
Polyphenols and Small Phenolic Acids as Cellular Metabolic Regulators

Abstract
:1. Introduction
2. Bioavailability of Dietary Polyphenols
3. Bacterial and Host Co-Metabolism of Polyphenols and Phenolic Acid Metabolites
4. Oxidative Stress, Immunomodulatory and Anti-Inflammatory Properties
5. Antipyretic, Antiviral, Antifungal and Analgesic Properties
6. Polyphenols in Obesity
7. Polyphenols in Heart Disease, Cancer, and Type 2 Diabetes
8. Epigenetic Modifications Targeted by Polyphenols
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Obrenovich, M.E.; Nair, N.G.; Beyaz, A.; Aliev, G.; Reddy, V.P. The role of polyphenolic antioxidants in health, disease, and aging. Rejuvenation Res. 2010, 13, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.H.; Moon, Y.J.; Seo, J.E.; Lee, Y.; Kim, K.H.; Chung, J.H. Reactive oxygen species produced by NADPH oxidase, xanthine oxidase, and mitochondrial electron transport system mediate heat shock-induced MMP-1 and MMP-9 expression. Free Radic. Biol. Med. 2008, 44, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lipton, J.M. Eugenol: Antipyretic activity in rabbits. Neuropharmacology 1987, 26, 1775–1778. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladacenco, O.; Roza, E.; Costachescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of Oxidative Stress, Neuroinflammation, and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef] [PubMed]
- Obrenovich, M.; Jaworski, H.; Tadimalla, T.; Mistry, A.; Sykes, L.; Perry, G.; Bonomo, R.A. The Role of the Microbiota-Gut-Brain Axis and Antibiotics in ALS and Neurodegenerative Diseases. Microorganisms 2020, 8, 784. [Google Scholar] [CrossRef]
- Rauscher, F.M.; Sanders, R.A.; Watkins, J.B., 3rd. Effects of isoeugenol on oxidative stress pathways in normal and streptozotocin-induced diabetic rats. J. Biochem. Mol. Toxicol. 2001, 15, 159–164. [Google Scholar] [CrossRef]
- Arfaoui, L. Dietary Plant Polyphenols: Effects of Food Processing on Their Content and Bioavailability. Molecules 2021, 26, 2959. [Google Scholar] [CrossRef]
- Pyo, I.S.; Yun, S.; Yoon, Y.E.; Choi, J.W.; Lee, S.J. Mechanisms of Aging and the Preventive Effects of Resveratrol on Age-Related Diseases. Molecules 2020, 25, 4649. [Google Scholar] [CrossRef]
- Deus, C.M.; Teixeira, J.; Raimundo, N.; Tucci, P.; Borges, F.; Saso, L.; Oliveira, P.J. Modulation of cellular redox environment as a novel therapeutic strategy for Parkinson’s disease. Eur. J. Clin. Investig. 2022, 2022, e13820. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Tima, M.; Polinkovsky, A.; Zhang, R.; Emancipator, S.N.; Donskey, C.J. Targeted Metabolomics Analysis Identifies Intestinal Microbiota-Derived Urinary Biomarkers of Colonization Resistance in Antibiotic-Treated Mice. Antimicrob. Agents Chemother. 2017, 61, e00477-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boer, V.C.; Dihal, A.A.; van der Woude, H.; Arts, I.C.; Wolffram, S.; Alink, G.M.; Rietjens, I.M.; Keijer, J.; Hollman, P.C. Tissue distribution of quercetin in rats and pigs. J. Nutr. 2005, 135, 1718–1725. [Google Scholar] [CrossRef] [PubMed]
