
Analysis of the Properties of 44 ABC Transporter Genes from Biocontrol Agent Trichoderma asperellum ACCC30536 and Their Responses to Pathogenic Alternaria alternata Toxin Stress
Abstract
:1. Introduction
2. Materials and methods
2.1. Analysis of 44 ABC Transporter Genes in the Trichoderma asperellum CBS433.97 Genome
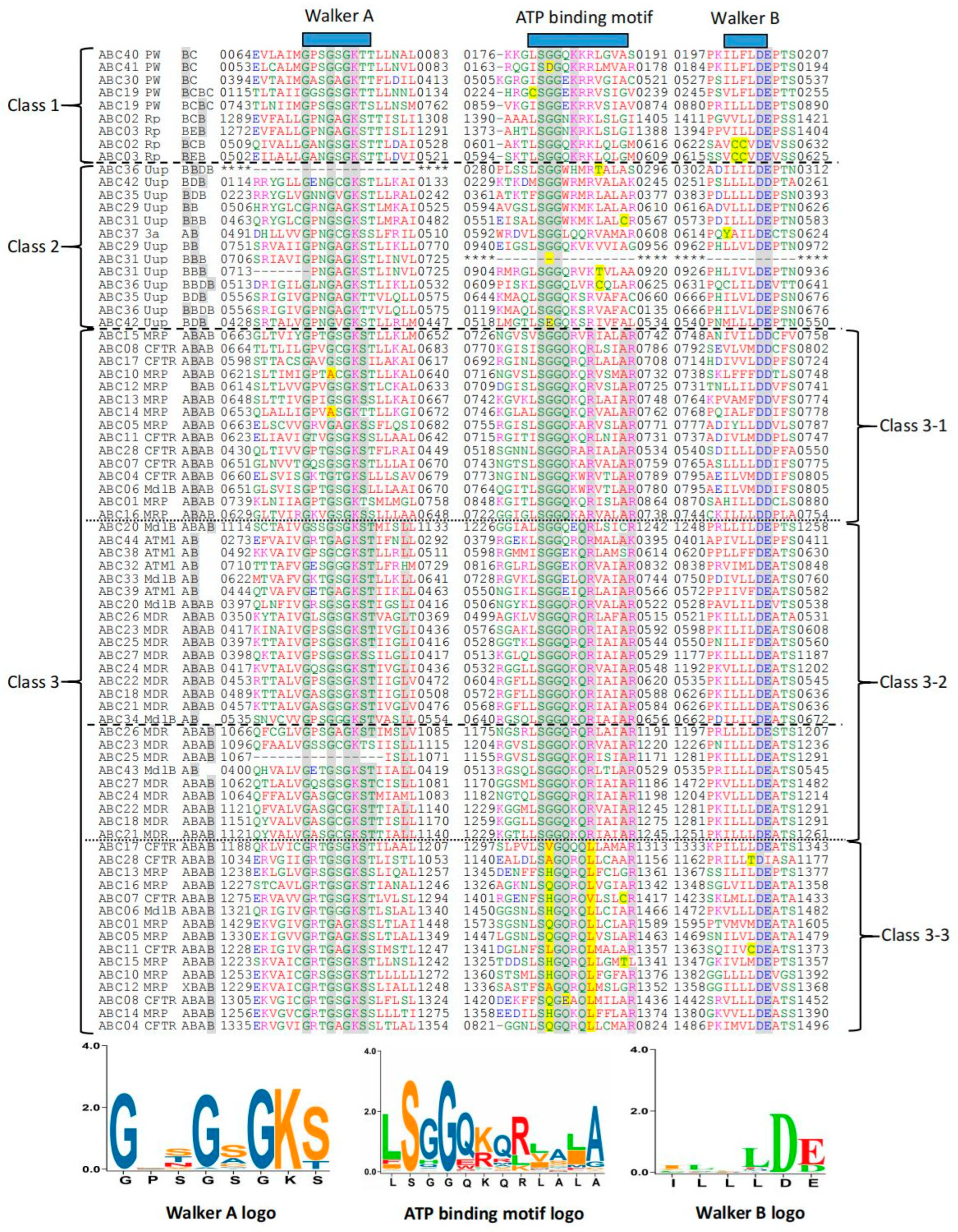
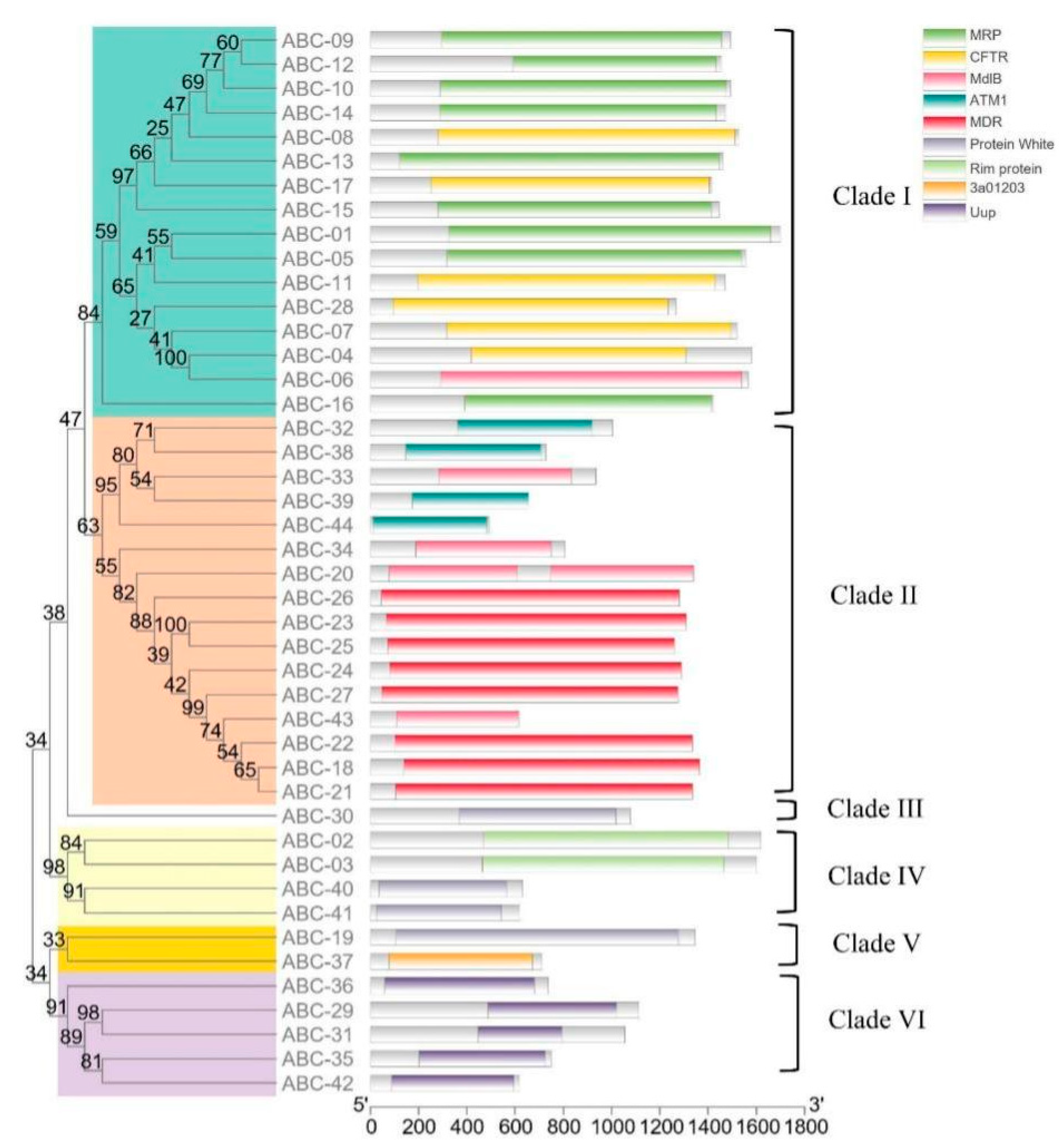
2.2. Multiple Sequence Alignment of 79 NBDs from the 44 ABCs of T. asperellum CBS433.97 and Phylogenetic Analysis of 44 ABCs
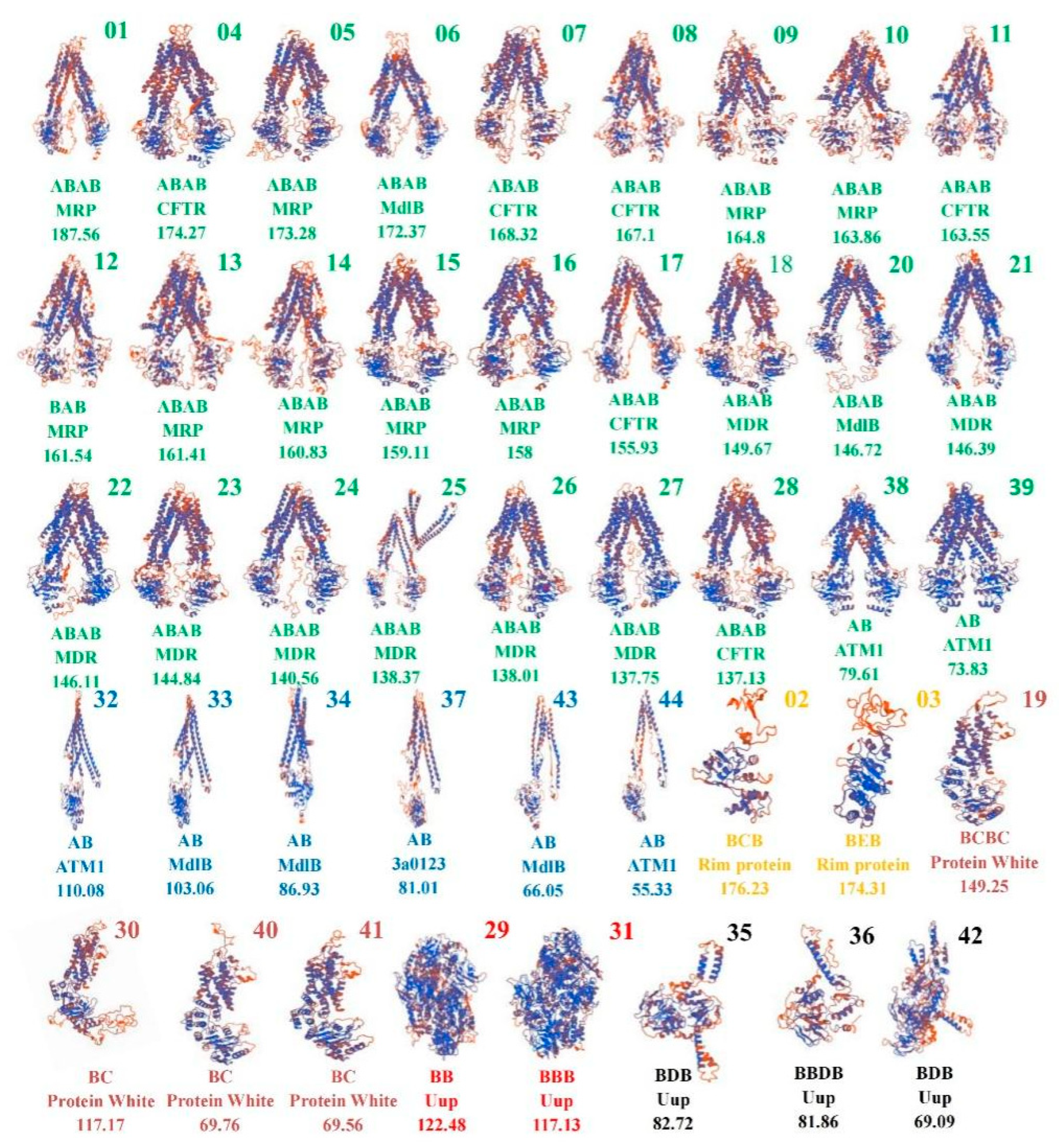
2.3. Prediction of 3D Structure and Promoter of 44 ABCs in T. asperellum CBS433.97
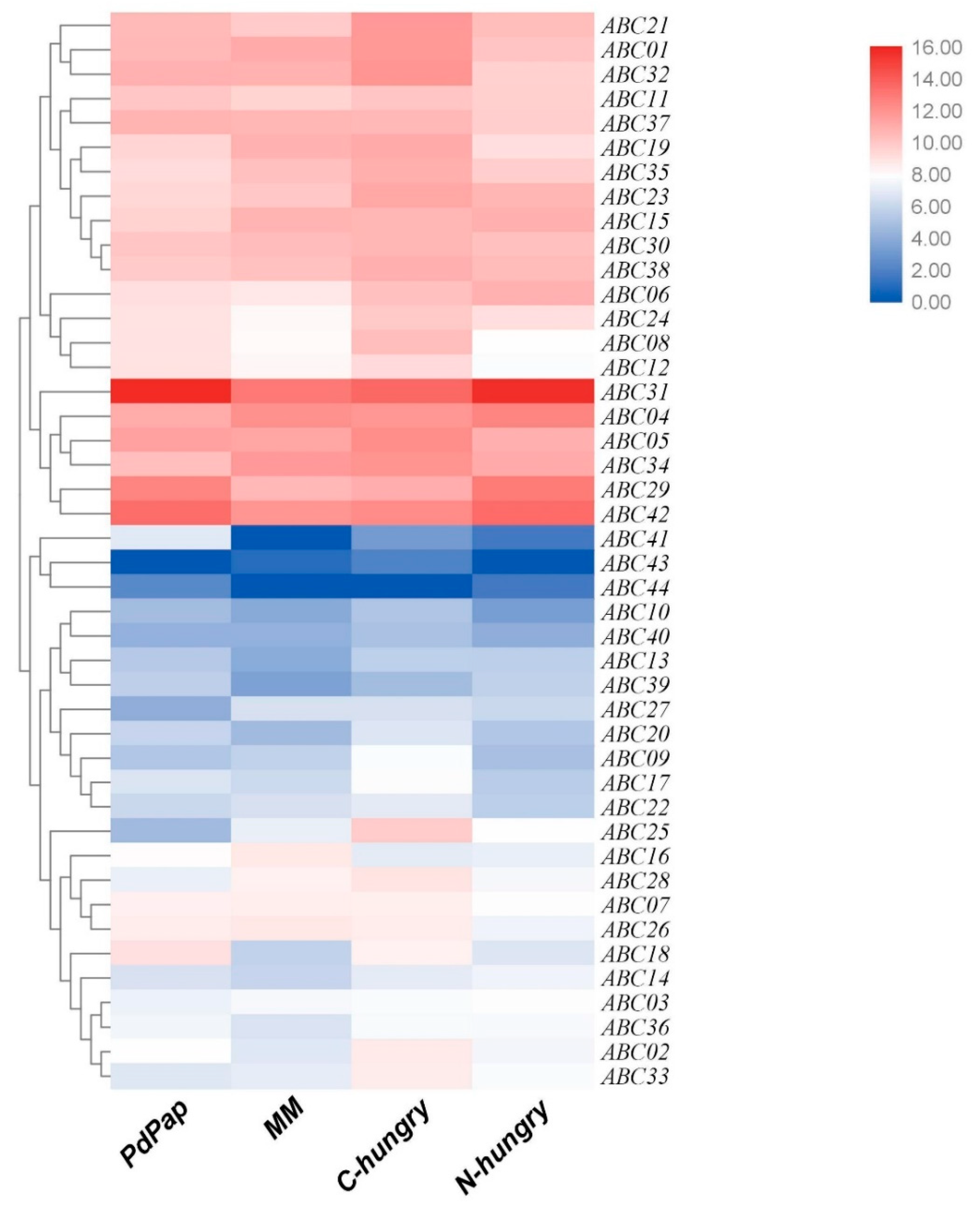
2.4. The Expression Analysis of ABC Transporter Genes under Different Media Based on RNA-Seq
2.5. Transcription Analysis of Six ABCs in Response to Toxins of Alternaria alternata CFCC82114
3. Results
3.1. Analysis of the 44 ABC Transporters in T. asperellum CBS433.97
3.2. Multiple Sequence Alignment of the 79 NBDs Present in the 44 ABCs in T. asperellum CBS433.97
3.3. Phylogenetic Analysis of the 44 ABC Proteins
3.4. Prediction of the 3D Structures of the 44 ABC Transporters in T. asperellum CBS433.97
3.5. Analysis of the Promoters of the 44 ABC Transporter Genes in T. asperellum CBS433.97
3.6. The Expression of 44 ABC Transporter Genes in T. asperellum ACCC30536 under Four Different Conditions
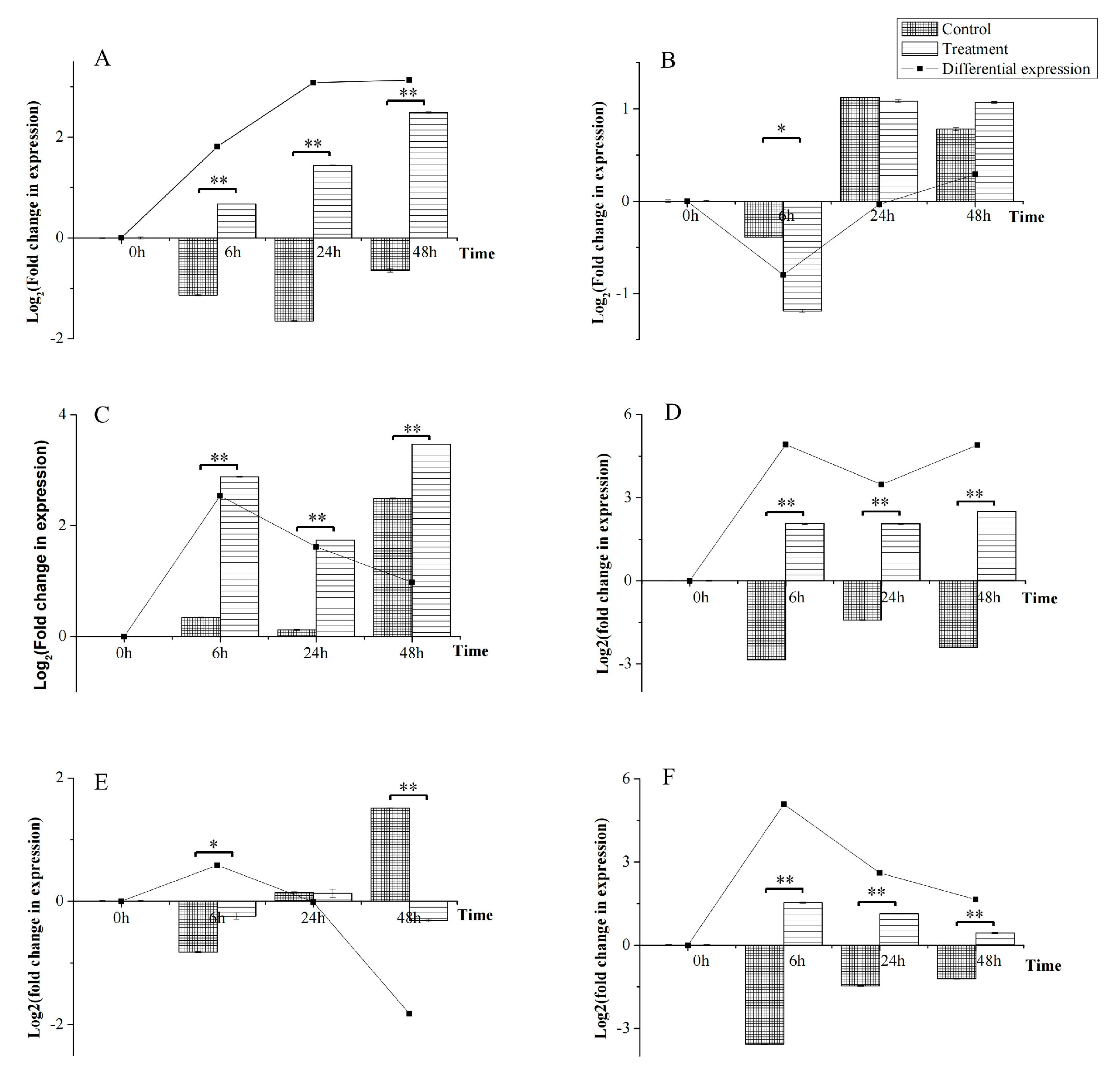
3.7. Differential Expression of Six ABCs in Response to A. alternata CFCC82114 Toxins
