
Changes of Signaling Pathways in Hypothalamic Neurons with Aging


Department Normal Physiology, Yaroslavl State Medical University, ul. Revoliucionnaya 5, 150000 Yaroslavl, Russia
Curr. Issues Mol. Biol. 2023, 45(10), 8289-8308; https://doi.org/10.3390/cimb45100523
Submission received: 28 August 2023
/
Revised: 1 October 2023
/
Accepted: 10 October 2023
/
Published: 12 October 2023
(This article belongs to the Collection Feature Papers in Current Issues in Molecular Biology)
Abstract
:The hypothalamus is an important regulator of autonomic and endocrine functions also involved in aging regulation. The aging process in the hypothalamus is accompanied by disturbed intracellular signaling including insulin/insulin-like growth factor-1 (IGF-1)/growth hormone (GH), phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K)/protein kinase B (AKT)/the mammalian target of rapamycin (mTOR), mitogen activated protein kinase (MAPK), janus kinase (JAK)/signal transducer and activator of transcription (STAT), AMP-activated protein kinase (AMPK), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-ĸB), and nitric oxide (NO). In the current review, I have summarized the current understanding of the changes in the above-mentioned pathways in aging with a focus on hypothalamic alterations.
1. Introduction
The hypothalamus is the regulatory hub controlling homeostasis, reproduction, circadian rhythms, and the endocrine system [1,2]. At the end of the XXth Century, it was proposed that hypothalamus participates in the aging regulation. According to this theory, age-related raising of the hypothalamus threshold for homeostatic signals leads to aging and the appearance of age-related disease in different species [3,4]. The advancement of new research techniques, including cutting-edge approaches in genetics, molecular biology, and neuroscience, allowed the study of the role of the hypothalamus in aging mechanisms in more detail [5,6,7]. In recent years, it has been suggested that the age-related loss of hypothalamic stem cells plays a major role in the development of hypothalamic neuroinflammation and mammalian aging [8,9].
The process of aging is associated with many chronic pathological conditions such as vascular diseases, diabetes mellitus, cancer, and metabolic syndrome. It has been established that aging is accompanied by disturbances of signaling cascades including insulin/insulin-like growth factor-1 (IGF-1)/growth hormone (GH), phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K)/protein kinase B (AKT)/the mammalian target of rapamycin (mTOR), mitogen activated protein kinase (MAPK), janus kinase (JAK)/signal transducer and activator of transcription (STAT), AMP-activated protein kinase (AMPK), and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-ĸB). These pathways regulate energy balance, cellular plasticity, mechanisms supporting homeostasis, growth, reproduction, and inflammation [10,11,12]. However, many studies of intracellular signaling pathways in aging focus on changes in peripheral organs, while little attention is paid to hypothalamic alterations.
2. A Brief Description of the Main Hypothalamic Nuclei with a Focus on Mediobasal Hypothalamus
With rare exceptions (supraoptic (SON) and paraventricular (PVN) nuclei), the majority of hypothalamic nuclei have obscure boundaries. They are no longer thought of as centers with strictly limited functions. PVN and SON nuclei control different physiological functions including stress and the adaptive responses of the organism. The PVN has a non-uniform cellular composition because it contains magnocellular, parvocellular, and neurons with distally projecting axons. The PVN regulates stress reactions, metabolism, growth, reproduction, and activity of the autonomic nervous system. Neurons of the magnocellular SON regulate salt and water homeostasis together with blood pressure [12,13].
The lateral hypothalamic region (LHA) and other hypothalamic nuclei, including the arcuate (ARN), ventromedial (VMN), and dorsomedial (DMN) nuclei, are all involved in regulation of metabolism. In addition, the ARN controls growth and sex gland function together with the median preoptic (MPN) and anteroventral periventricular (AVPV) nuclei [14,15]. The suprachiasmatic nucleus (SCN) is the primary pacemaker of circadian rhythms [16]. The tuberomamillary nucleus is known as the important wake-promoting center and participates in the regulation of temperature, learning, sleep, and behavior [17].
The VMN and ARN are combined into the mediobasal hypothalamus based on their location and functions, such as their role in regulating feeding and sexual behavior [18,19]. From data of D. Cai’s group, this region is extremely important for the control of whole-body aging [5]. In the ARN, proopiomelanocortin (POMC)/cocaine-amphetamine-regulated transcript (CART) neurons and agouti-related peptide (AGRP) neuropeptide Y (NPY) neurons counteractively regulate feeding. POMC and CART have anorexigenic action and increase energy expenditure. NPY and AGRP stimulate feeding behavior (Figure 1). Insulin, together with the leptin, stimulates POMC/CART neurons and inhibits NPY/AGRP ones. Conversely, ghrelin stimulates NPY/AGRP neurons and inhibits POMC/CART cells [15,20]. In contrast to leptin, insulin inhibits the activity of some POMC neurons.
Neurons secreting growth hormone-releasing hormone (GHRH) are found in the ARN [21], and those expressing GH-inhibitory hormone (somatostatin, SS) are located in the anterior periventricular, ARN, VMN, LHA, PVN [22]. The control of reproduction depending on the level of nutrients is mediated by kisspeptin-secreting neurons which regulate gonadotropin releasing hormone (GnRH)-producing neurons (Figure 2). Two populations of kisspeptin synthesizing neurons are located in the hypothalamus, in the ARN, and AVPV nuclei [23]. These ARN neurons, known as KNDy neurons, contain two stimulatory peptides, kisspeptin and neurokinin B (NKB), and an inhibitory peptide, dynorphin [24]. KNDy neurons mediate negative steroid feedback to the GnRH neurons and generate GnRH episodic release, whereas AVPV kisspeptin neurons act as positive feedback mediators and are responsible for GnRH/LH surges [23]. Mammalian GnRH neurons form a dispersed population throughout the rostral forebrain. Most GnRH neurons are observed in the medial septal region, the rostral preoptic area, and the anterior hypothalamic area [25].
In addition to leptin, ghrelin, and insulin, incretin hormones including glucagon-like peptide-1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) also control appetite and body weight (Figure 1 and Figure 2), and act on their cognate receptors GLP-1R and GIPR [26]. GLP-1Rs are expressed throughout the brain, majorly in the solitary tract nucleus, ARN, PVN, and DMN of the hypothalamus, along with central amygdale [27]. Intracerebroventricular injection of GLP-1 inhibits eating and reduces body weight. GLP-1R agonists can inhibit eating through activation of vagal afferent GLP-1Rs and via a direct effect on the brain [28].
3. Insulin/IGF-1/GH Signaling
The insulin-like growth factor (IGF) family includes three known ligands (IGF-1, IGF-2, and insulin). In contrast to invertebrates, mammals, such as mice and humans, have two distinct receptors for insulin and IGF-1, each of which is encoded by a separate gene. While insulin primarily controls the metabolism, IGFs are responsible for long-lasting effects on the regulation of growth, development, and differentiation of cells and tissues [29].
Insulin is synthetized by beta cells in the pancreas and crosses the blood–brain barrier by a saturable mechanism. Insulin in the CNS has opposite effects compared to the periphery, increasing blood glucose levels, decreasing feeding and body weight, and even decreasing the blood levels of insulin [30]. Some recent data indicate that brain insulin downregulates expression of genes involved in glucose metabolism, reduces oxidative stress, regulates the expression of glutamate receptors, upregulates GABA receptors, and suppresses multiple neuropeptides, which contribute to synaptic plasticity and neuronal activity, particularly in the hypothalamus [31].
In vitro biological effects of IGFs are relatively weak and often are observed in the presence of other hormones or growth factors. Possibly, IGFs act as permissive factors to augment the signals of other factors [32].
Insulin can bind to the IGF-1 receptor (IGF-1R) but IGFs bind less well to the insulin receptor (IR) [32]. When IGF-1Rs and IRs are produced in the same cells, some of them form a hybrid receptor [33]. Although structurally, the IGF-1R and the IR are highly homologous, the homology of their C-terminal regions is relatively low [32,33]. IGF-1Rs and IRs are found throughout the CNS including the hypothalamus, and their relative distributions vary, but overlap [34,35]. IRs’ expression is higher than that for IGF-1Rs in the ARN [30]. However, there are no data about changes in the number of IGF-1Rs and IRs in different hypothalamic nuclei with aging.
In contrast to the invertebrate system, mammalian IGF-1 production is regulated by GH secretion from the pituitary. In vivo, many physiological effects of GH are mediated by GH-induced hepatic IGF-1 and local IGF-1 [36]. The secretion of GH is stimulated by GHRH and ghrelin and inhibited by SS. GH exerts its effects by interacting with the GH receptor (GHR), a member of the cytokine receptor superfamily. The GHR is expressed in multiple tissues, including muscle, adipose, heart, kidney, intestine, and bone, with expression being most abundant in the liver [37]. GH secretion is inhibited by IGF-1 in a feedback loop and modulated by other hormones including insulin [38]. Short-term treatment with GH results in rapid insulin-like effects, including the stimulation of lipogenesis and inhibition of lipolysis [39]. However, long-term use of GH counteracts the effects of insulin and induces insulin resistance and diabetes [40,41].
IR signaling is mediated by the action of insulin receptor substrate (IRS) on two canonical pathways, the phosphatidylinositol-4,5-bisphosphate 3-kinase (PI3K)/protein kinase B (AKT)/the mammalian target of rapamycin (mTOR) signaling pathway, and Ras/mitogen activated kinase (MAPK) cascades (Figure 3) [42]. The PI3K/AKT/mTOR signaling pathway is critically important in the regulation of different biological processes such as cell cycle, metabolism, and signal transduction [43,44,45].
GH binding with GHR activates the tyrosine kinase janus kinase 2 (JAK2), which phosphorylates several tyrosine residues on the intracellular domain of the receptor and initiates a multitude of signaling cascades (Figure 3). Activation of the GHR/JAK2 complex leads to activation of signal transducers and activators of transcription (STATs), ERK/MAPK, and PI3K/AKT pathways [37,46].
GH and IGF-1 levels in the blood peak in adolescence and then progressively decline, becoming scarcely detectable in the elderly [47]. The level of GHRH in the median eminence decreases in aged rats, while the SS level in the median eminence, as well as GHRH and SS level in the neuronal somata remains intact [48]. It was proposed that SS-induced inhibition of GH was sensitized in aged rats, indicating a possible cause for the age-related decrease in GH level in plasma [49]. However, the percentage of SS-immunoreactive neurons in the mediobasal hypothalamus, as well as their number, does not change in males and females with aging [22].
In both humans and mice, a substantial increase in GH levels is associated with high risks of diseases and reduced life expectancy [50]. To date, the mechanism of age-related decline in GH and IGF-1 levels remains unclear [51].
Reduced activity of the insulin/IGF1 signaling increases longevity, improves health in aged animals from worms and flies to mice [52,53,54]. However, global reduction in insulin signaling by the deletion of IR, IGF1R, or IGF1 leads to early mortality [55,56]. Total disruption of insulin signaling also has defects such as metabolic syndrome, and reduced body size and fertility [57]. Total deletion of IR receptors in peripheral tissues after adulthood leads to a significant reduction in male life span [58]. On the other hand, insulin in the brain acts as a neuroprotector, lowering damage induced by ischemia, β-amyloid toxicity, oxidative stress, and apoptosis [59]. Brain insulin resistance and low levels of brain insulin often lead to metabolic and cognitive dysfunctions, including obesity, type 2 diabetes, and Alzheimer’s disease [60,61].
Reduced expression of the IGF-1R in the C57BL/6 mice caused only a 5% prolongation of lifespan in females [62]. However, its partial reduction in fat can lead to lifespan extension [63].
Hypothalamic insulin resistance caused by overnutrition occurs more rapidly than in other insulin-sensitive tissues [64]. Nevertheless, selective ablation of IR in the different populations of hypothalamic neurons, including AGRP and POMC cells, has only a moderate effect on energy balance [65,66]. In addition, mice lacking the IR in the MCH neurons of the LHA had a lean phenotype and exhibited improved locomotor activity and insulin sensitivity under a high-fat diet [67].
There is a link between the IGF-1 and GLP-1 signaling pathways. GLP-1, apart from regulating food intake, prevents neuronal death mediated by amyloidogenesis, cerebral glucose deprivation, neuroinflammation, and apoptosis through modulation of PI3/Akt mTOR and MAPK/ERK [68]. GLP-1 downregulation causes oligodendrocyte deterioration, demyelination, glial hyperactivity, immunological dysregulation, and neuroexcitation in the brain [69]. IGF-1 resistance and GLP-1 deficiency impair protective cellular signaling mechanisms, contributing to the progression of neurodegenerative diseases [68,70]. The anti-inflammatory effect of GLP-1 was lost in astrocytes with IGF1-R knockout [71].
However, there are only few data about changes in GLP-1 signaling in aging. The anorexic effect of GLP-1 was attenuated in aged mice [72], thus highlighting the alterations of hypothalamic sensitivity to hormones and environmental factors with aging.
4. PI3K/mTOR/AKT Pathway
The binding of IGF-I or insulin to the α subunit of its specific receptor on a target cell membrane leads to the conformational change in the β subunit, resulting in the activation of receptor tyrosine kinase activity [31,73]. The activated receptor in turn phosphorylates specific substrates, in particular insulin receptor substrate (IRS) and Src homology collagen (Shc) [32]. It is known that mice have four IRS proteins (IRS1-IRS4) but IRS3 is absent in humans [74]. Tyrosine phosphorylated substrates (pIRS-1, pIRS-2, pShc, pGab1) are recognized by some SH-2 domain containing molecules, including growth factor receptor-bound 2 (Grb2), a p85 regulatory subunit of PI3-kinase, and SHP2/Syp. Grb2 is required for IGF-I-induced Ras-MAPK pathway activation (Figure 3) [32]. The interaction between IRS and SHP-2 is important for the crosstalk between IGF-I and the integrin pathway [75].