- Obrenovich, M.E.; Jaskiw, G.E.; Mana, T.S.C.; Bennett, C.P.; Cadnum, J.; Donskey, C.J. Urinary Metabolites of Green Tea as Potential Markers of Colonization Resistance to Pathogenic Gut Bacteria in Mice. Pathog. Immun. 2019, 4, 271–293. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Nakagawa, T.; Nishioka, H.; Sato, E.; Hirose, A.; Ueno, Y.; Sun, B.; Yokozawa, T.; Nonaka, G. Preparation, characterization, and antioxidative effects of oligomeric proanthocyanidin-L-cysteine complexes. J. Agric. Food Chem. 2007, 55, 1525–1531. [Google Scholar] [CrossRef]
- Kohnert, E.; Kreutz, C.; Binder, N.; Hannibal, L.; Gorkiewicz, G.; Muller, A.; Storz, M.A.; Huber, R.; Lederer, A.K. Changes in Gut Microbiota after a Four-Week Intervention with Vegan vs. Meat-Rich Diets in Healthy Participants: A Randomized Controlled Trial. Microorganisms 2021, 9, 727. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Clifford, M.N. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem. Pharm. 2017, 139, 24–39. [Google Scholar] [CrossRef]
- Borges, G.; Ottaviani, J.I.; van der Hooft, J.J.J.; Schroeter, H.; Crozier, A. Absorption, metabolism, distribution and excretion of (-)-epicatechin: A review of recent findings. Mol. Asp. Med. 2018, 61, 18–30. [Google Scholar] [CrossRef]
- Stalmach, A.; Edwards, C.A.; Wightman, J.D.; Crozier, A. Colonic catabolism of dietary phenolic and polyphenolic compounds from Concord grape juice. Food Funct. 2013, 4, 52–62. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130 (Suppl. 8S), 2073S–2085S. [Google Scholar] [CrossRef]
- Pultz, N.J.; Donskey, C.J. Effect of antibiotic treatment on growth of and toxin production by Clostridium difficile in the cecal contents of mice. Antimicrob. Agents Chemother. 2005, 49, 3529–3532. [Google Scholar] [CrossRef]
- Takagaki, A.; Nanjo, F. Metabolism of (-)-epigallocatechin gallate by rat intestinal flora. J. Agric. Food Chem. 2010, 58, 1313–1321. [Google Scholar] [CrossRef]
- Kutschera, M.; Engst, W.; Blaut, M.; Braune, A. Isolation of catechin-converting human intestinal bacteria. J. Appl. Microbiol. 2011, 111, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. The decomposition of L-tyrosine and its derivatives by Proteus vulgaris. 2. Production of p-hydroxyphenylacetic acid, p-hydroxybenzaldehyde and melanin from L-tyrosine. Bull. Pharm. Res. Inst. 1965, 59, 1–10. [Google Scholar] [PubMed]
- Valerio, F.; Lavermicocca, P.; Pascale, M.; Visconti, A. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef]
- Shaw, W. Increased urinary excretion of a 3-(3-hydroxyphenyl)-3-hydroxypropionic acid (HPHPA), an abnormal phenylalanine metabolite of Clostridia spin the gastrointestinal tract, in urine samples from patients with autism and schizophrenia. Nutr. Neurosci. 2010, 13, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Obrenovich, M.E.; Donskey, C.J.; Schiefer, I.T.; Bongiovanni, R.; Li, L.; Jaskiw, G.E. Quantification of phenolic acid metabolites in humans by LC-MS: A structural and targeted metabolomics approach. Bioanalysis 2018, 10, 1591–1608. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Kumar, J.; Mistry, A.; Sankar Chittoor Mana, T.; Perry, G.; Reddy, V.P.; Obrenovich, M. The Transformative Possibilities of the Microbiota and Mycobiota for Health, Disease, Aging, and Technological Innovation. Biomedicines 2019, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Obrenovich, M.; Reddy, V. Special Issue: Microbiota-Gut-Brain Axis. Microorganisms 2022, 10, 309. [Google Scholar] [CrossRef]