4. Discussion
5. Disclosure
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Elad, Y. Biocontrol of foliar pathogens: Mechanisms and application. Commun. Agric. Appl. Biol. Sci. 2003, 68 Pt A, 17–24. [Google Scholar]
- Zhou, C.; Guo, R.; Ji, S.; Fan, H.; Wang, J.; Wang, Y.; Liu, Z. Isolation of Trichoderma from forestry model base and the antifungal properties of isolate TpsT17 toward Fusarium oxysporum. Microbiol. Res. 2020, 231, 126371. [Google Scholar] [CrossRef] [PubMed]
- Bal, U.; Altintas, S. Application of the antagonistic fungus Trichoderma harzianum (TrichoFlow WP™) to root zone increases yield of bell peppers grown in soil. Biol. Agric Hortic. 2006, 24, 149–163. [Google Scholar] [CrossRef]
- Patel, S.; Saraf, M. Biocontrol efficacy of Trichoderma asperellum MSST against tomato wilting by Fusarium oxysporum f. sp. lycopersici. Arch. Phytopathol. Plant Prot. 2017, 50, 228–238. [Google Scholar] [CrossRef]
- Gauthier, C.; Weber, S.; Alarco, A.M.; Alqawi, O.; Daoud, R.; Georges, E.; Raymond, M. Functional similarities and differences between Candida albicans Cdr1p and Cdr2p transporters. Antimicrob. Agents Chemother. 2003, 47, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Lennarz, W.; Lane, M. Encyclopedia of Biological Chemistry, 2nd ed.; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Adamska, A.; Falasca, M. ATP-binding cassette transporters in progression and clinical outcome of pancreatic cancer: What is the way forward? World J. Gastroenterol. 2018, 24, 3225–3238. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Bio. 2009, 10, 218–227. [Google Scholar] [CrossRef]