The PI3K family includes three classes, from I to III [76]. PI3K itself is a heterodimer consisting of regulatory p85 and catalytic p110 subunits. At rest, it is in an inactive state in the cytoplasm of the cell. The p85 regulatory domain of PI3K (p85-PI3K) binds the phosphorylated domains of IRS-1, activating PI3K [77]. The activated enzyme catalyzes the synthesis of phosphatidylinositol-3,4,5-triphosphate (PIP3) from the membrane phosphatidylinositol 4,5-bisphosphate (PIP2). PIP3 provides anchoring of phosphoinositol-dependent protein kinase (PDK-1/2) in the membrane. Membrane-bound PDK-1/2 is activated and, in turn, catalyzes the phosphorylation of inactive AKT. The activation of PI3K induces AKT phosphorylation at two residues: Thr 308 in the kinase domain and Ser473 in the hydrophobic motif. Phosphorylated AKT (pAKT) acquires catalytic properties, dissociates from the membrane, and provides phosphorylation of various intracellular proteins in the cytoplasm and nucleus [78,79]. AKT regulates the activity of some longevity genes, such as mTOR, NF-κB and forkhead box O (FOXO). AKT stimulates mTOR and NF-κB, and inhibits FOXO factors (Figure 3) [80].
An important downstream target of AKT is mTOR. mTOR is a serine/threonine protein kinase that is a part of two different protein complexes: the mTOR complex 1 (mTORC1) and the mTOR complex 2 (mTORC2). mTORC1 is inhibited by rapamycin, promotes protein synthesis and autophagy, and integrates hormonal and environmental signals. mTORC2 is involved in the insulin signaling pathway. In addition to mTOR protein kinase, mTORC1 includes RAPTOR protein and AKT PRAS40 substrate; mTORC2—RICTOR protein and other specific protein subunits such as mSin1 and Protor-1/2. In addition, both mTORC1 and mTORC2 also have the same components: mLST8/GβL as well as the DEPTOR regulatory protein [81].
With aging, the PI3K/AKT/mTOR signaling pathway dysregulates in many tissues including the brain and hypothalamus. There are some reports about changes in separate parts of this system, and selective action may reduce aging manifestations.
IRS-1 and IRS-2 are functionally different. IRS-1 knockdown or overexpression did not affect the IGF signals but IRS-2 knockdown impaired the IGF-1 signals and IGF bioactivities [82,83]. Age-related diseases such as obesity, skin pathology, osteoporosis, sarcopenia, and glucose intolerance were less common in IRS-1 complete knockout mice [84]. Nevertheless, the lifespan of mice was not increased by selective IRS-1 deletion in the liver, muscles, fat, or neurons. Compared to IRS1 deletion in the muscle, liver, and fat, neuron-selective IRS1 knockout increased energy consumption, movement activity, and insulin responsiveness, particularly in aged male mice [85].
There are contradictory data on the role of IRS-2 in aging regulation. Taguchi et al. (2007) reported that mice heterozygous for a null mutation in the IRS-2 in the whole body or separately in the brain had an increased life span [86]. However, using the same mouse model, Selman et al. (2008) observed no evidence for life-span extension and even shortened survival time [87]. It is suggested that IRS-1 is more involved in mitogenic signaling whereas IRS-2 is more important in the regulation of metabolism [88]. In this case, a more severe lifespan-prolonging effect in IRS-1 mutants could be due to lowered cell division while IRS-2 mutants may not demonstrate lifespan extension due to metabolic disturbances [58].
Leptin and insulin action can differently modulate PI3K activity in hypothalamic neurons. It is important for metabolic syndrome, which is a common manifestation of aging [89,90,91]. In the ARN, leptin directly activates PI3K in POMC neurons, but indirectly inhibits it in AGRP neurons [92]. Insulin, vice versa, stimulates PI3K signaling in both POMC and AGRP neurons [93]. Selective inactivation of PI3K reduced leptin-stimulated excitation of hypothalamic POMC neurons in the ARN and caused suppression of food intake [94]. Leptin and insulin influence to decrease food intake was interrupted by PI3K inhibitors [95]. In turn, pharmacological inhibition of PI3K eliminated insulin-induced activation of mTOR [96]. However, the exact role of PI3K in hypothalamic neurons in the regulation of metabolism during aging has yet not been fully established. In C. elegans, mutants lacking any active PI3K are extremely long lived and stress resistant [97].
Despite many studies having demonstrated that aging is closely related to the PI3K/AKT signaling pathway, the detailed mechanism underlying it is still not completely understood. Often constitutive activation of AKT signaling leads to tissue overgrowth and is frequently observed in cancer cells, whereas reduced AKT activity is associated with diabetes and growth defects [98,99]. In aging, the occurrence of diabetes and obesity is associated with insulin resistance [100], which leads to the downregulation of AKT and upregulation of FOXOs. In addition, AKT demonstrates differential actions in separate organs [101,102].
There are different results concerning AKT modulation and aging in the brain. Some studies showed a reduction in AKT activity, others found an increase in its phosphorylation status with the aging process [103,104]. In addition, there are contradictory data on AKT expression in Alzheimer’s disease with evidence of increased and decreased AKT activity [105,106]. In the hypothalamus of rodents, the expression of AKT does not change or there are small disturbances with aging [107,108]. On the other hand, activation of the PI3K/AKT/mTOR pathway in the hypothalamus leads to a comparable weight increase, and obesity is often observed with aging [92].
AKT phosphorylates FOXO, induces its nuclear translocation, and inhibits its action in vitro and in vivo. FOXO performs a considerable role in aging and age-related diseases including neurodegenerative and oncological pathology [109]. FOXO overexpression promotes longevity in many species including mammals by combined autocrine and paracrine effects [108]. However, there are contradictory facts that FOXO activity elevates with aging. Tumor necrosis factor (TNF)-α raises the FOXO1 activity by preventing its phosphorylation. Enlarged FOXO1 activity suppresses the gnrh1 gene and activates the NF-κB inflammatory signaling, preventing GnRH secretion [110].
Hypothalamic FOXO1 increases food intake and body weight by stimulating the transcription of orexigenic NPY/AGRP and suppressing POMC neurons in the ARN [45,111,112]. Nevertheless, dietary disturbances are not necessarily associated with the changes in PI3K/AKT/mTOR and FOXO signaling [112].
Different studies have shown that inactivation of mTOR signaling can prolong the duration of life, which might explain the lifespan extension of mice with rapamycin therapy [113,114]. Diet-induced hyperactivation of mTORC1 induces metabolic disturbances, including obesity and type 2 diabetes, as well as cancer and neurodegenerative diseases [115,116,117]. Upregulation of mTORC1 and mTORC2 inhibitory protein DEPTOR in hypothalamic neurons reduced obesity and improves glucose metabolism [118]. Nevertheless, the expression of mTOR in separate hypothalamic nuclei is different through aging. The proportion of mTOR-IR decreased in DMN and VMN neurons and increased in the ARN in 12-month-old and 24-month-old rats [119].
Loss of RICTOR, a part of the mTORC2 complex, results in a sex-specific reduction in male life expectancy even when the intervention is initiated at maturity, in contrast to deletion of IRS1 [120]. Moreover, both male and female mice exhibited a decreased lifespan, activity, glucose tolerance, and insulin responsiveness after knockout of RICTOR in the hypothalamus [121].
In addition to mTOR, AKT through different pathways could trigger the activation of the NF-κB system, which inhibits apoptosis, autophagy, and stimulates inflammatory responses, which is very important for aging [122].
5. Ras/Raf/MEK/ERK Pathway
The mitogen-activated protein kinases/extracellular signal-regulated kinase (MAPK/ERK) pathway is reported to be associated with cell proliferation, differentiation, migration, senescence, and apoptosis. Insulin is also able to modulate cell growth and proliferation through induction of Shc and its downstream targets Ras, ERK, and mitogen-activated protein kinase (MAPK). In the Ras/MAPK signaling pathway, activated IRSs interact with growth factor receptor-bound protein 2 (Grb2). The tyrosine 891 in IRS-1 is known to be a Grb2 binding site phosphorylated by IGF/insulin stimuli, which is important for the activation of the Ras-MAPK cascade [123]. Grb2, in turn, can activate MAPK cascades via an interaction with the Ras guanine nucleotide exchange factor SOS. Subsequently, SOS activates the small G protein Ras, which in turn recruits MAPK3, also known as Raf kinase. The Raf kinase phosphorylates and activates a MAPK/ERK kinase (MEK1 or MEK2). The MEK phosphorylates and activates a mitogen-activated protein kinase (MAPK). MAPK is referred to as extracellular signal-regulated kinase (ERK). ERK (MAPK) further phosphorylates different proteins in the cytoplasm or transcription factors in the nucleus (Figure 3) [124].
This pathway is more often activated by IGF-1R compared to IR [125]. Moreover, different IRα isoforms stimulate specific downstream pathways: the IR-A isoform better activates the Shc/Ras/ERK/MAPK signaling, while the IR-B stimulates PI3K/Akt/mTOR [126,127].
There is evidence of ERK/MAPK impairment in the brain pathology. ERK/MAPK stimulation decreases neuronal injury following ischemia-hypoxia [128]. Treatment of brain-derived neurotrophic factor (BDNF) leads to elevation of ERK/MAPK activity during 12 h after ischemic-hypoxic injury. Resveratrol and fisetin also activate ERK/MAPK and inhibit cell death in a model of Huntington’s disease [129].
During aging, the expression of phosphorylated ERK1/2 and MAPK was reduced in the rat cerebral cortex, striatum, and hippocampus [130]. However, ERK1/2 phosphorylation was increased in the hypothalamus of aged rats [131].
In the hypothalamus, insulin decreases mRNA expression of orexigenic neuropeptides NPY and AgRP through an ERK/MAPK dependent mechanism, [132]. Obese mice also exhibit an increased hypothalamic expression of the MAPK phosphatase 3 associated with a reduced phosphorylation of ERK [133,134].
In the SCN of the hypothalamus, nocturnal light activates ERK/MAPK [135], and inhibition of ERK/MAPK prevents the light response [136]. Thus, ERK/MAPK is an important signal pathway involved in clock function. Rhythmicity in the SCN declines with aging, and impairment in the ERK/MAPK pathway can play an important role in this process. However, there are no direct data about changes in the ERK/MAPK expression in the SCN with aging.
6. AMPK Signaling
AMPK has an important role in energy metabolism and is activated in conditions of low energy [137,138]. It is a serine/threonine protein kinase formed by a catalytic α subunit and regulatory β and γ subunits. Each subunit has several isoforms, which are differently expressed in separate tissues and have diverse cellular localization [139]. Increased AMP/ADP ratio stimulates AMPK by allosteric regulation. AMPK activation is dependent on the energy status and the action of upstream stimulatory and inhibitory signaling pathways [140]. AMPK is activated by phosphorylation of upstream kinases including the tumor suppressor liver kinase B1 (LKB1), Ca2+/calmodulin-dependent protein kinase α, and β (CaMKKα and CaMKKβ), protein phosphatase-2Cα (PP2Cα) [139,140].
AMPK facilitates energy manufacture from glucose and fatty acids in stress conditions and suppresses synthesis of proteins, cholesterol, and glycogen [137,138,139,140]. Caloric restriction can elevate AMPK expression whereas food excess decreases AMPK activity and simultaneously induces insulin resistance and the emergence of metabolic syndrome, including such symptoms as obesity, diabetes, and cardiovascular diseases. AMPK activity decreases with aging, elevating the aging process [141,142].
AMPK indirectly inhibits the NF-κB pathway, activates FoxO signaling, and autophagy. AMPK can suppress the action of mTOR complex 1 (mTORC1) either by direct phosphorylating the Raptor, a regulatory component of mTORC1, or by the phosphorylation of tuberous sclerosis protein 2 (TSC2), which consequently inhibits the mTOR activity [141,142]. In turn, S6K1, a downstream target of the insulin/IGF-mTOR pathway, inhibits AMPK activity. S6K1 knockout activates AMPK signaling and increases the lifespan. AKT also inhibits AMPK signaling (Figure 3) [143,144].
In the hypothalamus, AMPK activation drives feeding by modulating mitochondrial fatty acid oxidation and intracellular levels of reactive oxygen species [145,146]. AMPK activity is inhibited in the ARN and PVN by leptin, and in multiple hypothalamic regions by insulin, high glucose, and refeeding [147,148]. Ghrelin administration induced a time-dependent AMPK increase in VMN [149]. There is an increase in hypothalamic AMPK activity in rats with diet-induced obesity, and the increased AMPK activity is associated with oxidative stress and inflammation in the hypothalamus [150]. The baseline pAMPK level was increased with age in the rat hypothalamus, whereas the total AMPK level remained unchanged. The pAMPK/AMPK ratio was also significantly higher in the aged rats [151].
Metformin, a well-known anti-aging drug, is also an activator of AMPK at the periphery, reducing glucose levels in type 2 diabetes. Other phytochemical geroprotectors such as berberine, quercetin, and resveratrol, have also been reported to activate AMPK signaling [141,142,152]. Recently, strong evidence was obtained that the important target for metformin is the hypothalamus, and its effects on the hypothalamic signaling plays a significant role to metformin-induced improvement of metabolic disturbances and increase in lifespan [153].
Metformin does not stimulate and even reduces AMPK activity in the hypothalamus, in contrast to peripheral tissues, which results in the activation of thermogenesis and a reduction in body weight and adiposity [154,155]. In the hypothalamus of obese agouti-mice, AMPK and STAT3 activities were increased, and metformin treatment stabilized the AMPK and STAT3 levels, augmented the activity of AKT, and increased POMC expression in the hypothalamus [61,155].
