
SRY-Related Transcription Factors in Head and Neck Squamous Cell Carcinomas: In Silico Based Analysis
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. TGCA Data
2.2. Data Analysis
2.3. Gene Analysis
2.4. Association of SRY-Related Transcripts in Context to Stromal and Immune Scores and Immune Profile
2.5. Validation of the Results
2.6. Statistical Analysis
3. Results
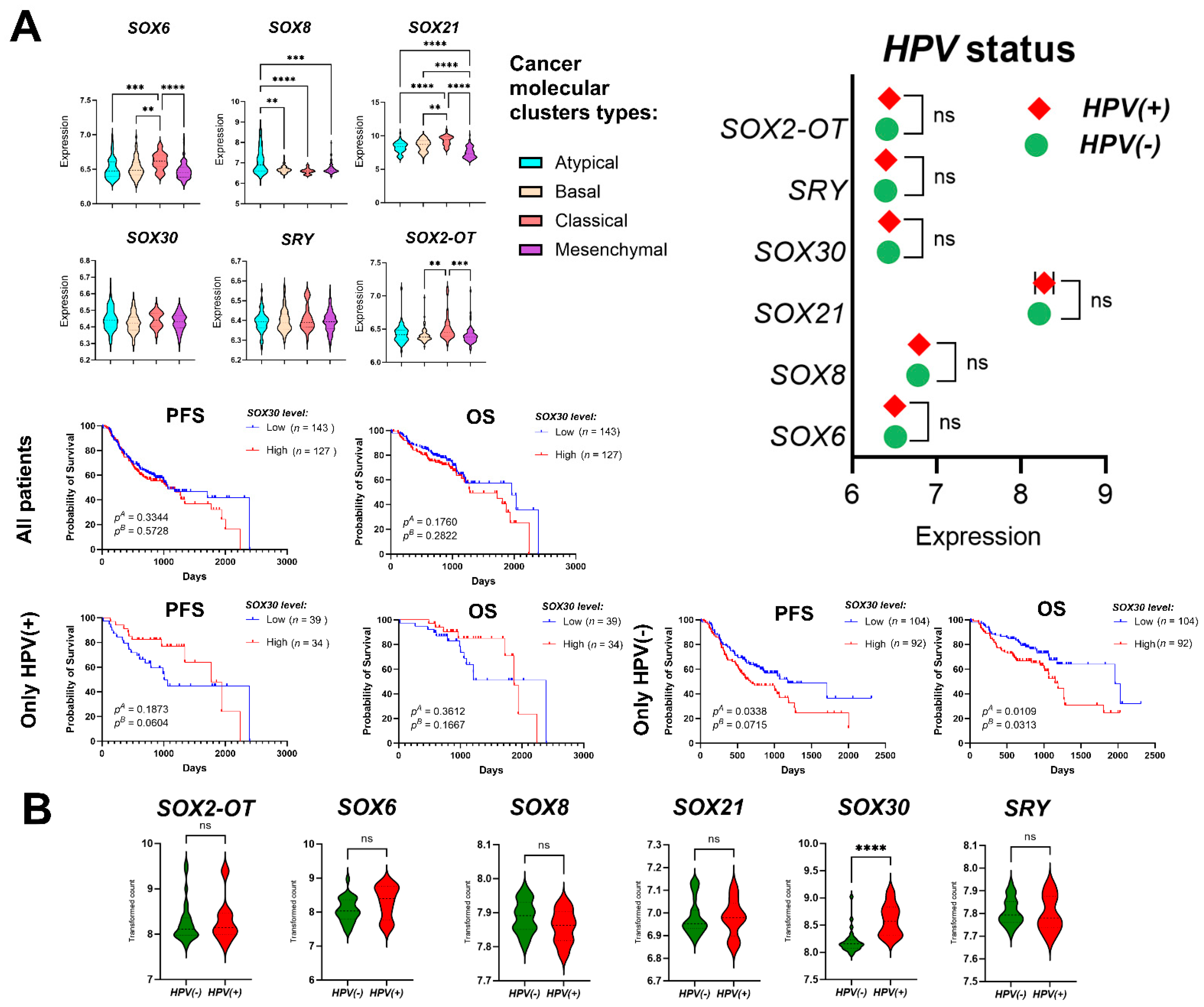
3.1. SOX Family Gene Expression Levels Distinguish between Healthy and Cancer Tissues
3.2. Expression Levels of SRY-Related Transcription Factors Depend on Tumor Localization
3.3. Expression Levels of SRY-Related Transcription Factors Depend on Specific Clinicopathological Parameters
3.4. Higher Survival Rates Are Associated with High SOX21 and SRY Expression Levels
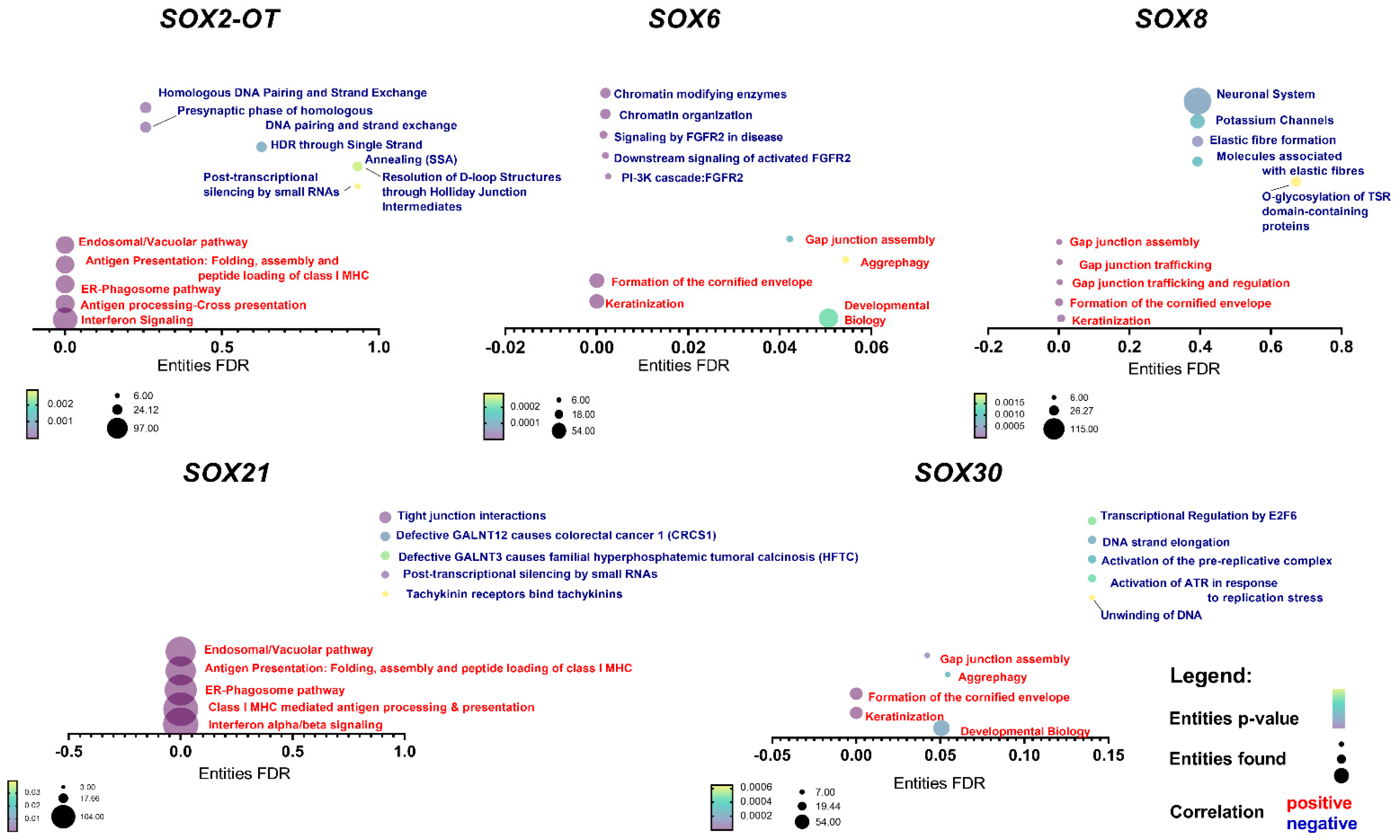
3.5. SOX Genes’ Expression Levels Correlate with Important Cellular Processes
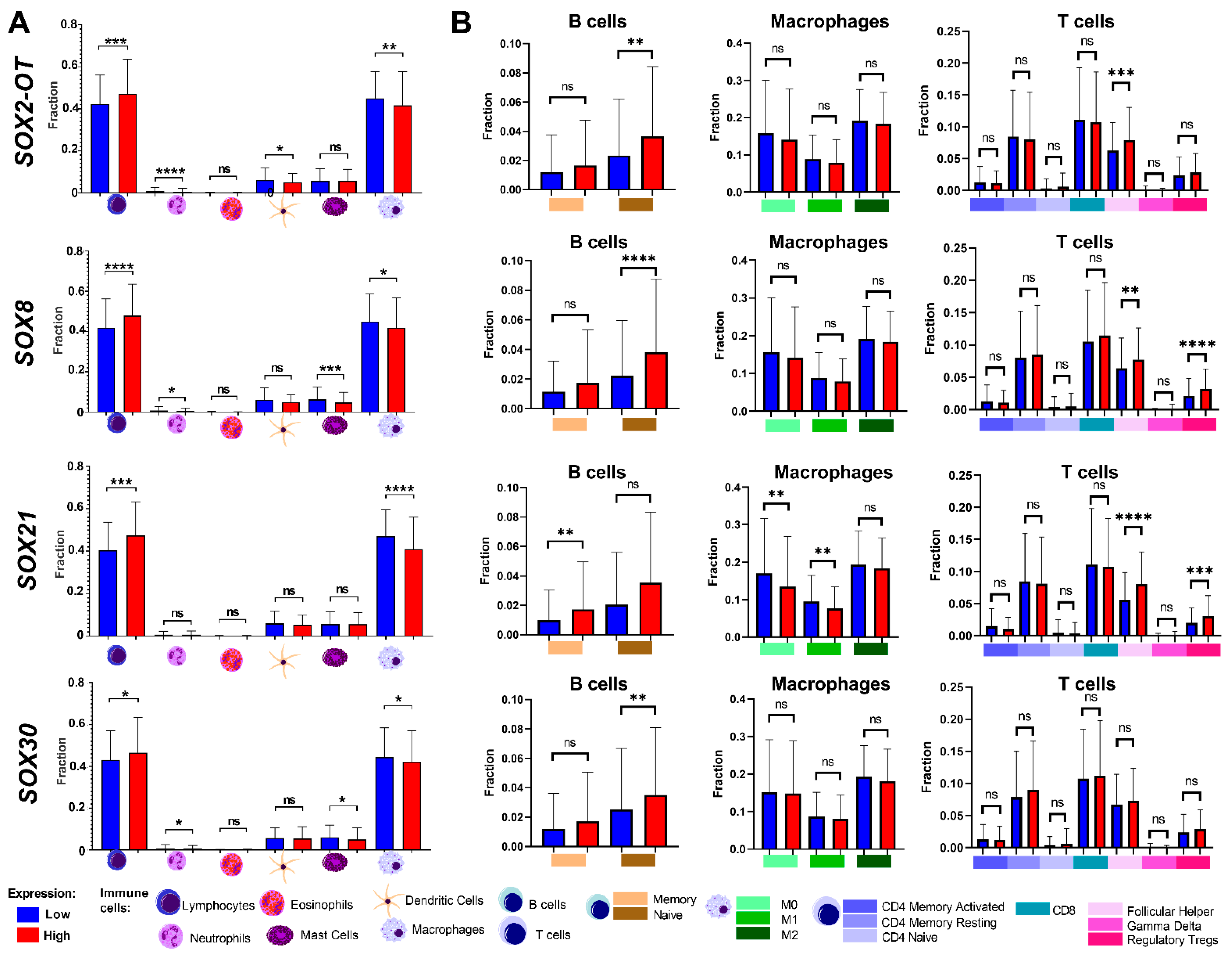
3.6. Tumor Composition Is Associated with Differences in the Expression of SRY-Related Transcription Factors
3.7. Validation of the Results for SRY-Related Transcription Factors Using the GEO Datasets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marur, S.; Forastiere, A.A. Head and neck cancer: Changing epidemiology, diagnosis, and treatment. Mayo Clin. Proc. 2008, 83, 489–501. [Google Scholar] [CrossRef]
- Dyzmann-Sroka, A.; Malicki, J.; Jędrzejczak, A. Cancer incidence in the Greater Poland region as compared to Europe. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2020, 25, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Braakhuis, B.J.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Dok, R.; Nuyts, S. HPV Positive Head and Neck Cancers: Molecular Pathogenesis and Evolving Treatment Strategies. Cancers 2016, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical update on head and neck cancer: Molecular biology and ongoing challenges. Cell Death Dis. 2019, 10, 540. [Google Scholar] [CrossRef] [PubMed]
- Rupar, M.J.; Golusinski, P.; Golusinski, W.; Masternak, M.M. Human Papillomavirus and the use of nanoparticles for immunotherapy in HPV-related cancer: A review. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2019, 24, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Rajabi-Moghaddam, M.; Abbaszadeh, H. Gene polymorphisms and prognosis of head and neck squamous cell carcinoma: A systematic review. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2022, 27, 1045–1057. [Google Scholar] [CrossRef]