- Schulz, B.; Kolukisaoglu, H.Ü. Genomics of plant ABC transporters: The alphabet of photosynthetic life forms or just holes in membranes? FEBS Lett. 2006, 580, 1010–1016. [Google Scholar] [CrossRef]
- Borst, P.; Elferink, R.O. Mammalian ABC transporters in health and disease. Annu. Rev. Biochem. 2002, 71, 537–592. [Google Scholar] [CrossRef]
- Bhubhanil, S.; Sittipo, P.; Chaoprasid, P.; Nookabkaew, S.; Sukchawalit, R.; Mongkolsuk, S. Control of zinc homeostasis in Agrobacterium tumefaciens via zur and the zinc uptake genes znuABC and zinT. Microbiology 2014, 160, 2452–2463. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.C.; Sorbo, G.D.; Nistelrooy, J.G.M.V.; Waard, M.A.D. The ABC transporter AtrB from Aspergillus nidulans mediates resistance to all major classes of fungicides and some natural toxic compounds. Microbiology 2000, 146, 1987–1997. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.; Bhargava, T.; Hamer, J.E. An ATP-driven efflux pump is a novel pathogenicity factor in rice blast disease. EMBO J. 1999, 18, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Stefanato, F.L.; Abou-Mansour, E.; Buchala, A.; Kretschmer, M.; Mosbach, A.; Hahn, M.; Bochet, C.G.; Métraux, J.P.; Schoonbeek, H.J. The ABC transporter BcatrB from Botrytis cinerea exports camalexin and is a virulence factor on Arabidopsis thaliana. Plant J. 2009, 58, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Liu, L.; Huang, X.; Li, Y.; Chen, Y.; Chen, J. Proteomic analysis of Trichoderma atroviride mycelia stressed by organophosphate pesticide dichlorvos. Can. J. Microbiol. 2010, 56, 121–127. [Google Scholar] [CrossRef]