7. JAK/STAT Signaling
The JAK/STAT pathway mediates signals from many hormones and cytokines including GH, leptin, erythropoietin, and cytokines. JAK/STAT proteins are involved in the control of a variety of biological processes including proliferation, apoptosis, the immune response, inflammation, food intake, and energy homeostasis [156,157].
Upon the binding of a ligand to its cognate receptor, receptor-associated JAKs are dimerized and activated through autophosphorylation. Inactive cytosolic STAT monomers are transformed into active homodimers, heterodimers, or tetramers after being phosphorylated by JAKs. The active STATs can then translocate into the nucleus where they act as transcription factors to regulate gene expression. [156,157].
In mammals, there are four members of the JAK family: JAK1, JAK2, JAK3, and TYK2, and seven members of the STAT family: STAT1–4, STAT5A/B, and STAT6. Different combinations of JAKs and STATs are stimulated with a high specificity by various cytokines or growth factors. Other kinases, including Flt3R and pyruvate kinase, can phosphorylate STATs in addition to JAKs (Figure 3) [93,156,157].
JAK2, STAT3, and STAT5 are necessary for signaling through both the growth hormone (GH) and leptin receptors. STAT5 is the predominant STAT protein activated by GH [31,158], STAT3 and STAT5 mediate leptin-induced anorexic effects [158,159]. In addition to JAK2, activation of leptin receptors directly or indirectly triggers multiple signaling pathways including MAPK, FOXO1, AMPK, and others [159]. GH except JAK2 can activate Lyn (SRC family kinase member). Lyn stimulation by GH was shown to activate MAPKs. GH has also been shown to stimulate the PI3K pathway, probably through phosphorylation of the IRS [160]. Additionally, many impacts of the AKT/mTOR pathway on the immune system and aging are performed via STAT3 signaling [122].
In the hypothalamus, leptin elevates the transcription of suppressor of cytokine signaling 3 (SOCS3). SOCS3 suppresses leptin actions by a negative feedback loop [161]. In the ARN, STAT3 phosphorylation facilitates the leptin-mediated transcriptional regulation of POMC, AgRP, and NPY by binding to the promoters, which activates POMC and inhibits AGRP expression, thereby reducing food intake and elevating energy consumption [159]. In mice, neural-specific STAT3 knockout and mutation of leptin receptors that do not bind STAT3 leads to hyperphagia, obesity, reduced energy consumption, and augmented mRNA level of AGRP [155,160].
Exogenous leptin increases the satiating effect of GLP-1. The leptin receptors deficiency in neuronal cells expressing GLP-1 located in the solitary tract nucleus resulted in hyperphagia and obesity [26].
Basal levels of phosphorylated STAT and total STAT protein were not significantly different among young and aged rats [152,161]. However, leptin receptor expression was higher in the young compared with aged rats. These data indicate that aged rats demonstrated reduced signal transduction in response to centrally administered leptin [162].
8. Sirtuins
Sirtuins (SIRTs) are an evolutionarily conserved family of NAD+-dependent deacylases that play a key role in longevity in various organisms, including invertebrates and vertebrates [163]. The mammalian sirtuin family consists of seven members, SIRT1–7. SIRT2 is observed in the cytoplasm, while SIRT1, SIRT6, and SIRT7 are located in the nucleus [163,164].
SIRT isotypes vary in their functional activity: SIRT1, SIRT2, SIRT3, and SIRT-7 mainly display properties of (NAD+)-dependent deacetylase; SIRT4 and SIRT6 possess activities of deacetylases and ADP-ribosyltransferases. SIRT5, in addition to deacetylase activity, has functions of demalonylase and desuccincylase. SIRTs possess histone deacetylase activity, which is (NAD+)-dependent. Since NAD+ is a significant redox signaling molecule, SIRTs control antioxidant and redox signaling cellular pathways [164,165]. Due primarily to their impact on the p53 and FOXO activities, SIRTs play a significant role in cellular homeostasis and apoptosis [164,165]. SIRT1 suppresses the NF-ĸB signaling (Figure 3), increases the activity of the key antioxidant enzymes, and has a variety of other effects on glucose and lipid metabolism [163,164,166]. The interaction between neuroinflammation, neurodegeneration, and metabolic changes has been shown to be heavily dependent on SIRTs [167].
SIRT1 expression decreases with age in the human liver, heart, kidneys, brain, and lungs. Overexpression of Sir2 in yeasts, or its homolog SIRT1 in animals, can increase lifespan [168,169]. The geroprotective properties of SIRT1, 2, 3, 6, and 7 are mediated by a variety of cellular signal cascades. The geroprotective effect of SIRT1 is regulated via p53, NF-κB, mTOR, PGC1α, and FOXO signal pathways [169,170,171]. Sirtuins are also required for caloric restriction-induced life span extension [171,172].
Resveratrol, a natural polyphenolic compound and potent geroprotector, allosterically modifies the activity of SIRT1. On the other hand, resveratrol also directly targets proteins other than SIRT1, such as AMPK and cAMP phosphodiesterases [169,171,173].
The level of SIRT1 decreases in the whole hypothalamus and separately in the ARN, VMN, DMN, and SCN with advanced age [171,174]. Supporting SIRT1 activity in the SCN could delay aging and extend lifespan [171,175]. In male and female transgenic mice overexpressing SIRT1 (BRASTO mice) in the DMN and LHA, the lifespan was increased [171,176]. Hypothalamic SIRT1 affects insulin sensitivity, systemic glucose, and lipid metabolism in the peripheral organs. SIRT1 in POMC neurons is required for normal adaptations against diet-induced obesity [177].
9. Nitric Oxide
Nitric oxide (NO) is a gaseous signal molecule participating in numerous brain functions including behavior, learning, memory, sleep, feeding, pain, and sexual activity [178,179]. NO is produced by NO synthase (NOS). There are three NOS isoforms: neuronal NOS (nNOS or NOS1), inducible NOS (iNOS or NOS2), and endothelial NOS (eNOS or NOS3). Ca2+ influx through NMDA receptors is necessary for nNOS activation and the subsequent generation of NO. Once NO is produced, it can easily diffuse through membranes and bind to the NO receptor, soluble guanylate cyclase, which synthetizes the second messenger cGMP [178,179,180].
In the hypothalamus, nNOS expression is largely restricted to areas involved in the control of bodily functions, including metabolism and reproduction. nNOS-expressing neurons are located in the preoptic region, supraoptic, ventral premamillary nucleus, DMN, VMN, PVN, and LHA [181,182,183].
Aging is associated with high levels of reactive oxygen species and the NO metabolite peroxynitrite in the brain [184,185]. Redundant NO production causes cell death, but augmented NO release can prevent apoptosis [186]. nNOS expression and activity increases in neurodegenerative diseases, epilepsy, inflammation, and ischemia [187,188]. There is some evidence that increased NO production in the hippocampus reduces hippocampal neurogenesis [186]. While nNOS-expressing neurons are thought to be more resistant to neurotoxic effects, NO can serve a protective role throughout aging [184].
In young rats, weakly nNOS-immunopositive neurons were found in the DMN and VMN, but were missing in the ARN. In aged rats, the number of neurons containing nNOS significantly increased in the DMN and VMN, and a large number of nNOS-immunoreactive neurons was found in the ARN [183].
10. Hypothalamic Inflammation in Aging
In the aged brain, augmented levels of pro-inflammatory cytokines including TNF, interleukin (IL) 1β, IL 6, and reduced levels of anti-inflammatory cytokines, such as IL 2 and IL 10 are observed. Activation of the signaling molecules such as Toll-like receptor 4 (TLR4), myeloid differentiation primary response gene 88 (MyD88), nuclear factor kappa B (NF-κB), inhibitor of nuclear factor kappa B kinase subunit beta (IKK), and c-Jun N-terminal kinase (JNK) is associated with the development of the obesity and the metabolic syndrome in the mediobasal hypothalamus [5,8,189]. The NF-κB signaling acts in close co-operation with a number of pathways, including JAK-STAT3 and insulin/IGF-1 [125]. The NF-κB system is stimulated by the PI3K/AKT/IKKα/β pathway. Nevertheless, it is known that InsR/IGF-1R can also affect the NF-κB complex through the RAF-1 and MAPK pathways [189].
Latest studies have also revealed a crucial role of this hypothalamic microinflammation in the aging processes. In the mice mediobasal hypothalamus, suppression of the NF-κB signaling extended the lifespan, whereas stimulation of the NF-κB pathway reduced the duration of life [5,190]. GABA-ergic and BDNF-expressing neurons of the mediobasal hypothalamus are vulnerable to hypothalamic inflammation induced by a high-fat diet. In this case, hypothalamic astrocytes miss their terminal processes, and reduce GABA uptake. Elevated GABA concentration in the intercellular space suppressed BDNF-expressing neurons [191,192]. Vice versa, the injection of BDNF viral vector into the mediobasal hypothalamus of middle-aged mice reduced the expression of inflammatory genes in the hypothalamus and prevented age-associated metabolic disturbances [193]. In the ARN, stimulation of GABAB receptor in POMC neurons prevented hypothalamic inflammation, obesity, and insulin resistance in mice [194]. Continued hypothalamic inflammation activates glutamatergic transmission, which can lead to hypertension as a part of metabolic syndrome [195].
GLP-1 also exerts its anti-inflammatory response by expressing GLP-1R in astrocytes and glial cells during neuroinflammation. This anti-inflammation effect of GLP-1 is realized via IGF-1 receptor and NF-κB signaling [71].
In rodents, NF-κB signaling suppresses GnRH gene transcription that can induce the age-associated GH decrease. However, intracerebroventricular or subcutaneous GnRH treatment prevented age-related decrease in hypothalamic hippocampal neurogenesis, restored muscular strength, skin thickness, bone mass, and collagen integrity in the caudal tendon in mice [8].
11. Conclusions Remarks
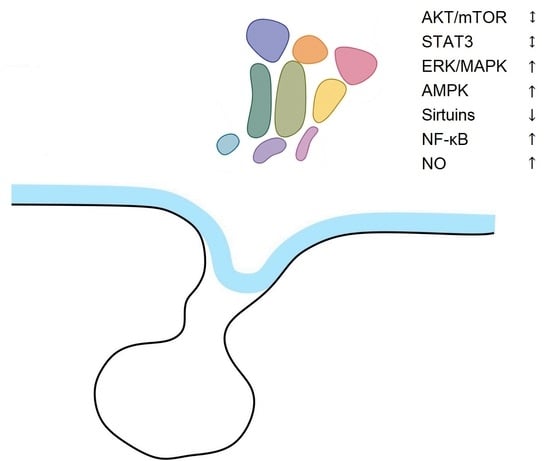
Based on Dillman’s elevation theory of aging, senescence is associated with an increase in the threshold of hypothalamic responsiveness to homeostatic signals. The aging process in the hypothalamus is accompanied by impaired intracellular signaling including PI3K/AKT/mTOR, ERK/MAPK, AMPK, Sirtuins, NF-κB, NO, and disturbances in the release of neurotransmitters (e.g., GnRH, NO, NPY, α-MSH). There are some differences between disturbances of the signaling pathways in the hypothalamus and periphery. The activity of ERK/MAPK, NF-κB, and nNOS increases, SIRT1 decreases, and the expression of AKT and STAT3 does not change in the aging hypothalamus in rodents (for a short summary, see Table 1). In addition, there are some differences in signaling in functionally antagonistic groups of hypothalamic neurons, for example POMC and AGRP neurons. The mechanisms of hypothalamic aging are still not fully understood, despite recent advances in our understanding of the organization of hypothalamic networks. Utilizing a variety of research techniques, such as molecular biology and genetics, will aid in the explanation of this issue.