- Rajabi-Moghaddam, M.; Abbaszadeh, H. Gene polymorphisms and risk of head and neck squamous cell carcinoma: A systematic review. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2022, 27, 1058–1076. [Google Scholar] [CrossRef]
- Guglas, K.; Kozłowska-Masłoń, J.; Kolenda, T.; Paszkowska, A.; Teresiak, A.; Bliźniak, R.; Lamperska, K. Midsize noncoding RNAs in cancers: A new division that clarifies the world of noncoding RNA or an unnecessary chaos? Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2022, 27, 1077–1093. [Google Scholar] [CrossRef]
- Kolenda, T.; Guglas, K.; Ryś, M.; Bogaczyńska, M.; Teresiak, A.; Bliźniak, R.; Łasińska, I.; Mackiewicz, J.; Lamperska, K.M. Biological role of long non-coding RNA in head and neck cancers. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2017, 22, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, T.; Guglas, K.; Kopczyńska, M.; Sobocińska, J.; Teresiak, A.; Bliźniak, R.; Lamperska, K. Good or not good: Role of miR-18a in cancer biology. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2020, 25, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, T.; Paszkowska, A.; Braska, A.; Kozłowska-Masłoń, J.; Guglas, K.; Poter, P.; Wojtczak, P.; Bliźniak, R.; Lamperska, K.; Teresiak, A. Host gene and its guest: Short story about relation of long-noncoding MIR31HG transcript and microRNA miR-31. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2023, 28, 114–134. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, T.; Kopczyńska, M.; Guglas, K.; Teresiak, A.; Bliźniak, R.; Łasińska, I.; Mackiewicz, J.; Lamperska, K. EGOT lncRNA in head and neck squamous cell carcinomas. Pol. J. Pathol. Off. J. Pol. Soc. Pathol. 2018, 69, 356–365. [Google Scholar] [CrossRef]
- Kopczyńska, M.; Kolenda, T.; Guglas, K.; Sobocińska, J.; Teresiak, A.; Bliźniak, R.; Mackiewicz, A.; Mackiewicz, J.; Lamperska, K. PRINS lncRNA Is a New Biomarker Candidate for HPV Infection and Prognosis of Head and Neck Squamous Cell Carcinomas. Diagnostics 2020, 10, 762. [Google Scholar] [CrossRef]
- Kolenda, T.; Guglas, K.; Baranowski, D.; Sobocińska, J.; Kopczyńska, M.; Teresiak, A.; Bliźniak, R.; Lamperska, K. cfRNAs as biomarkers in oncology—Still experimental or applied tool for personalized medicine already? Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2020, 25, 783–792. [Google Scholar] [CrossRef]