- Ishii, C.; Nakanishi, Y.; Murakami, S.; Nozu, R.; Ueno, M.; Hioki, K.; Aw, W.; Hirayama, A.; Soga, T.; Ito, M.; et al. A Metabologenomic Approach Reveals Changes in the Intestinal Environment of Mice Fed on American Diet. Int. J. Mol. Sci. 2018, 19, 4079. [Google Scholar] [CrossRef]
- Afanas’ev, I.B.; Dorozhko, A.I.; Brodskii, A.V.; Kostyuk, V.A.; Potapovitch, A.I. Chelating and free radical scavenging mechanisms of inhibitory action of rutin and quercetin in lipid peroxidation. Biochem. Pharm. 1989, 38, 1763–1769. [Google Scholar] [CrossRef]
- Saxena, P.; Saxena, A.K.; Cui, X.L.; Obrenovich, M.; Gudipaty, K.; Monnier, V.M. Transition metal-catalyzed oxidation of ascorbate in human cataract extracts: Possible role of advanced glycation end products. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1473–1481. [Google Scholar]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, K.J. Oxidative stress: The paradox of aerobic life. Biochem. Soc. Symp. 1995, 61, 1–31. [Google Scholar] [PubMed]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Herrmann, K.M.; Weaver, L.M. The Shikimate Pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Negreiros de Carvalho, P.L.; Silva Ede, O.; Chagas-Paula, D.A.; Hortolan Luiz, J.H.; Ikegaki, M. Importance and Implications of the Production of Phenolic Secondary Metabolites by Endophytic Fungi: A Mini-Review. Mini Rev. Med. Chem. 2016, 16, 259–271. [Google Scholar] [CrossRef]
- Mir, R.; Jallu, S.; Singh, T. The shikimate pathway: Review of amino acid sequence, function and three-dimensional structures of the enzymes. Crit. Rev. Microbiol. 2015, 41, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Bek, M.K.; Prince, N.Z.; Peralta Marzal, L.N.; Garssen, J.; Perez Pardo, P.; Kraneveld, A.D. The Role of Bacterial-Derived Aromatic Amino Acids Metabolites Relevant in Autism Spectrum Disorders: A Comprehensive Review. Front. Neurosci. 2021, 15, 738220. [Google Scholar] [CrossRef]
- Rajha, H.N.; Paule, A.; Aragones, G.; Barbosa, M.; Caddeo, C.; Debs, E.; Dinkova, R.; Eckert, G.P.; Fontana, A.; Gebrayel, P.; et al. Recent Advances in Research on Polyphenols: Effects on Microbiota, Metabolism, and Health. Mol. Nutr. Food Res. 2022, 66, e2100670. [Google Scholar] [CrossRef]
- Russell, W.; Duthie, G. Plant secondary metabolites and gut health: The case for phenolic acids. Proc. Nutr. Soc. 2011, 70, 389–396. [Google Scholar] [CrossRef]
- Russell, W.R.; Scobbie, L.; Chesson, A.; Richardson, A.J.; Stewart, C.S.; Duncan, S.H.; Drew, J.E.; Duthie, G.G. Anti-inflammatory implications of the microbial transformation of dietary phenolic compounds. Nutr. Cancer 2008, 60, 636–642. [Google Scholar] [CrossRef]
- Gonzales, A.M.; Orlando, R.A. Curcumin and resveratrol inhibit nuclear factor-kappaB-mediated cytokine expression in adipocytes. Nutr. Metab. 2008, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Ak, T.; Gulcin, I. Antioxidant and radical scavenging properties of curcumin. Chem. Biol. Interact. 2008, 174, 27–37. [Google Scholar] [CrossRef]