Funding
This work was supported by Russian Science Foundation (Project No. 19-15-00039).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Yang, F.; Zhao, S.; Wang, P.; Xiang, W. Hypothalamic neuroendocrine integration of reproduction and metabolism in mammals. J. Endocrinol. 2023, 258, e230079. [Google Scholar] [CrossRef]
- Haspula, D.; Cui, Z. Neurochemical Basis of Inter-Organ Crosstalk in Health and Obesity: Focus on the Hypothalamus and the Brainstem. Cells 2023, 12, 1801. [Google Scholar] [CrossRef]
- Dilman, V.M. Age-associated elevation of hypothalamic, threshold to feedback control, and its role in development, ageine, and disease. Lancet 1971, 1, 1211–1219. [Google Scholar] [CrossRef]
- Dilman, V.M.; Anisimov, V.N. Hypothalmic mechanisms of ageing and of specific age pathology--I. Sensitivity threshold of hypothalamo-pituitary complex to homeostatic stimuli in the reproductive system. Exp. Gerontol. 1979, 14, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Khor, S. Hypothalamic microinflammation. Handb. Clin. Neurol. 2021, 181, 311–322. [Google Scholar] [CrossRef]
- Kim, K.; Choe, H.K. Role of hypothalamus in aging and its underlying cellular mechanisms. Mech. Ageing Dev. 2019, 177, 74–79. [Google Scholar] [CrossRef]
- Masliukov, P.M.; Nozdrachev, A.D. Hypothalamic Regulatory Mechanisms of Aging. J. Evol. Biochem. Phys. 2021, 57, 473–491. [Google Scholar] [CrossRef]
- Zhang, G.; Li, J.; Purkayastha, S.; Tang, Y.; Zhang, H.; Yin, Y.; Li, B.; Liu, G.; Cai, D. Hypothalamic programming of systemic ageing involving IKK-β, NF-κB and GnRH. Nature 2013, 497, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kim, M.S.; Jia, B.; Yan, J.; Zuniga-Hertz, J.P.; Han, C.; Cai, D. Hypothalamic stem cells control ageing speed partly through exosomal miRNAs. Nature 2017, 548, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Moskalev, A.A.; Aliper, A.M.; Smit-McBride, Z.; Buzdin, A.; Zhavoronkov, A. Genetics and epigenetics of aging and longevity. Cell Cycle 2014, 13, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Tabibzadeh, S. Signaling pathways and effectors of aging. Front. Biosci. (Landmark Ed.) 2021, 26, 50–96. [Google Scholar] [CrossRef]
- Qin, C.; Li, J.; Tang, K. The Paraventricular Nucleus of the Hypothalamus: Development, Function, and Human Diseases. Endocrinology 2018, 159, 3458–3472. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.H.; Ludwig, M.; Tasker, J.G.; Stern, J.E. Somato-dendritic vasopressin and oxytocin secretion in endocrine and autonomic regulation. J. Neuroendocrinol. 2020, 32, e12856. [Google Scholar] [CrossRef]
- Wang, L.; Moenter, S.M. Differential Roles of Hypothalamic AVPV and Arcuate Kisspeptin Neurons in Estradiol Feedback Regulation of Female Reproduction. Neuroendocrinology. 2020, 110, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.G.; Kanoski, S.E.; Sanchez-Watts, G.; Langhans, W. The physiological control of eating: Signals, neurons, and networks. Physiol. Rev. 2022, 102, 689–813. [Google Scholar] [CrossRef]
- Patton, A.P.; Hastings, M.H. The suprachiasmatic nucleus. Curr. Biol. 2018, 28, R816–R822. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Nakamura, T.; Yanai, K. Histaminergic neurons in the tuberomammillary nucleus as a control centre for wakefulness. Br. J. Pharmacol. 2021, 178, 750–769. [Google Scholar] [CrossRef]
- Mota, C.M.D.; Madden, C.J. Mediobasal hypothalamic neurons contribute to the control of brown adipose tissue sympathetic nerve activity and cutaneous vasoconstriction. J. Therm. Biol. 2023, 114, 103551. [Google Scholar] [CrossRef]
- Kannangara, H.; Cullen, L.; Miyashita, S.; Korkmaz, F.; Macdonald, A.; Gumerova, A.; Witztum, R.; Moldavski, O.; Sims, S.; Burgess, J.; et al. Emerging roles of brain tanycytes in regulating blood-hypothalamus barrier plasticity and energy homeostasis. Ann. N. Y. Acad. Sci. 2023, 1525, 61–69. [Google Scholar] [CrossRef]
- Vohra, M.S.; Benchoula, K.; Serpell, C.J.; Hwa, W.E. AgRP/NPY and POMC neurons in the arcuate nucleus and their potential role in treatment of obesity. Eur. J. Pharmacol. 2022, 915, 174611. [Google Scholar] [CrossRef]
- Decourtye-Espiard, L.; Clemessy, M.; Leneuve, P.; Mire, E.; Ledent, T.; Le Bouc, Y.; Kappeler, L. Stimulation of GHRH Neuron Axon Growth by Leptin and Impact of Nutrition during Suckling in Mice. Nutrients 2023, 15, 1077. [Google Scholar] [CrossRef]
- Vishnyakova, P.A.; Moiseev, K.Y.; Porseval, V.V.; Pankrasheval, L.G.; Budnikl, A.F.; Nozdrachev, A.D.; Masliukov, P.M. Somatostatin-Expressing Neurons in the Tuberal Region of Rat Hypothalamus during Aging. J. Evol. Biochem. Phys. 2021, 57, 1480–1489. [Google Scholar] [CrossRef]
- Prashar, V.; Arora, T.; Singh, R.; Sharma, A.; Parkash, J. Hypothalamic Kisspeptin Neurons: Integral Elements of the GnRH System. Reprod. Sci. 2023, 30, 802–822. [Google Scholar] [CrossRef] [PubMed]
- Velasco, I.; Franssen, D.; Daza-Dueñas, S.; Skrapits, K.; Takács, S.; Torres, E.; Rodríguez-Vazquez, E.; Ruiz-Cruz, M.; León, S.; Kukoricza, K.; et al. Dissecting the KNDy hypothesis: KNDy neuron-derived kisspeptins are dispensable for puberty but essential for preserved female fertility and gonadotropin pulsatility. Metabolism 2023, 144, 155556. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.E.; Coolen, L.M.; Hoffman, G.E.; Hrabovszky, E. Highlights of neuroanatomical discoveries of the mammalian gonadotropin-releasing hormone system. J. Neuroendocrinol. 2022, 34, e13115. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Meier, J.J. Incretin hormones: Their role in health and disease. Diabetes Obes. Metab. 2018, 20 (Suppl. S1), 5–21. [Google Scholar] [CrossRef]
- Biglari, N.; Gaziano, I.; Schumacher, J.; Radermacher, J.; Paeger, L.; Klemm, P.; Chen, W.; Corneliussen, S.; Wunderlich, C.M.; Sue, M.; et al. Functionally distinct POMC-expressing neuron subpopulations in hypothalamus revealed by intersectional targeting. Nat. Neurosci. 2021, 24, 913–929. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, L.; Zhang, J.; Conde, K.; Phansalkar, J.; Li, Z.; Yao, L.; Xu, Z.; Wang, W.; Zhou, J.; et al. Glucose-sensing glucagon-like peptide-1 receptor neurons in the dorsomedial hypothalamus regulate glucose metabolism. Sci. Adv. 2022, 8, eabn5345. [Google Scholar] [CrossRef]
- Baxter, R.C. Signaling Pathways of the Insulin-like Growth Factor Binding Proteins. Endocr. Rev. 2023, 44, 753–778. [Google Scholar] [CrossRef]
- Chen, W.; Cai, W.; Hoover, B.; Kahn, C.R. Insulin action in the brain: Cell types, circuits, and diseases. Trends Neurosci. 2022, 45, 384–400. [Google Scholar] [CrossRef]
- Cai, W.; Zhang, X.; Batista, T.M.; García-Martín, R.; Softic, S.; Wang, G.; Ramirez, A.K.; Konishi, M.; O’Neill, B.T.; Kim, J.H.; et al. Peripheral Insulin Regulates a Broad Network of Gene Expression in Hypothalamus, Hippocampus, and Nucleus Accumbens. Diabetes 2021, 70, 1857–1873. [Google Scholar] [CrossRef]
- Hakuno, F.; Takahashi, S.I. IGF1 receptor signaling pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef]
- Jiráček, J.; Selicharová, I.; Žáková, L. Mutations at hypothetical binding site 2 in insulin and insulin-like growth factors 1 and 2. Vitam. Horm. 2023, 123, 187–230. [Google Scholar] [CrossRef]
- Pomytkin, I.; Costa-Nunes, J.P.; Kasatkin, V.; Veniaminova, E.; Demchenko, A.; Lyundup, A.; Lesch, K.P.; Ponomarev, E.D.; Strekalova, T. Insulin receptor in the brain: Mechanisms of activation and the role in the CNS pathology and treatment. CNS Neurosci. Ther. 2018, 24, 763–774. [Google Scholar] [CrossRef]
- Li, M.; Quan, C.; Toth, R.; Campbell, D.G.; MacKintosh, C.; Wang, H.Y.; Chen, S. Fasting and Systemic Insulin Signaling Regulate Phosphorylation of Brain Proteins That Modulate Cell Morphology and Link to Neurological Disorders. J. Biol. Chem. 2015, 290, 30030–30041. [Google Scholar] [CrossRef]
- Rotwein, P. Regulation of gene expression by growth hormone. Mol. Cell Endocrinol. 2020, 507, 110788. [Google Scholar] [CrossRef] [PubMed]
- Al-Samerria, S.; Radovick, S. Exploring the Therapeutic Potential of Targeting GH and IGF-1 in the Management of Obesity: Insights from the Interplay between These Hormones and Metabolism. Int. J. Mol. Sci. 2023, 24, 9556. [Google Scholar] [CrossRef]
- Al-Samerria, S.; Radovick, S. The Role of Insulin-like Growth Factor-1 (IGF-1) in the Control of Neuroendocrine Regulation of Growth. Cells 2021, 10, 2664. [Google Scholar] [CrossRef]
- Likitnukul, S.; Thammacharoen, S.; Sriwatananukulkit, O.; Duangtha, C.; Hemstapat, R.; Sunrat, C.; Mangmool, S.; Pinthong, D. Short-Term Growth Hormone Administration Mediates Hepatic Fatty Acid Uptake and De Novo Lipogenesis Gene Expression in Obese Rats. Biomedicines 2023, 11, 1050. [Google Scholar] [CrossRef] [PubMed]
- Høgild, M.L.; Hjelholt, A.J.; Hansen, J.; Pedersen, S.B.; Møller, N.; Wojtaszewski, J.F.P.; Johannsen, M.; Jessen, N.; Jørgensen, J.O.L. Ketone Body Infusion Abrogates Growth Hormone-Induced Lipolysis and Insulin Resistance. J. Clin. Endocrinol. Metab. 2023, 108, 653–664. [Google Scholar] [CrossRef]
- Sharma, R.; Kopchick, J.J.; Puri, V.; Sharma, V.M. Effect of growth hormone on insulin signaling. Mol. Cell Endocrinol. 2020, 518, 111038. [Google Scholar] [CrossRef]
- Sędzikowska, A.; Szablewski, L. Insulin and Insulin Resistance in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 9987. [Google Scholar] [CrossRef] [PubMed]
- Solinas, G.; Becattini, B. PI3K and AKT at the Interface of Signaling and Metabolism. Curr. Top Microbiol. Immunol. 2022, 436, 311–336. [Google Scholar] [CrossRef]
- Kumar, M.; Bansal, N. Implications of Phosphoinositide 3-Kinase-Akt (PI3K-Akt) Pathway in the Pathogenesis of Alzheimer’s Disease. Mol. Neurobiol. 2022, 59, 354–385. [Google Scholar] [CrossRef]
- Varela, L.; Horvath, T.L. Leptin and insulin pathways in POMC and AgRP neurons that modulate energy balance and glucose homeostasis. EMBO Rep. 2012, 13, 1079–1086. [Google Scholar] [CrossRef]
- Bergan-Roller, H.E.; Sheridan, M.A. The growth hormone signaling system: Insights into coordinating the anabolic and catabolic actions of growth hormone. Gen. Comp. Endocrinol. 2018, 258, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Ashpole, N.M.; Sanders, J.E.; Hodges, E.L.; Yan, H.; Sonntag, W.E. Growth hormone, insulin-like growth factor-1 and the aging brain. Exp. Gerontol. 2015, 68, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Spik, K.; Sonntag, W.E. Increased pituitary response to somatostatin in aging male rats: Relationship to somatostatin receptor number and affinity. Neuroendocrinology 1989, 50, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Biagetti, B.; Puig-Domingo, M. Age-Related Hormones Changes and Its Impact on Health Status and Lifespan. Aging Dis. 2023, 14, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Hage, C.; Salvatori, R. Growth Hormone and Aging. Endocrinol. Metab. Clin N. Am. 2023, 52, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Bartke, A. Growth Hormone and Aging: Updated Review. World J. Mens. Health 2019, 37, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Dravecz, N.; Shaw, T.; Davies, I.; Brown, C.; Ormerod, L.; Vu, G.; Walker, T.; Taank, T.; Shirras, A.D.; Broughton, S.J. Reduced Insulin Signaling Targeted to Serotonergic Neurons but Not Other Neuronal Subtypes Extends Lifespan in Drosophila melanogaster. Front. Aging Neurosci. 2022, 14, 893444. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.V. Recent Progress in Regulation of Aging by Insulin/IGF-1 Signaling in Caenorhabditis elegans. Mol. Cells 2022, 45, 763–770. [Google Scholar] [CrossRef]