- Kozłowska, J.; Kolenda, T.; Poter, P.; Sobocińska, J.; Guglas, K.; Stasiak, M.; Bliźniak, R.; Teresiak, A.; Lamperska, K. Long Intergenic Non-Coding RNAs in HNSCC: From “Junk DNA” to Important Prognostic Factor. Cancers 2021, 13, 2949. [Google Scholar] [CrossRef]
- Lv, L.; Zhou, M.; Zhang, J.; Liu, F.; Qi, L.; Zhang, S.; Bi, Y.; Yu, Y. SOX6 suppresses the development of lung adenocarcinoma by regulating expression of p53, p21CIPI, cyclin D1 and β-catenin. FEBS Open Bio 2020, 10, 135–146. [Google Scholar] [CrossRef]
- Liang, Z.; Xu, J.; Gu, C. Novel role of the SRY-related high-mobility-group box D gene in cancer. Semin. Cancer Biol. 2020, 67 Pt 1, 83–90. [Google Scholar] [CrossRef]
- Xie, S.L.; Fan, S.; Zhang, S.Y.; Chen, W.X.; Li, Q.X.; Pan, G.K.; Zhang, H.Q.; Wang, W.W.; Weng, B.; Zhang, Z.; et al. SOX8 regulates cancer stem-like properties and cisplatin-induced EMT in tongue squamous cell carcinoma by acting on the Wnt/β-catenin pathway. Int. J. Cancer 2018, 142, 1252–1265. [Google Scholar] [CrossRef] [PubMed]
- Shahryari, A.; Jazi, M.S.; Samaei, N.M.; Mowla, S.J. Long non-coding RNA SOX2OT: Expression signature, splicing patterns, and emerging roles in pluripotency and tumorigenesis. Front. Genet. 2015, 6, 196. [Google Scholar] [CrossRef]
- Staniewska, E.; Tomasik, B.; Tarnawski, R.; Łaszczych, M.; Miszczyk, M. The prognostic value of red cell distribution width (RDW), neutrophil-to-lymphocyte ratio (NLR), and platelet-to-lymphocyte ratio (PLR) in radiotherapy for oropharyngeal cancer. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2021, 26, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, M.S.; Narayanan, G.S.; Kumar, R. Change of telomerase activity in peripheral blood of patients with head and neck squamous cell carcinoma pre and post curative treatment. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2020, 25, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, A.K.; Kazmierska, J.; Marszalek, A.; Golusinski, P.; Heydrych, A.; Wiechec, K.; Cholewinski, W. Dual-time-point PET/CT study protocol can improve the larynx cancer diagnosis. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2020, 25, 533–538. [Google Scholar] [CrossRef]
- Sindhu, S.K.; Bauman, J.E. Current Concepts in Chemotherapy for Head and Neck Cancer. Oral Maxillofac. Surg. Clin. N. Am. 2019, 31, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.; Gulidov, I.; Semenov, A.; Golovanova, O.; Koryakin, S.; Makeenkova, T.; Ivanov, S.; Kaprin, A. Proton re-irradiation of unresectable recurrent head and neck cancers. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2021, 26, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Koiwai, K.; Hirasawa, D.; Sugimura, M.; Endo, Y.; Mizuhata, K.; Ina, H.; Fukazawa, A.; Kitoh, R.; Sakai, H.; Fujinaga, Y. Impact of upgraded radiotherapy system on outcomes in postoperative head and neck squamous cell carcinoma patients. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2022, 27, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Viani, G.A.; Faustino, A.C.; Danelichen AF, B.; Matsuura, F.K.; Neves LV, F.; Fernandes, M.H.; Fernandes, J.P. Radiotherapy for locally advanced head and neck cancer in elderly patients: Results and prognostic factors a single cohort. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2021, 26, 12–19. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Sobocińska, J.; Nowakowska, J.; Molenda, S.; Olechnowicz, A.; Guglas, K.; Kozłowska-Masłoń, J.; Kazimierczak, U.; Machnik, M.; Oleksiewicz, U.; Teresiak, A.; et al. Zinc Finger Proteins in Head and Neck Squamous Cell Carcinomas: ZNF540 May Serve as a Biomarker. Curr. Oncol. (Tor. Ont.) 2022, 29, 9896–9915. [Google Scholar] [CrossRef]
- Tomaszewska, W.; Kozłowska-Masłoń, J.; Baranowski, D.; Perkowska, A.; Szałkowska, S.; Kazimierczak, U.; Severino, P.; Lamperska, K.; Kolenda, T. miR-154 Influences HNSCC Development and Progression through Regulation of the Epithelial-to-Mesenchymal Transition Process and Could Be Used as a Potential Biomarker. Biomedicines 2021, 9, 1894. [Google Scholar] [CrossRef]
- Koteluk, O.; Bielicka, A.; Lemańska, Ż.; Jóźwiak, K.; Klawiter, W.; Mackiewicz, A.; Kazimierczak, U.; Kolenda, T. The Landscape of Transmembrane Protein Family Members in Head and Neck Cancers: Their Biological Role and Diagnostic Utility. Cancers 2021, 13, 4737. [Google Scholar] [CrossRef]
- Paszkowska, A.; Kolenda, T.; Guglas, K.; Kozłowska-Masłoń, J.; Podralska, M.; Teresiak, A.; Bliźniak, R.; Dzikiewicz-Krawczyk, A.; Lamperska, K. C10orf55, CASC2, and SFTA1P lncRNAs Are Potential Biomarkers to Assess Radiation Therapy Response in Head and Neck Cancers. J. Pers. Med. 2022, 12, 1696. [Google Scholar] [CrossRef]
- Kolenda, T.; Poter, P.; Guglas, K.; Kozłowska-Masłoń, J.; Braska, A.; Kazimierczak, U.; Teresiak, A. Biological role and diagnostic utility of ribosomal protein L23a pseudogene 53 in cutaneous melanoma. Rep. Pract. Oncol. Radiother. J. Greatpoland Cancer Cent. Pozn. Pol. Soc. Radiat. Oncol. 2023, 28, 255–270. [Google Scholar] [CrossRef]