- Nisar, M.F.; Khadim, M.; Rafiq, M.; Chen, J.; Yang, Y.; Wan, C.C. Pharmacological Properties and Health Benefits of Eugenol: A Comprehensive Review. Oxid. Med. Cell Longev. 2021, 2021, 2497354. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Rahman, U.; Khan, M.R.; Sahar, A.; Mehmood, T.; Khan, M. Essential oil eugenol: Sources, extraction techniques and nutraceutical perspectives. RSC Adv. 2017, 7, 32669–32681. [Google Scholar] [CrossRef]
- Szewczuk, L.M.; Forti, L.; Stivala, L.A.; Penning, T.M. Resveratrol is a peroxidase-mediated inactivator of COX-1 but not COX-2: A mechanistic approach to the design of COX-1 selective agents. J. Biol. Chem. 2004, 279, 22727–22737. [Google Scholar] [CrossRef]
- Barboza, J.N.; da Silva Maia Bezerra Filho, C.; Silva, R.O.; Medeiros, J.V.R.; de Sousa, D.P. An Overview on the Anti-inflammatory Potential and Antioxidant Profile of Eugenol. Oxid. Med. Cell Longev. 2018, 2018, 3957262. [Google Scholar] [CrossRef]
- Mao, T.K.; van de Water, J.; Keen, C.L.; Schmitz, H.H.; Gershwin, M.E. Modulation of TNF-alpha secretion in peripheral blood mononuclear cells by cocoa flavanols and procyanidins. Dev. Immunol. 2002, 9, 135–141. [Google Scholar] [CrossRef]
- Saraiva, R.A.; Araruna, M.K.; Oliveira, R.C.; Menezes, K.D.; Leite, G.O.; Kerntopf, M.R.; Costa, J.G.; Rocha, J.B.; Tome, A.R.; Campos, A.R.; et al. Topical anti-inflammatory effect of Caryocar coriaceum Wittm. (Caryocaraceae) fruit pulp fixed oil on mice ear edema induced by different irritant agents. J. Ethnopharmacol. 2011, 136, 504–510. [Google Scholar] [CrossRef]
- Aliev, G.; Obrenovich, M.E.; Tabrez, S.; Jabir, N.R.; Reddy, V.P.; Li, Y.; Burnstock, G.; Cacabelos, R.; Kamal, M.A. Link between cancer and Alzheimer disease via oxidative stress induced by nitric oxide-dependent mitochondrial DNA overproliferation and deletion. Oxid. Med. Cell Longev. 2013, 2013, 962984. [Google Scholar] [CrossRef]
- Vidhya, N.; Devaraj, S.N. Induction of apoptosis by eugenol in human breast cancer cells. Indian J. Exp. Biol. 2011, 49, 871–878. [Google Scholar]
- Bendre, R.S.; Rajput, J. Outlooks on medicinal properties of eugenol and its synthetic derivatives. Nat. Prod. Chem. Res. 2016, 4, 2. [Google Scholar] [CrossRef]
- Han, D.; Ryu, J.-Y.; Lee, H.; Hur, H.-G. Bacterial Biotransformation of Phenylpropanoid Compounds for Producing Flavor and Fragrance Compounds. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 125–133. [Google Scholar] [CrossRef]
- Qi, L.; Mao, H.; Lu, X.; Shi, T.; Wang, J. Cinnamaldehyde Promotes the Intestinal Barrier Functions and Reshapes Gut Microbiome in Early Weaned Rats. Front. Nutr. 2021, 8, 748503. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- Benencia, F.; Courreges, M.C. In vitro and in vivo activity of eugenol on human herpesvirus. Phytother. Res. 2000, 14, 495–500. [Google Scholar] [CrossRef]
- Palamara, A.T.; Perno, C.F.; Ciriolo, M.R.; Dini, L.; Balestra, E.; D’Agostini, C.; di Francesco, P.; Favalli, C.; Rotilio, G.; Garaci, E. Evidence for antiviral activity of glutathione: In vitro inhibition of herpes simplex virus type 1 replication. Antiviral. Res. 1995, 27, 237–253. [Google Scholar] [CrossRef]