- Sharma, P.K.; Gothalwal, R. Trichoderma: A Potent Fungus as Biological Control Agent, 1st ed.; Springer International Publishing AG: New York, NY, USA, 2017; pp. 113–125. [Google Scholar] [CrossRef]
- Wang, C.; Jing, R.; Mao, X.; Chang, X.; Li, A. TaABC1, a member of the activity of bc1 complex protein kinase family from common wheat, confers enhanced tolerance to abiotic stresses in Arabidopsis. J. Exp. Bot. 2011, 62, 1299–1311. [Google Scholar] [CrossRef]
- Ji, S.; Wang, Z.; Wang, J.; Fan, H.; Wang, Y.; Liu, Z. Properties analysis of transcription factor gene TasMYB36 from Trichoderma asperellum CBS433.97 and its heterogeneous transfomation to improve antifungal ability of Populus. Sci. Rep. 2017, 7, 12801. [Google Scholar] [CrossRef]
- Fan, H.; Liu, Z.; Zhang, R.; Wang, N.; Dou, K.; Mijiti, G.; Diao, G.; Wang, Z. Functional analysis of a subtilisin-like serine protease gene from biocontrol fungus Trichoderma harzianum. J. Microbiol. 2014, 52, 129–138. [Google Scholar] [CrossRef]
- Shentu, X.P.; Liu, W.P.; Zhan, X.H.; Xu, Y.P.; Xu, J.F.; Yu, X.P.; Zhang, C.X. Transcriptome sequencing and gene expression analysis of Trichoderma brevicompactum under different culture conditions. PLoS ONE 2014, 9, e94203. [Google Scholar] [CrossRef]