- Wichmann, G.; Rosolowski, M.; Krohn, K.; Kreuz, M.; Boehm, A.; Reiche, A.; Scharrer, U.; Halama, D.; Bertolini, J.; Bauer, U.; et al. The role of HPV RNA transcription, immune response-related gene expression and disruptive TP53 mutations in diagnostic and prognostic profiling of head and neck cancer. Int. J. Cancer 2015, 137, 2846–2857. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.P.; Yuan, Y.J.; Zhu, J.Q.; Li, B.L.; Zhang, T.T. Analysis of key genes and signal pathways of human papilloma virus-related head and neck squamous cell carcinoma. Chin. J. Stomatol. 2020, 55, 571–577. [Google Scholar] [CrossRef]
- Bowles, J.; Schepers, G.; Koopman, P. Phylogeny of the SOX family of developmental transcription factors based on sequence and structural indicators. Dev. Biol. 2000, 227, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Wise, P.; Krüger, M.; Simonsen, U.; Wehland, M.; Infanger, M.; Corydon, T.J. The role of SOX family members in solid tumours and metastasis. Semin. Cancer Biol. 2020, 67 Pt 1, 122–153. [Google Scholar] [CrossRef] [PubMed]
- Pouremamali, F.; Vahedian, V.; Hassani, N.; Mirzaei, S.; Pouremamali, A.; Kazemzadeh, H.; Faridvand, Y.; Jafari-Gharabaghlou, D.; Nouri, M.; Maroufi, N.F. The role of SOX family in cancer stem cell maintenance: With a focus on SOX2. Pathology 2022, 231, 153783. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, N.; Luo, X.; Zhang, X.; Liao, Q.; Wang, J. SOX2OT, a novel tumor-related long non-coding RNA. Biomed. Pharmacother. = Biomed. Pharmacother. 2020, 123, 109725. [Google Scholar] [CrossRef]
- Kumar, P.; Mistri, T.K. Transcription factors in SOX family: Potent regulators for cancer initiation and development in the human body. Semin. Cancer Biol. 2020, 67 Pt 1, 105–113. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, H.; Chen, T.; Zhu, X.; Ye, J.; Lv, K. Identification of HMG-box family establishes the significance of SOX6 in the malignant progression of glioblastoma. Aging 2020, 12, 8084–8106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Niu, X.; Zhang, X.; Zhan, G.; Xue, X.; Wang, X.; Zhang, H.; Guo, Z. SRY-related high-mobility-group box 6 suppresses cell proliferation and is downregulated in breast cancer. Anti-Cancer Drugs 2021, 32, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, H.; Li, X.; Chen, W.; Zhang, X.; Yang, Z.; Chen, Z.; Chen, J.; Zhang, Y.; Shi, D.; et al. High SOX8 expression promotes tumor growth and predicts poor prognosis through GOLPH3 signaling in tongue squamous cell carcinoma. Cancer Med. 2020, 9, 4274–4289. [Google Scholar] [CrossRef]