- Jadhav, B.K.; Khandelwal, K.R.; Ketkar, A.R.; Pisal, S.S. Formulation and evaluation of mucoadhesive tablets containing eugenol for the treatment of periodontal diseases. Drug Dev. Ind. Pharm. 2004, 30, 195–203. [Google Scholar] [CrossRef]
- Sarrami, N.; Pemberton, M.N.; Thornhill, M.H.; Theaker, E.D. Adverse reactions associated with the use of eugenol in dentistry. B.r Dent. J. 2002, 193, 257–259. [Google Scholar] [CrossRef]
- Chung, G.; Rhee, J.N.; Jung, S.J.; Kim, J.S.; Oh, S.B. Modulation of CaV2.3 calcium channel currents by eugenol. J. Dent. Res. 2008, 87, 137–141. [Google Scholar] [CrossRef]
- Lee, S.J.; Han, J.I.; Lee, G.S.; Park, M.J.; Choi, I.G.; Na, K.J.; Jeung, E.B. Antifungal effect of eugenol and nerolidol against Microsporum gypseum in a guinea pig model. Biol. Pharm. Bull. 2007, 30, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Akdeniz, E.; Yakisik, E.; Rasouli Pirouzian, H.; Akkin, S.; Turan, B.; Tipigil, E.; Toker, O.S.; Ozcan, O. Carob powder as cocoa substitute in milk and dark compound chocolate formulation. J. Food Sci. Technol. 2021, 58, 4558–4566. [Google Scholar] [CrossRef] [PubMed]
- Kumazawa, S.; Taniguchi, M.; Suzuki, Y.; Shimura, M.; Kwon, M.S.; Nakayama, T. Antioxidant activity of polyphenols in carob pods. J. Agric. Food Chem. 2002, 50, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Norikura, T.; Matsui-Yuasa, I.; Kumazawa, S.; Honda, S.; Sonoda, T.; Kojima-Yuasa, A. Carob pod polyphenols suppress the differentiation of adipocytes through posttranscriptional regulation of C/EBPbeta. PLoS ONE 2021, 16, e0248073. [Google Scholar] [CrossRef]
- Goody, D.; Pfeifer, A. MicroRNAs in brown and beige fat. Biochim. Biophys Acta Mol. Cell Biol. Lipids 2019, 1864, 29–36. [Google Scholar] [CrossRef]
- Silvester, A.J.; Aseer, K.R.; Yun, J.W. Dietary polyphenols and their roles in fat browning. J. Nutr. Biochem. 2019, 64, 1–12. [Google Scholar] [CrossRef]
- Merlin, J.; Evans, B.A.; Dehvari, N.; Sato, M.; Bengtsson, T.; Hutchinson, D.S. Could burning fat start with a brite spark? Pharmacological and nutritional ways to promote thermogenesis. Mol. Nutr. Food Res. 2016, 60, 18–42. [Google Scholar] [CrossRef]
- Mizunoya, W.; Miyahara, H.; Okamoto, S.; Akahoshi, M.; Suzuki, T.; Do, M.K.; Ohtsubo, H.; Komiya, Y.; Lan, M.; Waga, T.; et al. Improvement of Endurance Based on Muscle Fiber-Type Composition by Treatment with Dietary Apple Polyphenols in Rats. PLoS ONE 2015, 10, e0134303. [Google Scholar] [CrossRef]
- Tamura, Y.; Tomiya, S.; Takegaki, J.; Kouzaki, K.; Tsutaki, A.; Nakazato, K. Apple polyphenols induce browning of white adipose tissue. J. Nutr. Biochem. 2020, 77, 108299. [Google Scholar] [CrossRef]
- Arranz, S.; Chiva-Blanch, G.; Valderas-Martinez, P.; Medina-Remon, A.; Lamuela-Raventos, R.M.; Estruch, R. Wine, beer, alcohol and polyphenols on cardiovascular disease and cancer. Nutrients 2012, 4, 759–781. [Google Scholar] [CrossRef]