- Ponte-Sucre, A. ABC Transporters in Microorganisms: Research, Innovation and Value as Targets Against Drug Resistance, 1st ed.; Caister Academic Press: Poole, UK, 2009. [Google Scholar]
- Nieves-Morión, M.; Flores, E. Multiple ABC glucoside transporters mediate sugar-stimulated growth in the heterocyst-forming cyanobacterium Anabaena sp. strain PCC 7120. Environ. Microbiol. Rep. 2018, 10, 40–48. [Google Scholar] [CrossRef]
- Szewczyk, P.; Tao, H.; McGrath, A.P.; Villaluz, M.; Rees, S.D.; Lee, S.C.; Doshi, R.; Urbatsch, I.L.; Zhang, Q.; Chang, G. Snapshots of ligand entry, malleable binding and induced helical movement in P-glycoprotein. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Linton, K.J.; Holland, I.B. The ABC Transporters of Human Physiology and Disease: Genetics and Biochemistry of ATP Binding Cassette Transporters, 1st ed.; World Scientifc Publishing Co Pte Ltd: Singapore, 2011. [Google Scholar] [CrossRef]
- Ose, T.; Fujie, T.; Yao, M.; Watanabe, N.; Tanaka, I. Crystal structure of the ATP—Binding cassette of multisugar transporter from Pyrococcus horikoshii OT3. Proteins Struct. Funct. Bioinform. 2004, 57, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kinch, L.N.; Borek, D.M.; Wang, J.; Wang, J.; Urbatsch, I.L.; Xie, X.S.; Grishin, N.V.; Cohen, J.C.; Otwinowski, Z.; et al. Crystal structure of the human sterol transporter ABCG5/ABCG8. Nature 2016, 533, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Boël, G.; Smith, P.C.; Ning, W.; Englander, M.T.; Chen, B.; Hashem, Y.; Testa, A.J.; Fischer, J.J.; Wieden, H.J.; Frank, J.; et al. The ABC-F protein EttA gates ribosome entry into the translation elongation cycle. Nat. Struct. Mol. Biol. 2014, 21, 143–151. [Google Scholar] [CrossRef]