- Mehta, G.A.; Khanna, P.; Gatza, M.L. Emerging Role of SOX Proteins in Breast Cancer Development and Maintenance. J. Mammary Gland Biol. Neoplasia 2019, 24, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, Y.; Tian, J.; Zhang, S.; Li, X.; Zhai, X.; Feng, Q. High expression of SRY-box transcription factor 30 associates with well differentiation, absent lymph node metastasis and predicts longer survival in nonsmall-cell lung cancer patients. Medicine 2020, 99, e20122. [Google Scholar] [CrossRef]
- Han, F.; Liu, W.B.; Li, J.J.; Zhang, M.Q.; Yang, J.T.; Zhang, X.; Hao, X.L.; Yin, L.; Mao, C.Y.; Jiang, X.; et al. SOX30 is a prognostic biomarker and chemotherapeutic indicator for advanced-stage ovarian cancer. Endocr.-Relat. Cancer 2019, 26, 303–319. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Group | SOX2-OT | SOX6 | SOX8 | SOX21 | SOX30 | SRY | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | ||
| Age | ≤61 | 0.04783 ± 0.1198; n = 281 | 0.7991 | −0.1924 ± 0.1048; n = 281 | 0.2216 | −1.653 ± 0.1087; n = 281 | 0.2873 | 3.078 ± 0.1578; n = 281 | 0.3361 | 0.6262 ± 0.1301; n = 281 | 0.2568 | 0.8572 ± 0.08776; n = 281 | 0.04 |
| >61 | 0.1532 ± 0.1391; n = 240 | −0.3326 ± 0.1070; n = 240 | −1.885 ± 0.09462; n = 240 | 2.780 ± 0.1798; n = 240 | 0.3228 ± 0.1224; n = 240 | 0.6531 ± 0.09133; n = 240 | |||||||
| Gender | Female | 0.04783 ± 0.1198; n = 281 | 0.7991 | −0.1165 ± 0.1309; n = 137 | 0.4837 | −1.964 ± 0.1156; n = 137 | 0.1797 | 2.373 ± 0.2214; n = 137 | 0.0013 | 0.1220 ± 0.1505; n = 137 | 0.0556 | −0.3659 ± 0.005930; n = 137 | <0.0001 |
| Male | 0.1532 ± 0.1391; n = 240 | −0.3071 ± 0.09033; n = 384 | −1.687 ± 0.09008; n = 384 | 3.144 ± 0.1393; n = 384 | 0.6164 ± 0.1093; n = 384 | 1.166 ± 0.07612; n = 384 | |||||||
| Alcohol | Positive | 0.1556 ± 0.1136; n = 348 | 0.5642 | −0.2460 ± 0.09173; n = 348 | 0.7338 | −1.713 ± 0.08719; n = 348 | 0.1459 | 2.857 ± 0.1488; n = 348 | 0.4252 | 0.5789 ± 0.1124; n = 348 | 0.2288 | 0.8373 ± 0.07675; n = 348 | 0.023 |
| Negative | 0.002347 ± 0.1577; n = 163 | −0.2776 ± 0.1351; n = 163 | −1.856 ± 0.1399; n = 163 | 3.109 ± 0.1999; n = 163 | 0.2951 ± 0.1521; n = 163 | 0.6343 ± 0.1169; n = 163 | |||||||
| Smoking | Yes- and Ex-smoker | 0.2712 ± 0.1096; n = 392 | 0.0023 | −0.3185 ± 0.08872; n = 392 | <0.0001 | −1.751 ± 0.08042; n = 392 | < 0.0001 | 3.082 ± 0.1368; n = 392 | <0.0001 | 0.4488 ± 0.1002; n = 392 | 0.1163 | 0.8309 ± 0.07446; n = 392 | 0.0472 |