- Trost, K.; Klancnik, A.; Mozetic Vodopivec, B.; Sternad Lemut, M.; Jug Novsak, K.; Raspor, P.; Smole Mozina, S. Polyphenol, antioxidant and antimicrobial potential of six different white and red wine grape processing leftovers. J. Sci. Food Agric. 2016, 96, 4809–4820. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Chittoor Mana, T.; Rai, H.; Dorjee Shola, D.; Christopher Sass, C.; McCloskey, B.; Bruce, L. Recent findings within the microbiota–gut–brain– endocrine metabolic interactome. Pathol. Lab. Med. Int. 2017, 2017, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Hernández, T.; Estrella, I.; Dueñas, M.; de Simón, B.F.; Cadahía, E. Influence of wood origin in the polyphenolic composition of a Spanish red wine aging in bottle, after storage in barrels of Spanish, French and American oak wood. Eur. Food Res. Technol. 2007, 224, 695–705. [Google Scholar] [CrossRef]
- Reinisalo, M.; Karlund, A.; Koskela, A.; Kaarniranta, K.; Karjalainen, R.O. Polyphenol Stilbenes: Molecular Mechanisms of Defence against Oxidative Stress and Aging-Related Diseases. Oxid. Med. Cell Longev. 2015, 2015, 340520. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clement, C.; Courot, E. Biosynthesis, metabolism, molecular engineering, and biological functions of stilbene phytoalexins in plants. Biofactors 2010, 36, 331–341. [Google Scholar] [CrossRef]
- Cottart, C.H.; Nivet-Antoine, V.; Beaudeux, J.L. Review of recent data on the metabolism, biological effects, and toxicity of resveratrol in humans. Mol. Nutr. Food Res. 2014, 58, 7–21. [Google Scholar] [CrossRef]
- Ma, J.; Zheng, Y.; Tang, W.; Yan, W.; Nie, H.; Fang, J.; Liu, G. Dietary polyphenols in lipid metabolism: A role of gut microbiome. Anim. Nutr. 2020, 6, 404–409. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Obrenovich, M.; Siddiqui, B.; McCloskey, B.; Reddy, V.P. The Microbiota-Gut-Brain Axis Heart Shunt Part I: The French Paradox, Heart Disease and the Microbiota. Microorganisms 2020, 8, 490. [Google Scholar] [CrossRef]
- Obrenovich, M.; Tabrez, S.; Siddiqui, B.; McCloskey, B.; Perry, G. The Microbiota-Gut-Brain Axis-Heart Shunt Part II: Prosaic Foods and the Brain-Heart Connection in Alzheimer Disease. Microorganisms 2020, 8, 493. [Google Scholar] [CrossRef] [PubMed]
- Brunt, V.E.; Casso, A.G.; Gioscia-Ryan, R.A.; Sapinsley, Z.J.; Ziemba, B.P.; Clayton, Z.S.; Bazzoni, A.E.; VanDongen, N.S.; Richey, J.J.; Hutton, D.A.; et al. Gut Microbiome-Derived Metabolite Trimethylamine N-Oxide Induces Aortic Stiffening and Increases Systolic Blood Pressure With Aging in Mice and Humans. Hypertension 2021, 78, 499–511. [Google Scholar] [CrossRef]
- Roychowdhury, S.; Cadnum, J.; Glueck, B.; Obrenovich, M.; Donskey, C.; Cresci, G.A.M. Faecalibacterium prausnitzii and a Prebiotic Protect Intestinal Health in a Mouse Model of Antibiotic and Clostridium difficile Exposure. JPEN J. Parenter Enter. Nutr. 2018, 42, 1156–1167. [Google Scholar] [CrossRef]
- Niwano, Y.; Kohzaki, H.; Shirato, M.; Shishido, S.; Nakamura, K. Putative Mechanisms Underlying the Beneficial Effects of Polyphenols in Murine Models of Metabolic Disorders in Relation to Gut Microbiota. Curr. Issues Mol. Biol. 2022, 44, 1353–1375. [Google Scholar] [CrossRef]
- Obrenovich, M.E.M. Leaky Gut, Leaky Brain? Microorganisms 2018, 6, 107. [Google Scholar] [CrossRef]