- Andersen, C.B.; Becker, T.; Blau, M.; Anand, M.; Halic, M.; Balar, B.; Mielke, T.; Boesen, T.; Pedersen, J.S.; Spahn, C.M.; et al. Structure of eEF3 and the mechanism of transfer RNA release from the E-site. Nature 2006, 443, 663–668. [Google Scholar] [CrossRef]
- Barthelme, D.; Dinkelaker, S.; Albers, S.V.; Londei, P.; Ermler, U.; Tampé, R. Ribosome recycling depends on a mechanistic link between the FeS cluster domain and a conformational switch of the twin-ATPase ABCE1. Proc. Natl. Acad. Sci. USA 2011, 108, 3228–3233. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Yang, H. Classification and Identification of Trichoderma, 1st ed.; China Land Press: Beijing, China, 2009; pp. 4–5. (In Chinese) [Google Scholar]
- Olano, C.; Rodriguez, A.M.; Méndez, C.; Salas, J.A. A second ABC transporter is involved in oleandomycin resistance and its secretion by Streptomyces antibioticus. Mol. Microbiol. 1995, 16, 333–343. [Google Scholar] [CrossRef]
- Zepeda, M.Y.B.; Alessandri, K.; Murat, D.; Amri, C.E.; Dassa, E. C-terminal domain of the Uup ATP-binding cassette ATPase is an essential folding domain that binds to DNA. Biochim. Biophys. Acta 2009, 1804, 755–761. [Google Scholar] [CrossRef]
- Murat, D.; Bance, P.; Callebaut, I.; Dassa, E. ATP hydrolysis is essential for the function of the Uup ATP binding cassette ATPase in precise excision of transposons. J. Biol. Chem. 2006, 281, 6850–6859. [Google Scholar] [CrossRef]