| Non-smoker | −0.4950 ± 0.1496; n = 117 | −1.805 ± 0.1676; n = 117 | 2.525 ± 0.2500; n = 117 | 0.6635 ± 0.2126; n = 117 | 0.5933 ± 0.1253; n = 117 | 0.5933 ± 0.1253; n = 117 | |||||||
| Parameter | Group | SOX2-OT | SOX6 | SOX8 | SOX21 | SOX30 | SRY | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | Mean ± SEM | p-Value | ||
| Cancer Stage | I + II | 0.2040 ± 0.2092; n = 117 | 0.924 | 0.1695 ± 0.1433; n = 117 | 0.0014 | −1.904 ± 0.1304; n = 117 | 0.5406 | 2.957 ± 0.2369; n = 117 | 0.7 | 0.2561 ± 0.1733; n = 117 | 0.2227 | 0.7084 ± 0.1211; n = 117 | 0.9845 |
| III + IV | 0.06481 ± 0.1033; n = 390 | −0.3372 ± 0.08720; n = 390 | −1.700 ± 0.08909; n = 390 | 2.964 ± 0.1389; n = 390 | 0.5903 ± 0.1072; n = 390 | 0.7934 ± 0.07595; n = 390 | |||||||
| T Stage | T1 + T2 | 0.2248 ± 0.2679; n = 49 | 0.5677 | −0.01565 ± 0.2695; n = 49 | 0.1375 | −1.401 ± 0.2418; n = 49 | 0.0733 | 3.178 ± 0.3608; n = 49 | 0.3988 | 0.4485 ± 0.3039; n = 49 | 0.8232 | 0.9239 ± 0.2115; n = 49 | 0.2598 |
| T3 + T4 | 0.04335 ± 0.1013; n = 410 | −0.3313 ± 0.08443; n = 410 | −1.861 ± 0.07732; n = 410 | 2.832 ± 0.1321; n = 410 | 0.2970 ± 0.09282; n = 410 | 0.6437 ± 0.06757; n = 410 | |||||||
| N Stage | N0 + N1 | −0.1296 ± 0.1123; n = 242 | 0.2572 | −0.2025 ± 0.1081; n = 242 | 0.1625 | −1.845 ± 0.1005; n = 242 | 0.1763 | 2.988 ± 0.1669; n = 242 | 0.5351 | 0.05867 ± 0.1205; n = 242 | 0.0008 | 0.7524 ± 0.09294; n = 242 | 0.4682 |
| N2 + N3 | 0.2947 ± 0.1767; n = 179 | −0.4931 ± 0.1364; n = 179 | −1.685 ± 0.1211; n = 179 | 2.815 ± 0.2017; n = 179 | 0.5726 ± 0.1409; n = 179 | 0.6242 ± 0.09903; n = 179 | |||||||
| Grade | G1 + G2 | 0.01121 ± 0.1074; n = 368 | 0.2242 | −0.2435 ± 0.08779; n = 368 | 0.3276 | −1.940 ± 0.07623; n = 368 | 0.0004 | 2.958 ± 0.1342; n = 368 | 0.8503 | 0.1451 ± 0.09247; n = 368 | <0.0001 | 0.6729 ± 0.07275; n = 368 | 0.1391 |
| G3 + G4 | 0.1540 ± 0.1750; n = 131 | −0.4276 ± 0.1544; n = 131 | −1.361 ± 0.1612; n = 131 | 2.765 ± 0.2660; n = 131 | 1.177 ± 0.2132; n = 131 | 0.9374 ± 0.1368; n = 131 | |||||||
| Perineural invasion | Positive | −0.3446 ± 0.1501; n = 169 | 0.0003 | −0.4608 ± 0.1270; n = 169 | 0.2902 | −1.985 ± 0.08808; n = 169 | 0.7462 | 2.222 ± 0.1991; n = 169 | 0.0007 | 0.1076 ± 0.1265; n = 169 | 0.321 | 0.5787 ± 0.09613; n = 169 | 0.6863 |
| Negative | 0.3188 ± 0.1490; n = 195 | −0.3495 ± 0.1333; n = 195 | −1.720 ± 0.1292; n = 195 | 3.116 ± 0.1936; n = 195 | 0.3887 ± 0.1430; n = 195 | 0.5730 ± 0.09789; n = 195 | |||||||