- Kaur, N.; Chugh, H.; Tomar, V.; Sakharkar, M.K.; Dass, S.K.; Chandra, R. Cinnamon attenuates adiposity and affects the expression of metabolic genes in diet-induced obesity model of zebrafish. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2930. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Minghetti, L.; D’Archivio, M. Interaction between Gut Microbiota and Curcumin: A New Key of Understanding for the Health Effects of Curcumin. Nutrients 2020, 12, 2499. [Google Scholar] [CrossRef]
- McFadden, R.M.; Larmonier, C.B.; Shehab, K.W.; Midura-Kiela, M.; Ramalingam, R.; Harrison, C.A.; Besselsen, D.G.; Chase, J.H.; Caporaso, J.G.; Jobin, C.; et al. The Role of Curcumin in Modulating Colonic Microbiota During Colitis and Colon Cancer Prevention. Inflamm. Bowel. Dis. 2015, 21, 2483–2494. [Google Scholar] [CrossRef]
- Bogusławska-Tryk, M.; Szymeczko, R.; Piotrowska, A.; Burlikowska, K.; Śliżewska, K. Ileal and cecal microbial population and short-chain fatty acid profile in broiler chickens fed diets supplemented with lignocellulose. Pak. Vet. J. 2015, 35, 212–216. [Google Scholar]
- Greiner, A.K.; Papineni, R.V.; Umar, S. Chemoprevention in gastrointestinal physiology and disease. Natural products and microbiome. Am. J. Physiol. Gastrointest. Liver. Physiol. 2014, 307, G1–G15. [Google Scholar] [CrossRef]
- Cheng, B.; Gong, H.; Xiao, H.; Petersen, R.B.; Zheng, L.; Huang, K. Inhibiting toxic aggregation of amyloidogenic proteins: A therapeutic strategy for protein misfolding diseases. Biochim. Biophys. Acta 2013, 1830, 4860–4871. [Google Scholar] [CrossRef]
- Obrenovich, M.; Monnier, V. Vitamin B1 Blocks Damage Caused by Hyperglycemia. Sci. Aging Knowl. Environ. 2003, pe6. [Google Scholar] [CrossRef] [PubMed]
- Portela, A.; Esteller, M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.U.; Hassan, F.; Rehman, M.; Khan, M.S.; Ali, M.A.; Javed, A.; Nawaz, A.; Yang, C. Curcumin as an Alternative Epigenetic Modulator: Mechanism of Action and Potential Effects. Front. Genet. 2019, 10, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Cui, L.; Ahmed, S.; Zainab, F.; Wu, Q.; Wang, X.; Yuan, Z. An overview of epigenetic agents and natural nutrition products targeting DNA methyltransferase, histone deacetylases and microRNAs. Food Chem. Toxicol. 2019, 123, 574–594. [Google Scholar] [CrossRef]
- Ogawa, O.; Zhu, X.; Lee, H.G.; Raina, A.; Obrenovich, M.E.; Bowser, R.; Ghanbari, H.A.; Castellani, R.J.; Perry, G.; Smith, M.A. Ectopic localization of phosphorylated histone H3 in Alzheimer’s disease: A mitotic catastrophe? Acta Neuropathol. 2003, 105, 524–528. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Rai, H.; Chittoor Mana, T.; Dorjee Shola, D.; Christopher Sass, C.; McCloskey, B.; Levison, B. Dietary Co-Metabolism within the Microbiota-Gut-Brain-Endocrine Metabolic Interactome. BAO Microbiol. 2017, 2, 22. [Google Scholar]
- Jaskiw, G.E.; Obrenovich, M.E.; Donskey, C.J. The phenolic interactome and gut microbiota: Opportunities and challenges in developing applications for schizophrenia and autism. Psychopharmacology 2019, 236, 1471–1489. [Google Scholar] [CrossRef]




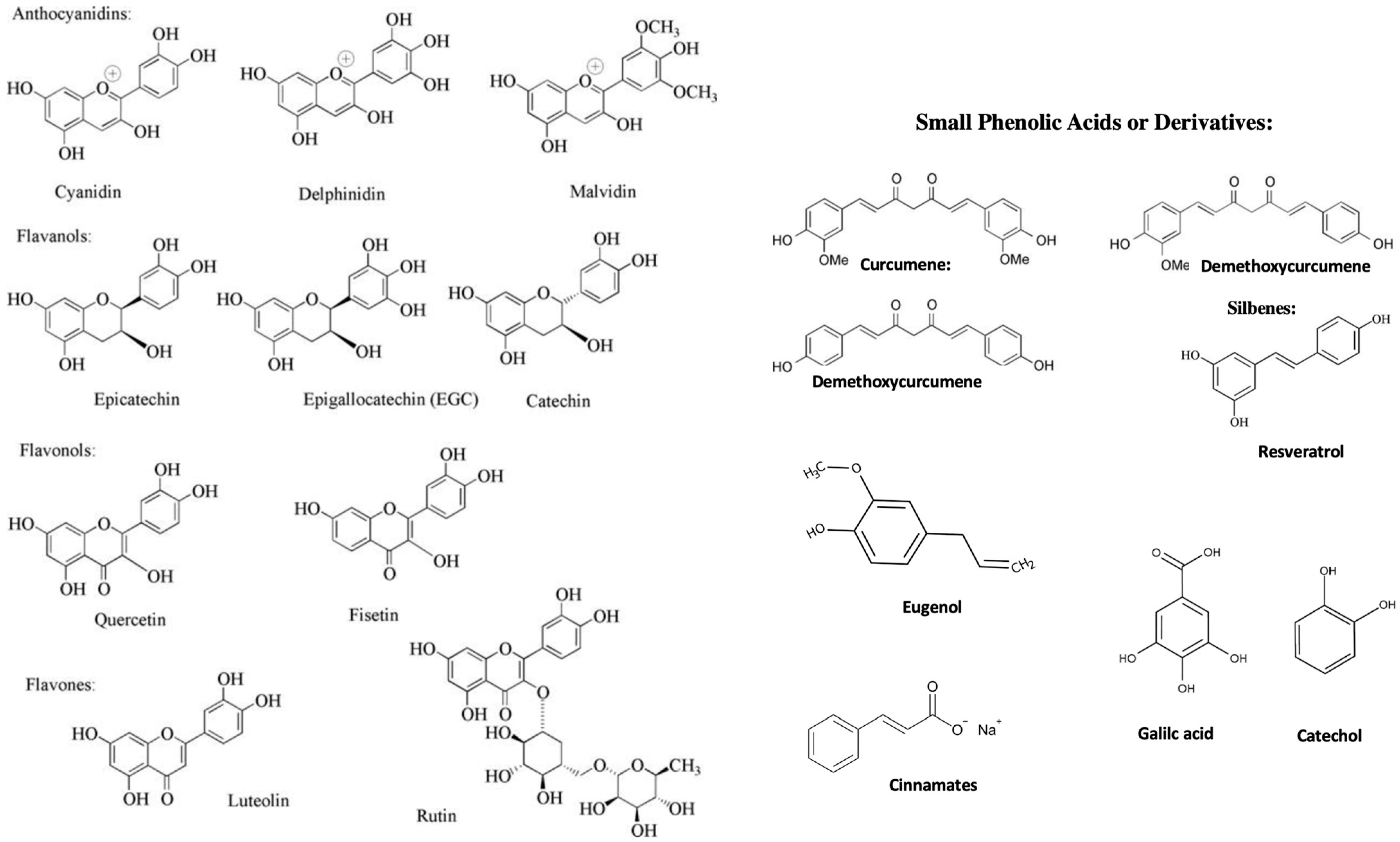{kind=link}