- Chitsaz, M.; Brown, M.H. The role played by drug efflux pumps in bacterial multidrug resistance. Essays Biochem. 2017, 61, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Venter, H.; Mowla, R.; Ohene-Agyei, T.; Ma, S. RND-type drug efflux pumps from Gram-negative bacteria: Molecular mechanism and inhibition. Front. Microbiol. 2015, 6, 377–386. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Name | Sequence (5′–3′) | Tm (°C) | Size of Product (bp) |
|---|---|---|---|---|
| ABC01 | ABC1-L | TGCTGAAGAAGCCAACCGTCAT | 58.8 | 250 |
| ABC1-R | CTTGTCTCGCACATGCTCCTGA | 59.0 | ||
| ABC02 | ABC2-L | GCTGATCCGAGGCGATCTGAAG | 59.2 | 248 |
| ABC2-R | TGCGTGTGGAGAAGGAACTGAG | 54.5 | ||
| ABC04 | ABC4-L | CTGCTTCAACCGCCTCTCCAA | 58.8 | 231 |
| ABC4-R | GCGTGCTGTCAGTGAACTCCT | 58.8 | ||
| ABC05 | ABC5-L | GACACTGGCTCTGTTCCGACTC | 59.0 | 246 |
| ABC5-R | CCACCATCCATGCTGGCTACAT | 58.7 | ||
| ABC30 | ABC30-L | CCACCAACCTCGCTCCAACAT | 58.8 | 221 |
| ABC30-R | CGTCGGCATCAATTCGTCCATC | 58.5 | ||
| ABC31 | ABC31-L | AGCGTCGTGTCATTGGCATCA | 58.9 | 258 |
| ABC31-R | CGCAGTGGCTCTCAATCTCCTT | 58.7 | ||
| α-tubulin | α-tu-L | CACATGGTTGACTGGTGCCCTA | 58.6 | 240 |
| α-tu-R | CTCGCCCTCTTCCATACCCTCT | 59.0 | ||
| β-tubulin | β-tu-L | TTCTTCCACCTTTGTCGGCAACTCCT | 59.1 | 227 |
| β-tu-R | CCTCGTACTCCTCACCATCA | 59.1 | ||
| actin | Act-L | GGCTCAGTCTAAGCGTGGTATCC | 59.0 | 251 |
| Act-R | ACAGAACGGCCTGGATGGAGA | 59.0 |
| Three-Dimensional Structure | Multidomain | Conserved Domain Arrangement Order |