- Bartke, A.; Brown-Borg, H. Mutations Affecting Mammalian Aging: GH and GHR vs IGF-1 and Insulin. Front. Genet. 2021, 12, 667355. [Google Scholar] [CrossRef]
- Liu, J.P.; Baker, J.; Perkins, A.S.; Robertson, E.J.; Efstratiadis, A. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (Igf1r). Cell 1993, 75, 59–72. [Google Scholar] [CrossRef]
- Accili, D.; Drago, J.; Lee, E.J.; Johnson, M.D.; Cool, M.H.; Salvatore, P.; Asico, L.D.; José, P.A.; Taylor, S.I.; Westphal, H. Early neonatal death in mice homozygous for a null allele of the insulin receptor gene. Nat. Genet. 1996, 12, 106–109. [Google Scholar] [CrossRef]
- Rincon, M.; Muzumdar, R.; Atzmon, G.; Barzilai, N. The paradox of the insulin/IGF-1 signaling pathway in longevity. Mech. Ageing Dev. 2004, 125, 397–403. [Google Scholar] [CrossRef]
- Bokov, A.F.; Garg, N.; Ikeno, Y.; Thakur, S.; Musi, N.; DeFronzo, R.A.; Zhang, N.; Erickson, R.C.; Gelfond, J.; Hubbard, G.B.; et al. Does reduced IGF-1R signaling in Igf1r+/− mice alter aging? PLoS ONE 2011, 6, e26891. [Google Scholar] [CrossRef]
- Agrawal, R.; Reno, C.M.; Sharma, S.; Christensen, C.; Huang, Y.; Fisher, S.J. Insulin action in the brain regulates both central and peripheral functions. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E156–E163. [Google Scholar] [CrossRef]
- Ezkurdia, A.; Ramírez, M.J.; Solas, M. Metabolic Syndrome as a Risk Factor for Alzheimer’s Disease: A Focus on Insulin Resistance. Int. J. Mol. Sci. 2023, 24, 4354. [Google Scholar] [CrossRef]
- Shpakov, A.O.; Derkach, K.V.; Berstein, L.M. Brain signaling systems in the Type 2 diabetes and metabolic syndrome: Promising target to treat and prevent these diseases. Future Sci. OA 2015, 1, FSO25. [Google Scholar] [CrossRef]
- Merry, T.L.; Kuhlow, D.; Laube, B.; Pöhlmann, D.; Pfeiffer, A.F.H.; Kahn, C.R.; Ristow, M.; Zarse, K. Impairment of insulin signalling in peripheral tissue fails to extend murine lifespan. Aging Cell 2017, 16, 761–772. [Google Scholar] [CrossRef]
- Blüher, M.; Kahn, B.B.; Kahn, C.R. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 2003, 299, 572–574. [Google Scholar] [CrossRef]
- Ono, H. Molecular Mechanisms of Hypothalamic Insulin Resistance. Int. J. Mol. Sci. 2019, 20, 1317. [Google Scholar] [CrossRef]
- Könner, A.C.; Janoschek, R.; Plum, L.; Jordan, S.D.; Rother, E.; Ma, X.; Xu, C.; Enriori, P.; Hampel, B.; Barsh, G.S.; et al. Insulin action in AgRP-expressing neurons is required for suppression of hepatic glucose production. Cell Metab. 2007, 5, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Shin, A.C.; Filatova, N.; Lindtner, C.; Chi, T.; Degann, S.; Oberlin, D.; Buettner, C. Insulin Receptor Signaling in POMC, but Not AgRP, Neurons Controls Adipose Tissue Insulin Action. Diabetes 2017, 66, 1560–1571. [Google Scholar] [CrossRef]
- Hausen, A.C.; Ruud, J.; Jiang, H.; Hess, S.; Varbanov, H.; Kloppenburg, P.; Brüning, J.C. Insulin-Dependent Activation of MCH Neurons Impairs Locomotor Activity and Insulin Sensitivity in Obesity. Cell Rep. 2016, 17, 2512–2521. [Google Scholar] [CrossRef]
- Bhalla, S.; Mehan, S.; Khan, A.; Rehman, M.U. Protective role of IGF-1 and GLP-1 signaling activation in neurological dysfunctions. Neurosci. Biobehav. Rev. 2022, 142, 104896. [Google Scholar] [CrossRef]
- Shandilya, A.; Mehan, S. Dysregulation of IGF-1/GLP-1 signaling in the progression of ALS: Potential target activators and influences on neurological dysfunctions. Neurol. Sci. 2021, 42, 3145–3166. [Google Scholar] [CrossRef]
- Cignarelli, A.; Genchi, V.A.; Le Grazie, G.; Caruso, I.; Marrano, N.; Biondi, G.; D’Oria, R.; Sorice, G.P.; Natalicchio, A.; Perrini, S.; et al. Mini Review: Effect of GLP-1 Receptor Agonists and SGLT-2 Inhibitors on the Growth Hormone/IGF Axis. Front. Endocrinol. 2022, 13, 846903. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Y.; Cheng, L.; Li, J.; Zhang, T.; Zhao, G.; Zhang, H. Glucagon-Like Peptide-1 Cleavage Product GLP-1(9–36) Reduces Neuroinflammation From Stroke via the Activation of Insulin-Like Growth Factor 1 Receptor in Astrocytes. Eur. J. Pharmacol. 2020, 887, 173581. [Google Scholar] [CrossRef]
- Okada, T.; Mita, Y.; Sakoda, H.; Nakazato, M. Impaired adaptation of energy intake induces severe obesity in aged mice on a high-fat diet. Physiol. Rep. 2019, 7, e13989. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.C.; McKern, N.M.; Ward, C.W. Insulin receptor structure and its implications for the IGF-1 receptor. Curr. Opin. Struct. Biol. 2007, 17, 699–705. [Google Scholar] [CrossRef]
- Björnholm, M.; He, A.R.; Attersand, A.; Lake, S.; Liu, S.C.; Lienhard, G.E.; Taylor, S.; Arner, P.; Zierath, J.R. Absence of functional insulin receptor substrate-3 (IRS-3) gene in humans. Diabetologia 2002, 45, 1697–1702. [Google Scholar] [CrossRef]
- Ling, Y.; Maile, L.A.; Badley-Clarke, J.; Clemmons, D.R. DOK1 mediates SHP-2 binding to the alphaVbeta3 integrin and thereby regulates insulin-like growth factor I signaling in cultured vascular smooth muscle cells. J. Biol. Chem. 2005, 280, 3151–3158. [Google Scholar] [CrossRef]
- Safaroghli-Azar, A.; Sanaei, M.J.; Pourbagheri-Sigaroodi, A.; Bashash, D. Phosphoinositide 3-kinase (PI3K) classes: From cell signaling to endocytic recycling and autophagy. Eur. J. Pharmacol. 2023, 953, 175827. [Google Scholar] [CrossRef]
- Medina-Vera, D.; Navarro, J.A.; Tovar, R.; Rosell-Valle, C.; Gutiérrez-Adan, A.; Ledesma, J.C.; Sanjuan, C.; Pavón, F.J.; Baixeras, E.; Rodríguez de Fonseca, F.; et al. Activation of PI3K/Akt Signaling Pathway in Rat Hypothalamus Induced by an Acute Oral Administration of D-Pinitol. Nutrients 2021, 13, 2268. [Google Scholar] [CrossRef]
- Dibble, C.C.; Cantley, L.C. Regulation of mTORC1 by PI3K signaling. Trends Cell Biol. 2015, 25, 545–555. [Google Scholar] [CrossRef]
- Saltiel, A.R. Insulin signaling in health and disease. J Clin Investig. 2021, 131, e142241. [Google Scholar] [CrossRef]
- Haeusler, R.A.; McGraw, T.E.; Accili, D. Biochemical and cellular properties of insulin receptor signalling. Nat. Rev. Mol. Cell Biol. 2018, 19, 31–44. [Google Scholar] [CrossRef]
- Simcox, J.; Lamming, D.W. The central moTOR of metabolism. Dev. Cell. 2022, 57, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Hakuno, F.; Fukushima, T.; Yoneyama, Y.; Kamei, H.; Ozoe, A.; Yoshihara, H.; Yamanaka, D.; Shibano, T.; Sone-Yonezawa, M.; Yu, B.C.; et al. The Novel Functions of High-Molecular-Mass Complexes Containing Insulin Receptor Substrates in Mediation and Modulation of Insulin-Like Activities: Emerging Concept of Diverse Functions by IRS-Associated Proteins. Front. Endocrinol. 2015, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Yoshihara, H.; Furuta, H.; Kamei, H.; Hakuno, F.; Luan, J.; Duan, C.; Saeki, Y.; Tanaka, K.; Iemura, S.; et al. Nedd4-induced monoubiquitination of IRS-2 enhances IGF signalling and mitogenic activity. Nat. Commun. 2015, 6, 6780. [Google Scholar] [CrossRef]
- Selman, C.; Lingard, S.; Choudhury, A.I.; Batterham, R.L.; Claret, M.; Clements, M.; Ramadani, F.; Okkenhaug, K.; Schuster, E.; Blanc, E.; et al. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. FASEB J. 2008, 22, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Baghdadi, M.; Nespital, T.; Mesaros, A.; Buschbaum, S.; Withers, D.J.; Grönke, S.; Partridge, L. Reduced insulin signaling in neurons induces sex-specific health benefits. Sci. Adv. 2023, 9, eade8137. [Google Scholar] [CrossRef]
- Taguchi, A.; Wartschow, L.M.; White, M.F. Brain IRS2 signaling coordinates life span and nutrient homeostasis. Science 2007, 317, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Selman, C.; Lingard, S.; Gems, D.; Partridge, L.; Withers, D.J. Comment on “Brain IRS2 signaling coordinates life span and nutrient homeostasis”. Science 2008, 320, 1012. [Google Scholar] [CrossRef]
- Valverde, A.M.; Mur, C.; Pons, S.; Alvarez, A.M.; White, M.F.; Kahn, C.R.; Benito, M. Association of insulin receptor substrate 1 (IRS-1) y895 with Grb-2 mediates the insulin signaling involved in IRS-1-deficient brown adipocyte mitogenesis. Mol. Cell Biol. 2001, 21, 2269–2280. [Google Scholar] [CrossRef] [PubMed]
- Russo, B.; Menduni, M.; Borboni, P.; Picconi, F.; Frontoni, S. Autonomic Nervous System in Obesity and Insulin-Resistance-The Complex Interplay between Leptin and Central Nervous System. Int. J. Mol. Sci. 2021, 22, 5187. [Google Scholar] [CrossRef]
- Wen, X.; Zhang, B.; Wu, B.; Xiao, H.; Li, Z.; Li, R.; Xu, X.; Li, T. Signaling pathways in obesity: Mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2022, 7, 298. [Google Scholar] [CrossRef] [PubMed]
- Bathina, S.; Das, U.N. Dysregulation of PI3K-Akt-mTOR pathway in brain of streptozotocin-induced type 2 diabetes mellitus in Wistar rats. Lipids Health Dis. 2018, 17, 168. [Google Scholar] [CrossRef]
- Ma, Y.; Murgia, N.; Liu, Y.; Li, Z.; Sirakawin, C.; Konovalov, R.; Kovzel, N.; Xu, Y.; Kang, X.; Tiwari, A.; et al. Neuronal miR-29a protects from obesity in adult mice. Mol. Metab. 2022, 61, 101507. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Kim, K.W.; Kim, M.S. Leptin signalling pathways in hypothalamic neurons. Cell. Mol. Life Sci. 2016, 73, 1457–1477. [Google Scholar] [CrossRef] [PubMed]
- Benite-Ribeiro, S.A.; Rodrigues, V.A.L.; Machado, M.R.F. Food intake in early life and epigenetic modifications of pro-opiomelanocortin expression in arcuate nucleus. Mol. Biol. Rep. 2021, 48, 3773–3784. [Google Scholar] [CrossRef]
- Williams, K.W.; Margatho, L.O.; Lee, C.E.; Choi, M.; Lee, S.; Scott, M.M.; Elias, C.F.; Elmquist, J.K. Segregation of acute leptin and insulin effects in distinct populations of arcuate proopiomelanocortin neurons. J. Neurosci. 2010, 30, 2472–2479. [Google Scholar] [CrossRef]
- Borges, B.C.; Elias, C.F.; Elias, L.L. PI3K signaling: A molecular pathway associated with acute hypophagic response during inflammatory challenges. Mol. Cell Endocrinol. 2016, 438, 36–41. [Google Scholar] [CrossRef]
- Bharill, P.; Ayyadevara, S.; Alla, R.; Shmookler Reis, R.J. Extreme Depletion of PIP3 Accompanies the Increased Life Span and Stress Tolerance of PI3K-null C. elegans Mutants. Front. Genet. 2013, 4, 34. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Revathidevi, S.; Munirajan, A.K. Akt in cancer: Mediator and more. Semin. Cancer Biol. 2019, 59, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Kim, D.H.; Lee, E.K.; Kim, N.D.; Im, D.S.; Lee, J.; Yu, B.P.; Chung, H.Y. Age-related inflammation and insulin resistance: A review of their intricate interdependency. Arch. Pharm. Res. 2014, 37, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wang, B.; Fei, J.; Santanam, N.; Blough, E.R. Important roles of Akt/PKB signaling in the aging process. Front. Biosci. (Schol. Ed.) 2010, 2, 1169–1188. [Google Scholar] [CrossRef]
- Kim, D.H.; Bang, E.; Ha, S.; Jung, H.J.; Choi, Y.J.; Yu, B.P.; Chung, H.Y. Organ-differential Roles of Akt/FoxOs Axis as a Key Metabolic Modulator during Aging. Aging Dis. 2021, 12, 1713–1728. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tao, X.; Sun, R.; Han, C.; Li, X.; Zhu, Z.; Li, W.; Huang, P.; Gong, W. Cognitive-exercise dual-task intervention ameliorates cognitive decline in natural aging rats via inhibiting the promotion of LncRNA NEAT1/miR-124-3p on caveolin-1-PI3K/Akt/GSK3β Pathway. Brain Res. Bull. 2023, 202, 110761. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Xuanyuan, S.; Zhang, B.; Shi, C. Activation of PI3K/Akt prevents hypoxia/reoxygenation-induced GnRH decline via FOXO3a. Physiol. Res. 2022, 71, 509–516. [Google Scholar] [CrossRef]