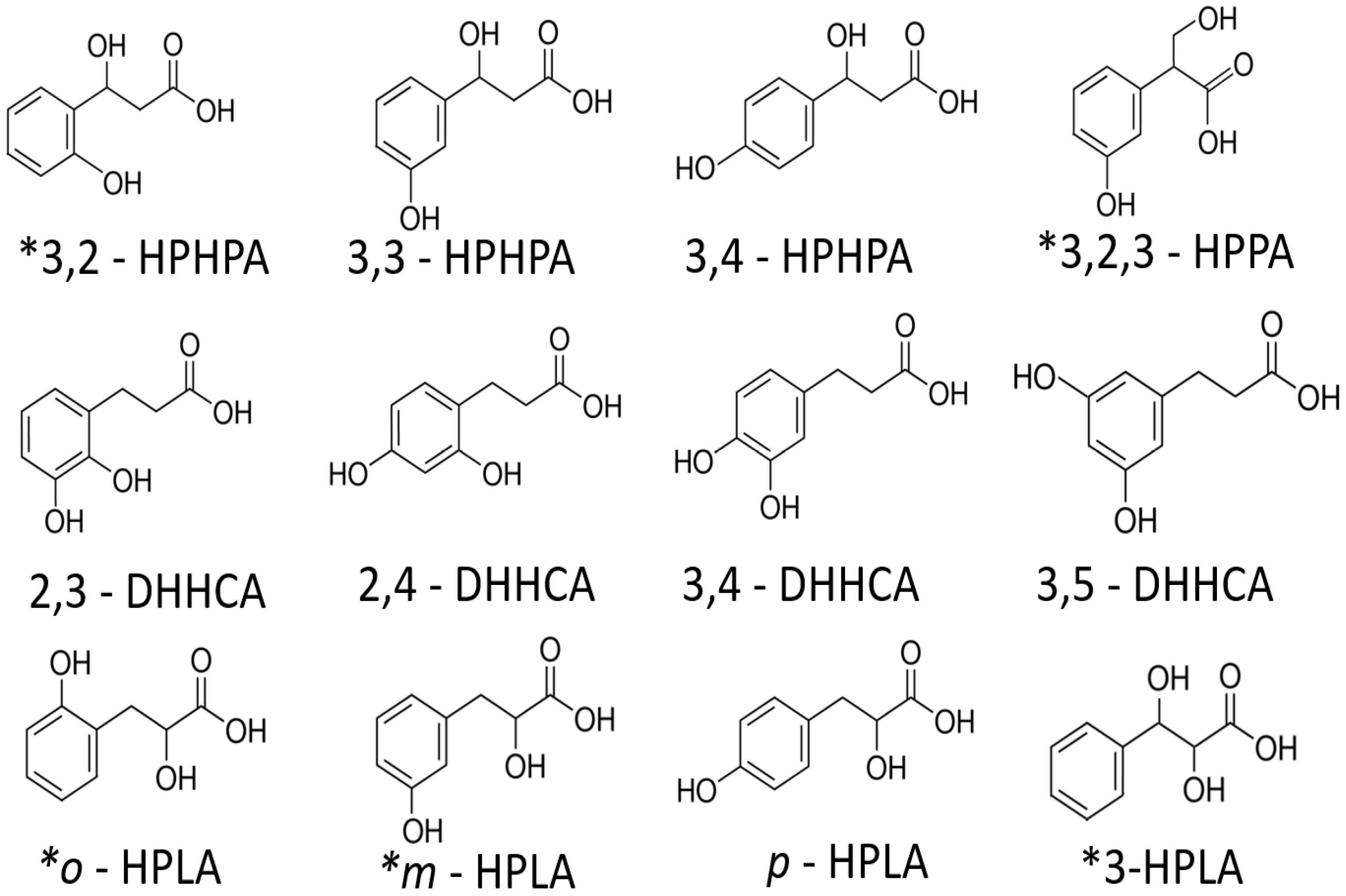{kind=link}
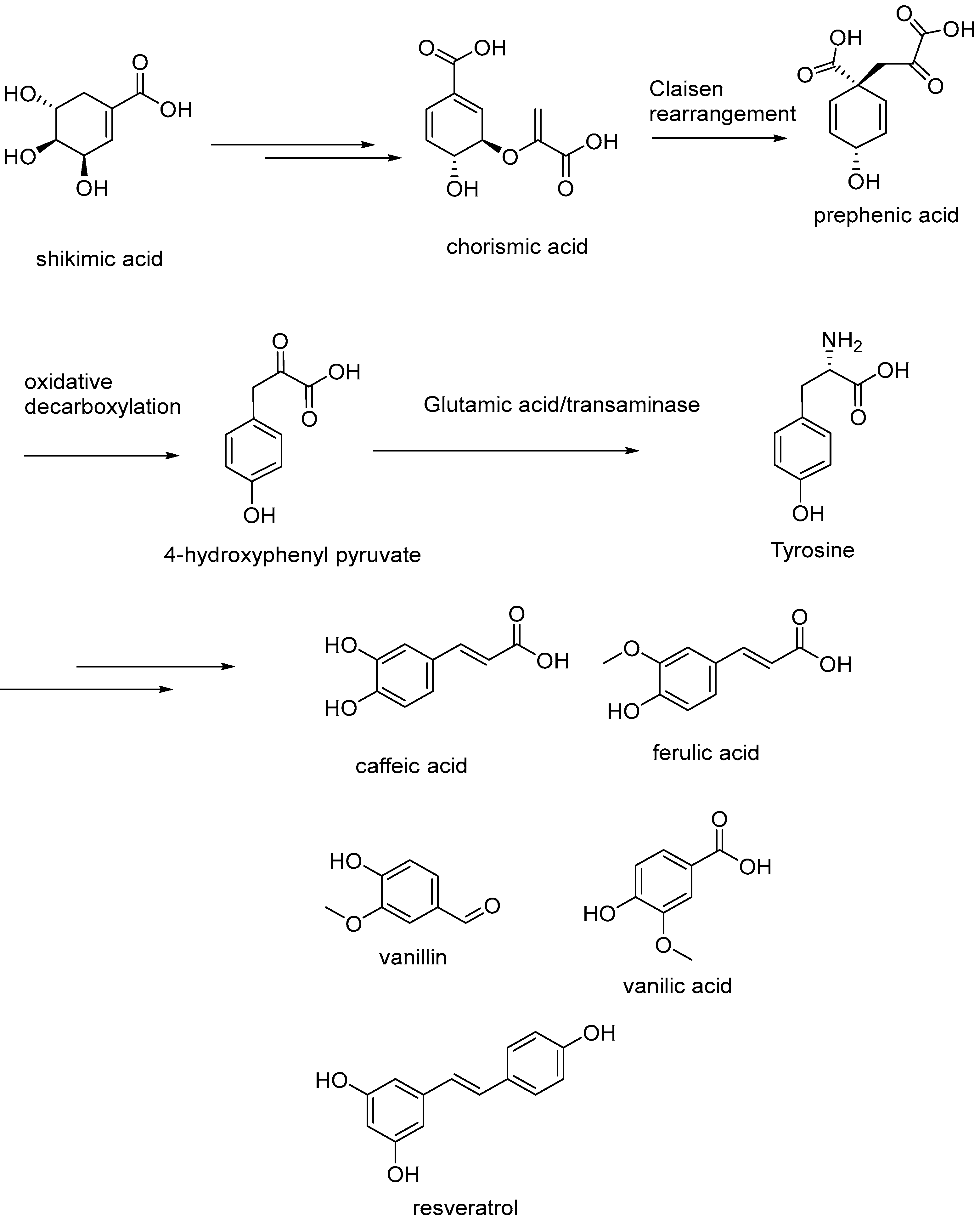{kind=link}
| Polyphenols | Sources | Chemical or Physiological Functions | Colonizing Bacteria |
|---|---|---|---|
| Curcuminoids | Dietary, Plants, such as: Curcuma longa | Shikimate pathway intermediate inhibits NF-kB, TNF-a, IL-1b, IL-6, and COX-2 gene expression, anti-inflammatory activity | Lactobacilli sp. Prevotella sp., Many uncharacterized species |
| (-)-epigallocatechin-3-O-gallate | Dietary, Plants, such as: Green tea (Camellia sinensis) | Shikimate pathway intermediate, chelating properties, anti-inflammatory and oxidative stress activity | Clostridium orbiscindens, Enterobacter aerogenes, Raoultella planticola, Bifidobacterium longum, Eubacterium ramulus in |
| (-)-epicatechin-3-O-gallate | Dietary, Plants, such as: green tea (Camellia sinensis) | Shikimate pathway intermediate, chelating properties, anti-inflammatory and oxidative stress activity | Clostridium orbiscindens, Enterobacter aerogenes, Raoultella planticola, Bifidobacterium longum, Eubacterium ramulus in |
| (-)-epigallocatechin | Dietary, Plants, such as: green tea (Camellia sinensis) | Shikimate pathway intermediate, chelating properties, anti-inflammatory and anti-oxidative stress activity | Clostridium orbiscindens, Enterobacter aerogenes, Raoultella planticola, Bifidobacterium longum, Eubacterium ramulus in |
| (-)-epicatechin | Dietary, Plants, such as: green tea (Camellia sinensis) | Shikimate pathway intermediate, chelating properties, anti-inflammatory and oanti-xidative stress activity | Clostridium orbiscindens, Enterobacter aerogenes, Raoultella planticola, Bifidobacterium longum, Eubacterium ramulus in |
| Cinnamon | Dietary, plants, such as: bark of Cinnamomum versum | Shikimate pathway intermediate, chelating properties, anti-inflammatory and anti-oxidative stress activity | Enterococcus spp. and Lactobacillus spp., Campylobacter spp. and Enterococcus spp. kkermansia, Bacteroides, Clostridium III, Psychrobacter |
| Eugenol | Dietary, Plants, such as: Eugenia caryophyllata | anti-inflammatory activity, decreased proinflammatory cytokines, such as TNF-α, NF-κB, synthesis of ferulic acid and other aromatic compounds via shikimate pathway through enzymes phenylalanine ammonia lyase; tyrosine ammonia lyase; S-adenosyl methionine (methyl donor). | Many uncharacterized species, Staphylococcus aureus, Pseudomonas aeruginosa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obrenovich, M.; Li, Y.; Tayahi, M.; Reddy, V.P. Polyphenols and Small Phenolic Acids as Cellular Metabolic Regulators. Curr. Issues Mol. Biol. 2022, 44, 4152-4166. https://doi.org/10.3390/cimb44090285
Obrenovich M, Li Y, Tayahi M, Reddy VP. Polyphenols and Small Phenolic Acids as Cellular Metabolic Regulators. Current Issues in Molecular Biology. 2022; 44(9):4152-4166. https://doi.org/10.3390/cimb44090285
Chicago/Turabian StyleObrenovich, Mark, Yi Li, Moncef Tayahi, and V. Prakash Reddy. 2022. "Polyphenols and Small Phenolic Acids as Cellular Metabolic Regulators" Current Issues in Molecular Biology 44, no. 9: 4152-4166. https://doi.org/10.3390/cimb44090285