| Lymph node neck dissection | Positive | −0.01959 ± 0.1016; n = 421 | 0.0018 | −0.3538 ± 0.08515; n = 421 | 0.0165 | −1.811 ± 0.07666; n = 421 | 0.4513 | 2.774 ± 0.1316; n = 421 | 0.0019 | 0.2630 ± 0.09197; n = 421 | <0.0001 | 0.7351 ± 0.06894; n = 421 | 0.6681 |
| Negative | 0.6196 ± 0.2022; n = 97 | 0.1205 ± 0.1519; n = 97 | −1.527 ± 0.2069; n = 97 | 3.620 ± 0.2725; n = 97 | 1.505 ± 0.2486; n = 97 | 0.9131 ± 0.1620; n = 97 | |||||||
| Lymphovascular invasion | Positive | 0.4095 ± 0.2077; n = 125 | 0.0242 | −0.4722 ± 0.1745; n = 125 | 0.7387 | −1.706 ± 0.1411; n = 125 | 0.0449 | 3.139 ± 0.2387; n = 125 | 0.0371 | 0.5804 ± 0.1807; n = 125 | 0.0089 | 0.6793 ± 0.1221; n = 125 | 0.2787 |
| Negative | −0.2250 ± 0.1262; n = 225 | −0.3567 ± 0.1143; n = 225 | −1.973 ± 0.09506; n = 225 | 2.538 ± 0.1816; n = 225 | 0.02504 ± 0.1149; n = 225 | 0.5405 ± 0.08737; n = 225 | |||||||
| HPV in p16 test | Positive | 1.157 ± 0.2662; n = 39 | 0.009 | −0.1204 ± 0.3127; n = 39 | 0.2585 | −0.5596 ± 0.4108; n = 39 | 0.0147 | 4.145 ± 0.3795; n = 39 | 0.0559 | 3.471 ± 0.3312; n = 39 | <0.0001 | 1.423 ± 0.2762; n = 39 | 0.1264 |
| Negative | 0.3251 ± 0.2632; n = 73 | −0.3236 ± 0.2012; n = 73 | −1.633 ± 0.1988; n = 73 | 3.295 ± 0.2976; n = 73 | −0.1825 ± 0.1713; n = 73 | 0.8632 ± 0.1700; n = 73 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolenda, T.; Graczyk, Z.; Żarska, B.; Łosiewski, W.; Smolibowski, M.; Wartecki, A.; Kozłowska-Masłoń, J.; Guglas, K.; Florczak, A.; Kazimierczak, U.; et al. SRY-Related Transcription Factors in Head and Neck Squamous Cell Carcinomas: In Silico Based Analysis. Curr. Issues Mol. Biol. 2023, 45, 9431-9449. https://doi.org/10.3390/cimb45120592
Kolenda T, Graczyk Z, Żarska B, Łosiewski W, Smolibowski M, Wartecki A, Kozłowska-Masłoń J, Guglas K, Florczak A, Kazimierczak U, et al. SRY-Related Transcription Factors in Head and Neck Squamous Cell Carcinomas: In Silico Based Analysis. Current Issues in Molecular Biology. 2023; 45(12):9431-9449. https://doi.org/10.3390/cimb45120592
Chicago/Turabian StyleKolenda, Tomasz, Zuzanna Graczyk, Barbara Żarska, Wojciech Łosiewski, Mikołaj Smolibowski, Adrian Wartecki, Joanna Kozłowska-Masłoń, Kacper Guglas, Anna Florczak, Urszula Kazimierczak, and et al. 2023. "SRY-Related Transcription Factors in Head and Neck Squamous Cell Carcinomas: In Silico Based Analysis" Current Issues in Molecular Biology 45, no. 12: 9431-9449. https://doi.org/10.3390/cimb45120592