- Yang, S.; Du, Y.; Zhao, X.; Wu, C.; Yu, P. Reducing PDK1/Akt Activity: An Effective Therapeutic Target in the Treatment of Alzheimer’s Disease. Cells 2022, 11, 1735. [Google Scholar] [CrossRef]
- Chen, Y.R.; Li, Y.H.; Hsieh, T.C.; Wang, C.M.; Cheng, K.C.; Wang, L.; Lin, T.Y.; Cheung, C.H.A.; Wu, C.L.; Chiang, H. Aging-induced Akt activation involves in aging-related pathologies and Aβ-induced toxicity. Aging Cell 2019, 18, e12989. [Google Scholar] [CrossRef] [PubMed]
- García-San Frutos, M.; Fernández-Agulló, T.; De Solís, A.J.; Andrés, A.; Arribas, C.; Carrascosa, J.M.; Ros, M. Impaired central insulin response in aged Wistar rats: Role of adiposity. Endocrinology 2007, 148, 5238–5247. [Google Scholar] [CrossRef]
- Anfimova, P.A.; Pankrasheva, L.G.; Moiseev, K.Y.; Shirina, E.S.; Porseva, V.V.; Masliukov, P.M. Ontogenetic Changes in the Expression of the Lin28 Protein in the Rat Hypothalamic Tuberal Nuclei. Int. J. Mol. Sci. 2022, 23, 13468. [Google Scholar] [CrossRef]
- Du, S.; Zheng, H. Role of FoxO transcription factors in aging and age-related metabolic and neurodegenerative diseases. Cell Biosci. 2021, 11, 188. [Google Scholar] [CrossRef]
- Shi, C.; Shi, R.; Guo, H. Tumor necrosis factor α reduces gonadotropin-releasing hormone release through increase of forkhead box protein O1 activity. Neuroreport 2020, 31, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, H. Modulation of Sirt1 and FoxO1 on Hypothalamic Leptin-Mediated Sympathetic Activation and Inflammation in Diet-Induced Obese Rats. J. Am. Heart Assoc. 2021, 10, e020667. [Google Scholar] [CrossRef] [PubMed]
- Dakic, T.; Jevdjovic, T.; Djordjevic, J.; Vujovic, P. Short-term fasting differentially regulates PI3K/AkT/mTOR and ERK signalling in the rat hypothalamus. Mech. Ageing Dev. 2020, 192, 111358. [Google Scholar] [CrossRef] [PubMed]
- Papadopoli, D.; Boulay, K.; Kazak, L.; Pollak, M.; Mallette, F.; Topisirovic, I.; Hulea, L. mTOR as a central regulator of lifespan and aging. F1000Research 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Chrienova, Z.; Nepovimova, E.; Kuca, K. The role of mTOR in age-related diseases. J. Enzyme Inhib. Med. Chem. 2021, 36, 1679–1693. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, L.; Qin, S.; Zhang, T.; Yao, J.; Yi, Y.; Deng, L. Mechanistic Target of Rapamycin Complex 1: From a Nutrient Sensor to a Key Regulator of Metabolism and Health. Adv. Nutr. 2022, 13, 1882–1900. [Google Scholar] [CrossRef] [PubMed]
- Saoudaoui, S.; Bernard, M.; Cardin, G.B.; Malaquin, N.; Christopoulos, A.; Rodier, F. mTOR as a senescence manipulation target: A forked road. Adv. Cancer Res. 2021, 150, 335–363. [Google Scholar] [CrossRef]
- Muta, K.; Morgan, D.A.; Rahmouni, K. The role of hypothalamic mTORC1 signaling in insulin regulation of food intake, body weight, and sympathetic nerve activity in male mice. Endocrinology 2015, 156, 1398–1407. [Google Scholar] [CrossRef]
- Caron, A.; Labbé, S.M.; Lanfray, D.; Blanchard, P.G.; Villot, R.; Roy, C.; Sabatini, D.M.; Richard, D.; Laplante, M. Mediobasal hypothalamic overexpression of DEPTOR protects against high-fat diet-induced obesity. Mol. Metab. 2015, 5, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Anfimova, P.A.; Moiseev, K.Y.; Porseva, V.V.; Pankrasheva, L.G.; Masliukov, P.M. mTOR Expression in Neurons of the Rat Tuberal Hypothalamus in Aging. J. Evol. Biochem. Phys. 2022, 58, 1464–1470. [Google Scholar] [CrossRef]
- Panwar, V.; Singh, A.; Bhatt, M.; Tonk, R.K.; Azizov, S.; Raza, A.S.; Sengupta, S.; Kumar, D.; Garg, M. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct Target Ther. 2023, 8, 375. [Google Scholar] [CrossRef]
- Chellappa, K.; Brinkman, J.A.; Mukherjee, S.; Morrison, M.; Alotaibi, M.I.; Carbajal, K.A.; Alhadeff, A.L.; Perron, I.J.; Yao, R.; Purdy, C.S.; et al. Hypothalamic mTORC2 is essential for metabolic health and longevity. Aging Cell 2019, 18, e13014. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Insulin/IGF-1 signaling promotes immunosuppression via the STAT3 pathway: Impact on the aging process and age-related diseases. Inflamm. Res. 2021, 70, 1043–1061. [Google Scholar] [CrossRef]
- Yoshizawa, R.; Umeki, N.; Yamamoto, A.; Murata, M.; Sako, Y. Biphasic spatiotemporal regulation of GRB2 dynamics by p52SHC for transient RAS activation. Biophys. Physicobiol. 2021, 18, 1–12. [Google Scholar] [CrossRef]
- Ullah, R.; Yin, Q.; Snell, A.H.; Wan, L. RAF-MEK-ERK pathway in cancer evolution and treatment. Semin. Cancer Biol. 2022, 85, 123–154. [Google Scholar] [CrossRef]
- Milstein, J.L.; Ferris, H.A. The brain as an insulin-sensitive metabolic organ. Mol. Metab. 2021, 52, 101234. [Google Scholar] [CrossRef]
- Kleinridders, A. Deciphering Brain Insulin Receptor and Insulin-Like Growth Factor 1 Receptor Signalling. J. Neuroendocrinol. 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Metz, M.; O’Hare, J.; Cheng, B.; Puchowicz, M.; Buettner, C.; Scherer, T. Brain insulin signaling suppresses lipolysis in the absence of peripheral insulin receptors and requires the MAPK pathway. Mol. Metab. 2023, 73, 101723. [Google Scholar] [CrossRef]
- Karmarkar, S.W.; Bottum, K.M.; Krager, S.L.; Tischkau, S.A. ERK/MAPK is essential for endogenous neuroprotection in SCN2.2 cells. PLoS ONE 2011, 6, e23493. [Google Scholar] [CrossRef]
- Maher, P.; Dargusch, R.; Bodai, L.; Gerard, P.E.; Purcell, J.M.; Marsh, J.L. ERK activation by the polyphenols fisetin and resveratrol provides neuroprotection in multiple models of Huntington’s disease. Hum. Mol. Genet. 2011, 20, 261–270. [Google Scholar] [CrossRef]
- Zhen, X.; Uryu, K.; Cai, G.; Johnson, G.P.; Friedman, E. Age-associated impairment in brain MAPK signal pathways and the effect of caloric restriction in Fischer 344 rats. J. Gerontol. A Biol. Sci. Med. Sci. 1999, 54, B539–B548. [Google Scholar] [CrossRef]
- Song, G.Y.; Kang, J.S.; Lee, S.Y.; Myung, C.S. Region-specific reduction of Gbeta4 expression and induction of the phosphorylation of PKB/Akt and ERK1/2 by aging in rat brain. Pharmacol. Res. 2007, 56, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.M.; Belsham, D.D. Insulin directly regulates NPY and AgRP gene expression via the MAPK MEK/ERK signal transduction pathway in mHypoE-46 hypothalamic neurons. Mol. Cell Endocrinol. 2009, 307, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, R.; Gabriele, C.; Santamaria, G.; Giuliano, M.; Vella, V.; Massimino, M.; Vigneri, P.; Cuda, G.; Gaspari, M.; Belfiore, A. Comparative proteomic analysis of insulin receptor isoform A and B signaling. Mol Cell Endocrinol. 2022, 557, 111739. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, B.A.; Muñoz, V.R.; Kuga, G.K.; Gaspar, R.C.; Nakandakari, S.C.B.R.; Crisol, B.M.; Botezelli, J.D.; Pauli, L.S.S.; da Silva, A.S.R.; de Moura, L.P.; et al. Obesity Increases Mitogen-Activated Protein Kinase Phosphatase-3 Levels in the Hypothalamus of Mice. Front. Cell Neurosci. 2017, 11, 313. [Google Scholar] [CrossRef]
- Obrietan, K.; Impey, S.; Storm, D.R. Light and circadian rhythmicity regulate MAP kinase activation in the suprachiasmatic nuclei. Nat. Neurosci. 1998, 1, 693–700. [Google Scholar] [CrossRef]
- Alzate-Correa, D.; Aten, S.; Campbell, M.J.; Hoyt, K.R.; Obrietan, K. Light-induced changes in the suprachiasmatic nucleus transcriptome regulated by the ERK/MAPK pathway. PLoS ONE 2021, 16, e0249430. [Google Scholar] [CrossRef]
- Sharma, A.; Anandl, S.K.; Singh, N.; Dwivedi, U.N.; Kakkar, P. AMP-activated protein kinase: An energy sensor and survival mechanism in the reinstatement of metabolic homeostasis. Exp. Cell Res. 2023, 428, 113614. [Google Scholar] [CrossRef]
- Saikia, R.; Joseph, J. AMPK: A key regulator of energy stress and calcium-induced autophagy. J. Mol. Med. 2021, 99, 1539–1551. [Google Scholar] [CrossRef]
- Carling, D. AMPK signalling in health and disease. Curr. Opin. Cell Biol. 2017, 45, 31–37. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Zhou, M.; Chen, C.; Wu, X.; Wang, X. Role of AMPK mediated pathways in autophagy and aging. Biochimie 2022, 195, 100–113. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Age-related changes in AMPK activation: Role for AMPK phosphatases and inhibitory phosphorylation by upstream signaling pathways. Ageing Res. Rev. 2016, 28, 15–26. [Google Scholar] [CrossRef]
- Ning, J.; Xi, G.; Clemmons, D.R. Suppression of AMPK activation via S485 phosphorylation by IGF-I during hyperglycemia is mediated by AKT activation in vascular smooth muscle cells. Endocrinology 2011, 152, 3143–3154. [Google Scholar] [CrossRef]
- Schultze, S.M.; Hemmings, B.A.; Niessen, M.; Tschopp, O. PI3K/AKT, MAPK and AMPK signalling: Protein kinases in glucose homeostasis. Exp. Rev. Mol. Med. 2012, 14, e1. [Google Scholar] [CrossRef] [PubMed]
- Neves, L.D.S.; Oliveira, R.K.G.; Dos Santos, L.S.; Ribeiro, I.O.; Barreto-Medeiros, J.M.B.; Matos, R.J.B. Modulation of hypothalamic AMPK and hypothalamic neuropeptides in the control of eating behavior: A systematic review. Life Sci. 2022, 309, 120947. [Google Scholar] [CrossRef]
- Song, K.; Zhang, Y.; Ga, Q.; Bai, Z.; Ge, R.L. Increased Insulin Sensitivity by High-Altitude Hypoxia in Mice with High-Fat Diet-Induced Obesity Is Associated with Activated AMPK Signaling and Subsequently Enhanced Mitochondrial Biogenesis in Skeletal Muscles. Obes. Facts 2020, 13, 455–472. [Google Scholar] [CrossRef]
- Minokoshi, Y.; Alquier, T.; Furukawa, N.; Kim, Y.B.; Lee, A.; Xue, B.; Mu, J.; Foufelle, F.; Ferré, P.; Birnbaum, M.J.; et al. AMP-kinase regulates food intake by responding to hormonal and nutrient signals in the hypothalamus. Nature 2004, 428, 569–574. [Google Scholar] [CrossRef]
- Claret, M.; Smith, M.A.; Batterham, R.L.; Selman, C.; Choudhury, A.I.; Fryer, L.G.; Clements, M.; Al-Qassab, H.; Heffron, H.; Xu, A.W.; et al. AMPK is essential for energy homeostasis regulation and glucose sensing by POMC and AgRP neurons. J. Clin. Investig. 2007, 117, 2325–2336. [Google Scholar] [CrossRef]
- López, M.; Lage, R.; Saha, A.K.; Pérez-Tilve, D.; Vázquez, M.J.; Varela, L.; Sangiao-Alvarellos, S.; Tovar, S.; Raghay, K.; Rodríguez-Cuenca, S.; et al. Hypothalamic fatty acid metabolism mediates the orexigenic action of ghrelin. Cell Metab. 2008, 7, 389–399. [Google Scholar] [CrossRef]
- Cavaliere, G.; Viggiano, E.; Trinchese, G.; De Filippo, C.; Messina, A.; Monda, V.; Valenzano, A.; Cincione, R.I.; Zammit, C.; Cimmino, F.; et al. Long Feeding High-Fat Diet Induces Hypothalamic Oxidative Stress and Inflammation, and Prolonged Hypothalamic AMPK Activation in Rat Animal Model. Front. Physiol. 2018, 9, 818. [Google Scholar] [CrossRef]
- Toklu, H.Z.; Scarpace, P.J.; Sakarya, Y.; Kirichenko, N.; Matheny, M.; Bruce, E.B.; Carter, C.S.; Morgan, D.; Tümer, N. Intracerebroventricular tempol administration in older rats reduces oxidative stress in the hypothalamus but does not change STAT3 signalling or SIRT1/AMPK pathway. Appl. Physiol. Nutr. Metab. 2017, 42, 59–67. [Google Scholar] [CrossRef]
- Xu, W.; Luo, Y.; Yin, J.; Huang, M.; Luo, F. Targeting AMPK signaling by polyphenols: A novel strategy for tackling aging. Food Funct. 2023, 14, 56–73. [Google Scholar] [CrossRef]
- López, M. Hypothalamic AMPK as a possible target for energy balance-related diseases. Trends Pharmacol. Sci. 2022, 43, 546–556. [Google Scholar] [CrossRef]