|---|---|---|
| Structural type 1 | MRP, CFTR, MDR, MdlB | A-B-A-B |
| ATM1 | A-B | |
| Structural type 2 | ATM1, MdlB, 3a0123 | A-B |
| Structural type 3 | Rim protein | B-C-B, B-E-B |
| Structural type 4 | Protein White | B-C-B-C, B-C |
| Structural type 5 | Uup | B-B, B-B-B |
| Structural type 6 | Uup | B-D-B, B-B-D-B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, H.-Y.; Zhang, Y.-Z.; Liu, K.; Gu, P.-W.; Cao, S.; Gao, X.; Wang, Z.-Y.; Liu, Z.-H.; Yu, Z.-Y. Analysis of the Properties of 44 ABC Transporter Genes from Biocontrol Agent Trichoderma asperellum ACCC30536 and Their Responses to Pathogenic Alternaria alternata Toxin Stress. Curr. Issues Mol. Biol. 2023, 45, 1570-1586. https://doi.org/10.3390/cimb45020101
Du H-Y, Zhang Y-Z, Liu K, Gu P-W, Cao S, Gao X, Wang Z-Y, Liu Z-H, Yu Z-Y. Analysis of the Properties of 44 ABC Transporter Genes from Biocontrol Agent Trichoderma asperellum ACCC30536 and Their Responses to Pathogenic Alternaria alternata Toxin Stress. Current Issues in Molecular Biology. 2023; 45(2):1570-1586. https://doi.org/10.3390/cimb45020101
Chicago/Turabian StyleDu, Hua-Ying, Yu-Zhou Zhang, Kuo Liu, Pei-Wen Gu, Shuang Cao, Xiang Gao, Zhi-Ying Wang, Zhi-Hua Liu, and Ze-Yang Yu. 2023. "Analysis of the Properties of 44 ABC Transporter Genes from Biocontrol Agent Trichoderma asperellum ACCC30536 and Their Responses to Pathogenic Alternaria alternata Toxin Stress" Current Issues in Molecular Biology 45, no. 2: 1570-1586. https://doi.org/10.3390/cimb45020101