] [CrossRef]
- Oh, H.J.; Magar, T.B.T.; Pun, N.T.; Lee, Y.; Kim, E.H.; Lee, E.-S.; Park, P.-H. YJI-7 Suppresses ROS Production and Expression of Inflammatory Mediators via Modulation of p38MAPK and JNK Signaling in RAW 264.7 Macrophages. Biomol. Ther. 2018, 26, 191–200. [Google Scholar] [CrossRef]
- Roh, E.; Lee, H.S.; Kwak, J.A.; Hong, J.T.; Nam, S.Y.; Jung, S.H.; Lee, J.Y.; Kim, N.D.; Han, S.B.; Kim, Y. MD-2 as the target of nonlipid chalcone in the inhibition of endotoxin LPS-induced TLR4 activity. J. Infect. Dis. 2011, 203, 1012–1020. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Assay ID | NCBI Reference Sequence |
|---|---|---|
| Inducible nitric oxide synthase (iNOS) | Mm00440502_m1 | NM 010927.3 |
| Cyclooxygenase-2 (COX-2) | Mm00478374_m1 | NM_011198 |
| β-actin | Mm00607939_s1 | NM_007393.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, Y.C.; Lee, A.; Ryuk, J.A.; Hwang, Y.-H. Isodorsmanin A Prevents Inflammatory Response in LPS-Stimulated Macrophages by Inhibiting the JNK and NF-κB Signaling Pathways. Curr. Issues Mol. Biol. 2023, 45, 1601-1612. https://doi.org/10.3390/cimb45020103
Chung YC, Lee A, Ryuk JA, Hwang Y-H. Isodorsmanin A Prevents Inflammatory Response in LPS-Stimulated Macrophages by Inhibiting the JNK and NF-κB Signaling Pathways. Current Issues in Molecular Biology. 2023; 45(2):1601-1612. https://doi.org/10.3390/cimb45020103
Chicago/Turabian StyleChung, You Chul, Ami Lee, Jin Ah Ryuk, and Youn-Hwan Hwang. 2023. "Isodorsmanin A Prevents Inflammatory Response in LPS-Stimulated Macrophages by Inhibiting the JNK and NF-κB Signaling Pathways" Current Issues in Molecular Biology 45, no. 2: 1601-1612. https://doi.org/10.3390/cimb45020103