- Chau-Van, C.; Gamba, M.; Salvi, R.; Gaillard, R.C.; Pralong, F.P. Metformin inhibits adenosine 5’-monophosphate-activated kinase activation and prevents increases in neuropeptide Y expression in cultured hypothalamic neurons. Endocrinology 2007, 148, 507–511. [Google Scholar] [CrossRef]
- Derkach, K.; Zakharova, I.; Zorina, I.; Bakhtyukov, A.; Romanova, I.; Bayunova, L.; Shpakov, A. The evidence of metabolic-improving effect of metformin in Ay/a mice with genetically-induced melanocortin obesity and the contribution of hypothalamic mechanisms to this effect. PLoS ONE 2019, 14, e0213779. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK-STAT Signaling as a Target for Inflammatory and Autoimmune Diseases: Current and Future Prospects. Drugs 2017, 77, 521–546. [Google Scholar] [CrossRef]
- Dodington, D.W.; Desai, H.R.; Woo, M. JAK/STAT—Emerging Players in Metabolism. Trends Endocrinol. Metab. 2018, 29, 55–65. [Google Scholar] [CrossRef]
- Villarino, A.V.; Kanno, Y.; Ferdinand, J.R.; O’Shea, J.J. Mechanisms of Jak/STAT signaling in immunity and disease. J. Immunol. 2015, 194, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Du, T.; Li, C.; Yang, G. STAT3 phosphorylation in central leptin resistance. Nutr. Metab. 2021, 18, 39. [Google Scholar] [CrossRef]
- Strous, G.J.; Almeida, A.D.S.; Putters, J.; Schantl, J.; Sedek, M.; Slotman, J.A.; Nespital, T.; Hassink, G.C.; Mol, J.A. Growth Hormone Receptor Regulation in Cancer and Chronic Diseases. Front. Endocrinol. 2020, 11, 597573. [Google Scholar] [CrossRef]
- Zieba, D.A.; Biernat, W.; Barć, J. Roles of leptin and resistin in metabolism, reproduction, and leptin resistance. Domest. Anim. Endocrinol. 2020, 106472. [Google Scholar] [CrossRef] [PubMed]
- Scarpace, P.J.; Matheny, M.; Tümer, N. Hypothalamic leptin resistance is associated with impaired leptin signal transduction in aged obese rats. Neuroscience 2001, 104, 1111–1117. [Google Scholar] [CrossRef]
- Michan, S.; Sinclair, D. Sirtuins in mammals: Insights into their biological function. Biochem. J. 2007, 404, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chojdak-Łukasiewicz, J.; Bizoń, A.; Waliszewska-Prosół, M.; Piwowar, A.; Budrewicz, S.; Pokryszko-Dragan, A. Role of Sirtuins in Physiology and Diseases of the Central Nervous System. Biomedicines 2022, 10, 2434. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid Redox Signal 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Lu, C.; Zhao, H.; Liu, Y.; Yang, Z.; Yao, H.; Liu, T.; Gou, T.; Wang, L.; Zhang, J.; Tian, Y.; et al. Novel Role of the SIRT1 in Endocrine and Metabolic Diseases. Int. J. Biol. Sci. 2023, 19, 484–501. [Google Scholar] [CrossRef] [PubMed]
- Ungurianu, A.; Zanfirescu, A.; Margină, D. Sirtuins, resveratrol and the intertwining cellular pathways connecting them. Ageing Res. Rev. 2023, 88, 101936. [Google Scholar] [CrossRef]
- Wu, Q.J.; Zhang, T.N.; Chen, H.H.; Yu, X.F.; Lv, J.L.; Liu, Y.Y.; Liu, Y.S.; Zheng, G.; Zhao, J.Q.; Wei, Y.F.; et al. The sirtuin family in health and disease. Signal Transduct Target Ther. 2022, 7, 402. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhou, M.; Ge, Y.; Wang, X. SIRT1 and aging related signaling pathways. Mech. Ageing Dev. 2020, 187, 111215. [Google Scholar] [CrossRef] [PubMed]
- Pukhalskaia, A.E.; Kvetnoy, I.M.; Linkova, N.S.; Diatlova, A.S.; Gutop, E.O.; Kozlov, K.L.; Paltsev, M.A. Sirtuins and Aging. Neurosci. Behav. Physiol. 2022, 52, 1482–1490. [Google Scholar] [CrossRef]
- Satoh, A.; Imaim, S.I.; Guarente, L. The brain, sirtuins, and ageing. Nat. Rev. Neurosci. 2017, 18, 362–374. [Google Scholar] [CrossRef]
- Mayor, E. Neurotrophic effects of intermittent fasting, calorie restriction and exercise: A review and annotated bibliography. Front. Aging 2023, 4, 1161814. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, S.; Misrani, A.; Huang, H.X.; Zhang, Z.Y.; Li, Q.W.; Long, C. Resveratrol Attenuates Chronic Unpredictable Mild Stress-Induced Alterations in the SIRT1/PGC1α/SIRT3 Pathway and Associated Mitochondrial Dysfunction in Mice. Mol. Neurobiol. 2023, 60, 5102–5116. [Google Scholar] [CrossRef] [PubMed]
- Spirichev, A.A.; Moiseev, K.Y.; Masliukov, P.M. Sirtuin 1 Expression in the Rat Ventromedial and Dorsomedial Hypothalamic Nuclei during Ageing. Bull. Exp. Biol. Med. 2020, 169, 698–700. [Google Scholar] [CrossRef]
- Kukkemane, K.; Jagota, A. Therapeutic effects of hydro-alcoholic leaf extract of Withania somnifera on age-induced changes in daily rhythms of Sirt1, Nrf2 and Rev-erbα in the SCN of male Wistar rats. Biogerontology 2020, 21, 593–607. [Google Scholar] [CrossRef]
- Satoh, A.; Brace, C.S.; Rensing, N.; Cliften, P.; Wozniak, D.F.; Herzog, E.D.; Yamada, K.A.; Imai, S. Sirt1 extends life span and delays aging in mice through the regulation of Nk2 homeobox 1 in the DMH and LH. Cell Metab. 2013, 18, 416–430. [Google Scholar] [CrossRef]
- Sasaki, T.; Kitamura, T. Roles of FoxO1 and Sirt1 in the central regulation of food intake. Endocr. J. 2010, 57, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Carlström, M.; Mirmiran, P.; Ghasemi, A. Nitric oxide: To be or not to be an endocrine hormone? Acta Physiol. 2020, 229, e13443. [Google Scholar] [CrossRef] [PubMed]
- Chachlaki, K.; Prevot, V. Nitric oxide signalling in the brain and its control of bodily functions. Br. J. Pharmacol. 2020, 177, 5437–5458. [Google Scholar] [CrossRef]
- Chachlaki, K.; Garthwaite, J.; Prevot, V. The gentle art of saying NO: How nitric oxide gets things done in the hypothalamus. Nat. Rev. Endocrinol. 2017, 9, 521–535. [Google Scholar] [CrossRef]
- Delli, V.; Silva, M.S.B.; Prévot, V.; Chachlaki, K. The KiNG of reproduction: Kisspeptin/nNOS interactions shaping hypothalamic GnRH release. Mol. Cell. Endocrinol. 2021, 532, 111302. [Google Scholar] [CrossRef] [PubMed]
- Chachlaki, K.; Malone, S.A.; Qualls-Creekmore, E.; Hrabovszky, E.; Münzberg, H.; Giacobini, P.; Ango, F.; Prevot, V. Phenotyping of nNOS neurons in the postnatal and adult female mouse hypothalamus. J. Comp. Neurol. 2017, 525, 3177–3189. [Google Scholar] [CrossRef]
- Moiseev, K.Y.; Vishnyakova, P.A.; Porseva, V.V.; Masliukov, A.P.; Spirichev, A.A.; Emanuilov, A.I.; Masliukov, P.M. Changes of nNOS expression in the tuberal hypothalamic nuclei during ageing. Nitric Oxide 2020, 100–101, 1–6. [Google Scholar] [CrossRef]
- Dawson, T.M.; Dawson, V.L. Nitric Oxide Signaling in Neurodegeneration and Cell Death. Adv. Pharmacol. 2018, 82, 57–83. [Google Scholar] [CrossRef]
- Stepanichev, M.; Aniol, V.; Lazareva, N.; Gulyaeva, N. Decreased Hippocampal Neurogenesis in Aged Male Wistar Rats Is Not Associated with Memory Acquisition in a Water Maze. Int. J. Mol. Sci. 2023, 24, 13276. [Google Scholar] [CrossRef]
- Andrabi, S.M.; Sharma, N.S.; Karan, A.; Shahriar, S.M.S.; Cordon, B.; Ma, B.; Xie, J. Nitric Oxide: Physiological Functions, Delivery, and Biomedical Applications. Adv. Sci. 2023, e2303259. [Google Scholar] [CrossRef]
- Tewari, D.; Sah, A.N.; Bawari, S.; Nabavi, S.F.; Dehpour, A.R.; Shirooie, S.; Braidy, N.; Fiebich, B.L.; Vacca, R.A.; Nabavi, S.M. Role of Nitric Oxide in Neurodegeneration: Function, Regulation, and Inhibition. Curr. Neuropharmacol. 2021, 19, 114–126. [Google Scholar] [CrossRef]
- Carletti, F.; Gambino, G.; Rizzo, V.; Ferraro, G.; Sardo, P. Cannabinoid and nitric oxide signaling interplay in the modulation of hippocampal hyperexcitability: Study on electrophysiological and behavioral models of temporal lobe epilepsy in the rat. Neuroscience 2015, 303, 149–159. [Google Scholar] [CrossRef]
- Belgardt, B.F.; Mauer, J.; Wunderlich, F.T.; Ernst, M.B.; Pal, M.; Spohn, G.; Brönneke, H.S.; Brodesser, S.; Hampel, B.; Schauss, A.C.; et al. Hypothalamic and pituitary c-Jun N-terminal kinase 1 signaling coordinately regulates glucose metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 6028–6033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, G.; Zhang, H.; Karin, M.; Bai, H.; Cai, D. Hypothalamic IKKbeta/NF-kappaB and ER stress link overnutrition to energy imbalance and obesity. Cell 2008, 135, 61–73. [Google Scholar] [CrossRef]
- Zhang, Y.; Reichel, J.M.; Han, C.; Zuniga-Hertz, J.P.; Cai, D. Astrocytic Process Plasticity and IKKβ/NF-κB in Central Control of Blood Glucose, Blood Pressure, and Body Weight. Cell Metab. 2017, 25, 1091–1102.e4. [Google Scholar] [CrossRef] [PubMed]
- González-García, I.; García-Cáceres, C. Hypothalamic Astrocytes as a Specialized and Responsive Cell Population in Obesity. Int. J. Mol. Sci. 2021, 22, 6176. [Google Scholar] [CrossRef] [PubMed]
- McMurphy, T.; Huang, W.; Liu, X.; Siu, J.J.; Queen, N.J.; Xiao, R.; Cao, L. Hypothalamic gene transfer of BDNF promotes healthy aging in mice. Aging Cell 2019, 18, e12846. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Banno, R.; Shibata, M.; Adachi, K.; Hagimoto, S.; Hagiwara, D.; Ozawa, Y.; Goto, M.; Suga, H.; Sugimura, Y.; et al. GABA type B receptor signaling in proopiomelanocortin neurons protects against obesity, insulin resistance, and hypothalamic inflammation in male mice on a high-fat diet. J. Neurosci. 2013, 33, 17166–71713. [Google Scholar] [CrossRef]
- Yu, B.; Cai, D. Neural Programmatic Role of Leptin, TNFα, Melanocortin, and Glutamate in Blood Pressure Regulation vs Obesity-Related Hypertension in Male C57BL/6 Mice. Endocrinology 2017, 158, 1766–1775. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The scheme of orexigenic (AGRP/NPY) and anorexigenic (POMC/CART) ARN neurons and their regulation by circulating hormones. ARN—the arcuate nucleus, AGRP—agouti-related peptide, GLP-1—glucagon-like peptide-1, NPY—neuropeptide Y, POMC—proopiomelanocortin. Red arrows indicate decrease or increase in food intake.
Figure 1.
The scheme of orexigenic (AGRP/NPY) and anorexigenic (POMC/CART) ARN neurons and their regulation by circulating hormones. ARN—the arcuate nucleus, AGRP—agouti-related peptide, GLP-1—glucagon-like peptide-1, NPY—neuropeptide Y, POMC—proopiomelanocortin. Red arrows indicate decrease or increase in food intake.

Figure 2.
The scheme of two main populations of kisspeptin neurons. ARN—the arcuate nucleus, AVPV—anteroventral periventricular nucleus, FSH—follicle stimulating hormone, LH—luteinising hormone.
Figure 2.
The scheme of two main populations of kisspeptin neurons. ARN—the arcuate nucleus, AVPV—anteroventral periventricular nucleus, FSH—follicle stimulating hormone, LH—luteinising hormone.

Figure 3.
The basic scheme of insulin/IGF-1/GH, PI3K)/AKT)/mTOR, MAPK, JAK/STAT, AMPK, and NF-ĸB pathways. AKT—protein kinase B, AMPK—AMP-activated protein kinase, ERK—extracellular signal-regulated kinase, FOXO—forkhead box O, GH—growth hormone, GRB2—growth factor receptor-bound 2, IGF-1 -insulin-like growth factor-1, IKK—inhibitor of nuclear factor kappa B kinase, IRS—insulin receptor substrate, JAK—janus kinase, MEK—mitogen activated protein kinase/ERK kinase, mTOR—the mammalian target of rapamycin, NF-ĸB—nuclear factor kappa-light-chain-enhancer of activated B cells, PDK—phosphoinositol-dependent protein kinase, p85 and p110—subunits of the phosphatidylinositol-4,5-bisphosphate 3-kinase, PIP2—phosphatidylinositol 4,5-bisphosphate, PIP3—phosphatidylinositol-3,4,5-triphosphate, PTEN—phosphatase and tensin homolog deleted on chromosome 10, SIRT—sirtuins, SOCS—suppressor of cytokine signaling, STAT—signal transducer and activator of transcription.
Figure 3.
The basic scheme of insulin/IGF-1/GH, PI3K)/AKT)/mTOR, MAPK, JAK/STAT, AMPK, and NF-ĸB pathways. AKT—protein kinase B, AMPK—AMP-activated protein kinase, ERK—extracellular signal-regulated kinase, FOXO—forkhead box O, GH—growth hormone, GRB2—growth factor receptor-bound 2, IGF-1 -insulin-like growth factor-1, IKK—inhibitor of nuclear factor kappa B kinase, IRS—insulin receptor substrate, JAK—janus kinase, MEK—mitogen activated protein kinase/ERK kinase, mTOR—the mammalian target of rapamycin, NF-ĸB—nuclear factor kappa-light-chain-enhancer of activated B cells, PDK—phosphoinositol-dependent protein kinase, p85 and p110—subunits of the phosphatidylinositol-4,5-bisphosphate 3-kinase, PIP2—phosphatidylinositol 4,5-bisphosphate, PIP3—phosphatidylinositol-3,4,5-triphosphate, PTEN—phosphatase and tensin homolog deleted on chromosome 10, SIRT—sirtuins, SOCS—suppressor of cytokine signaling, STAT—signal transducer and activator of transcription.


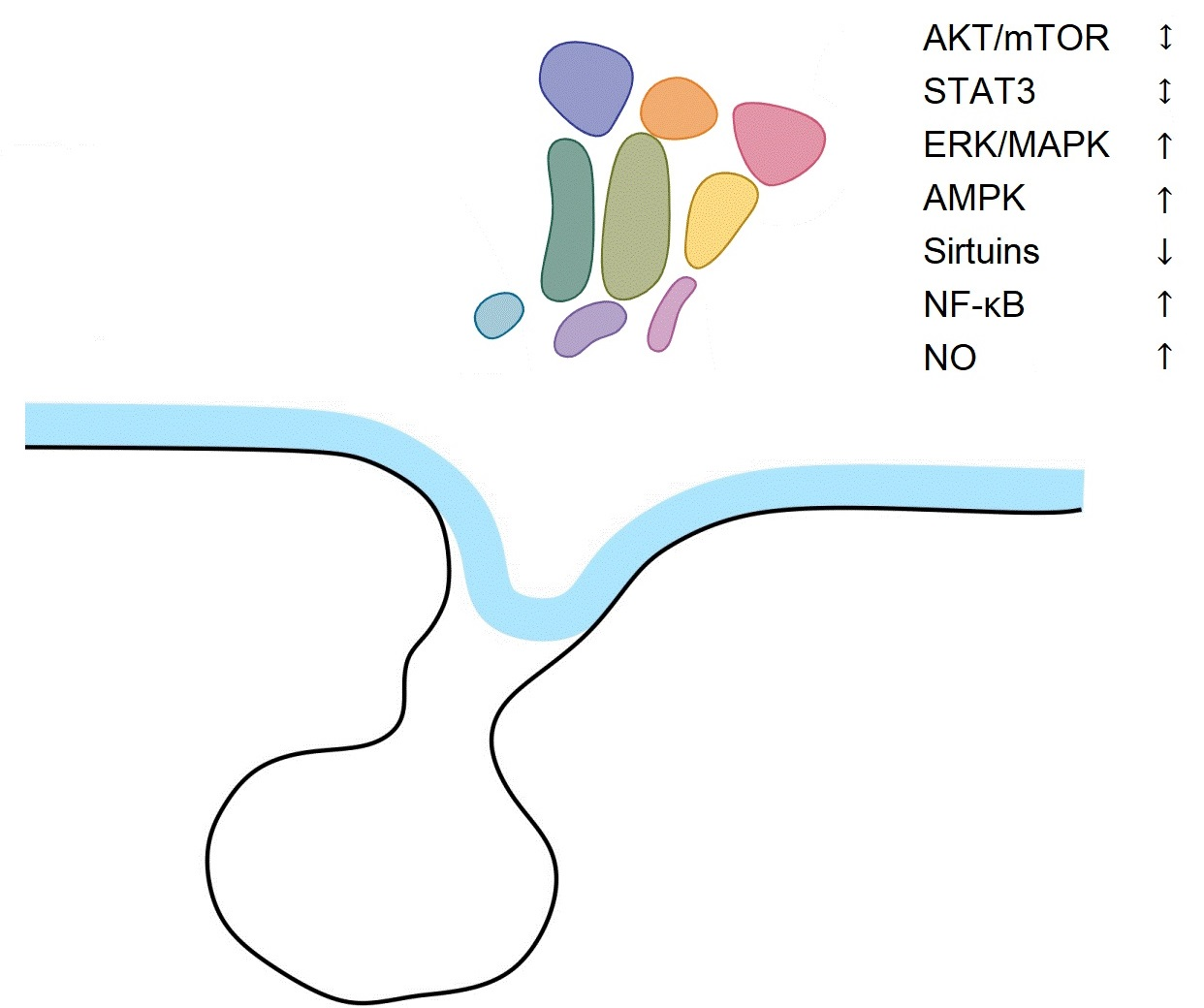{kind=link}
{kind=link}
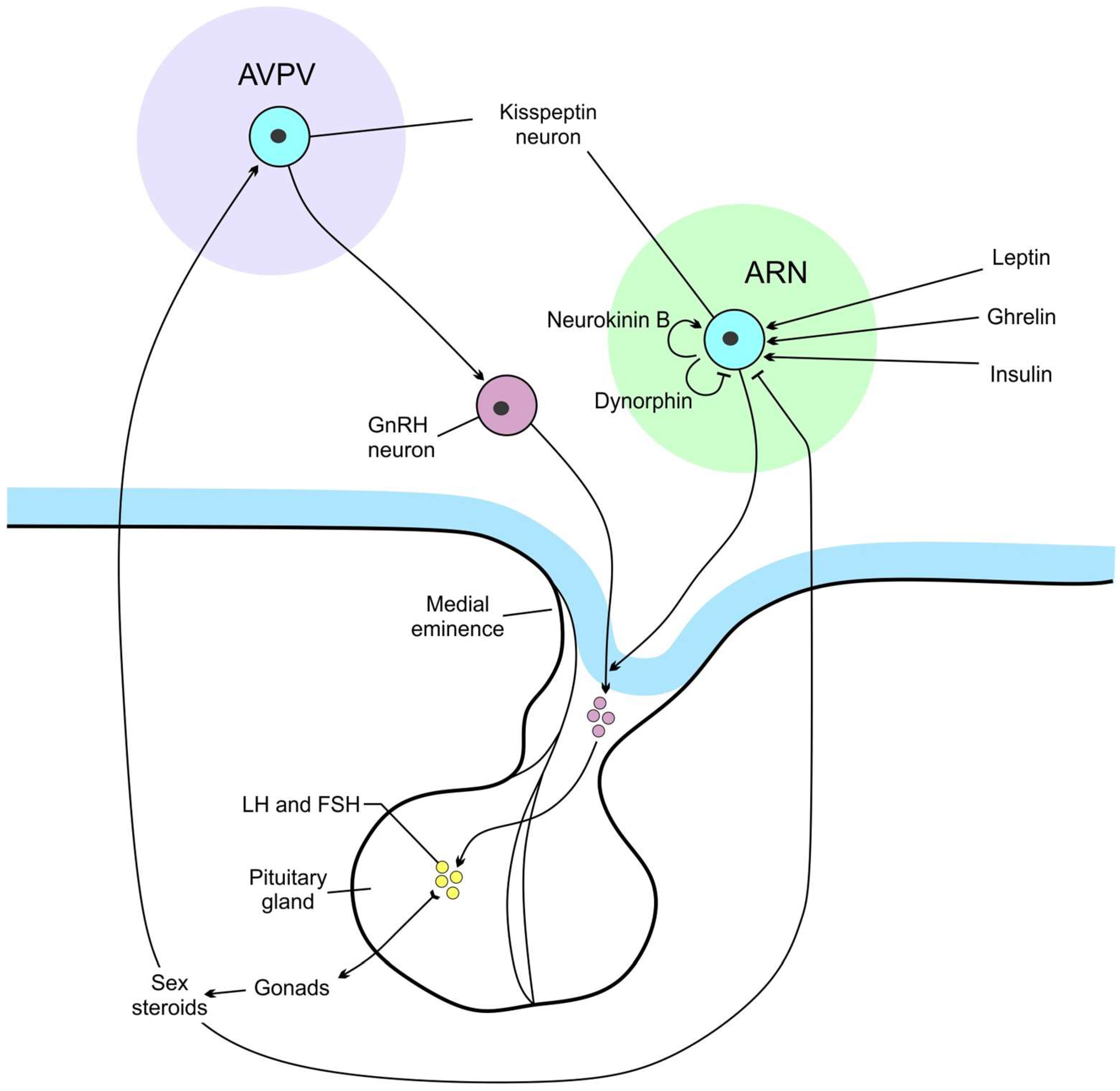{kind=link}
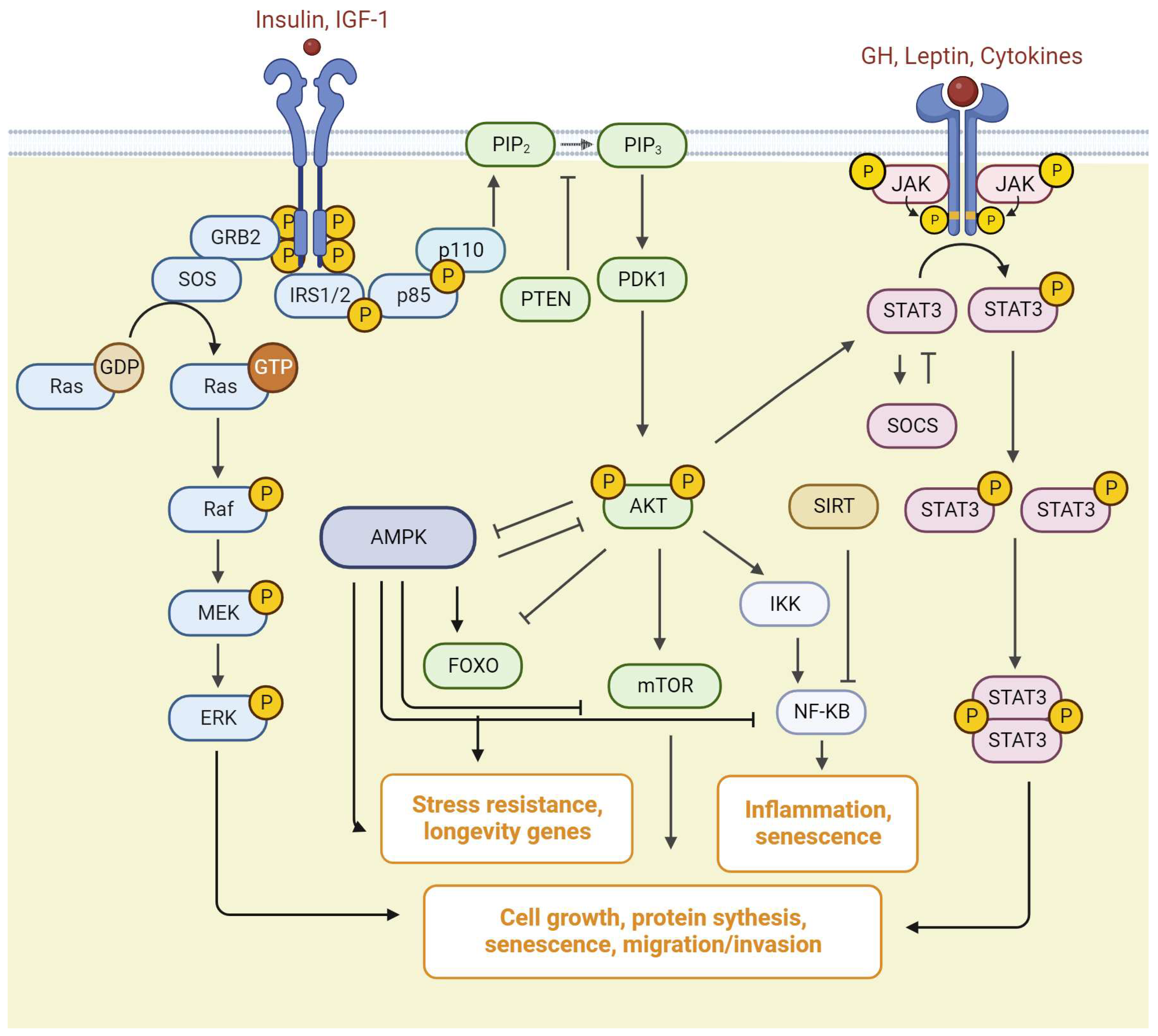{kind=link}
Table 1.
The summary of changes in signaling pathways during aging.
| Signaling Pathway | Aging Tissues | Aging Hypothalamus |
|---|---|---|
| AKT/mTOR | Differential actions in separate organs | No changes |
| STAT3 | No changes | No changes |
| ERK/MAPK | Decreased | Increased |
| AMPK | Decreased | Increased |
| Sirtuins | Decreased | Decreased |
| NF-κB | Increased | Increased |
| NO | Increased | Increased |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Masliukov, P.M. Changes of Signaling Pathways in Hypothalamic Neurons with Aging. Curr. Issues Mol. Biol. 2023, 45, 8289-8308. https://doi.org/10.3390/cimb45100523
AMA Style
Masliukov PM. Changes of Signaling Pathways in Hypothalamic Neurons with Aging. Current Issues in Molecular Biology. 2023; 45(10):8289-8308. https://doi.org/10.3390/cimb45100523
Chicago/Turabian StyleMasliukov, Petr M. 2023. "Changes of Signaling Pathways in Hypothalamic Neurons with Aging" Current Issues in Molecular Biology 45, no. 10: 8289-8308. https://doi.org/10.3390/cimb45100523