
Multifunctionality of Clausena harmandiana Extract and Its Active Constituents against Alzheimer’s Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
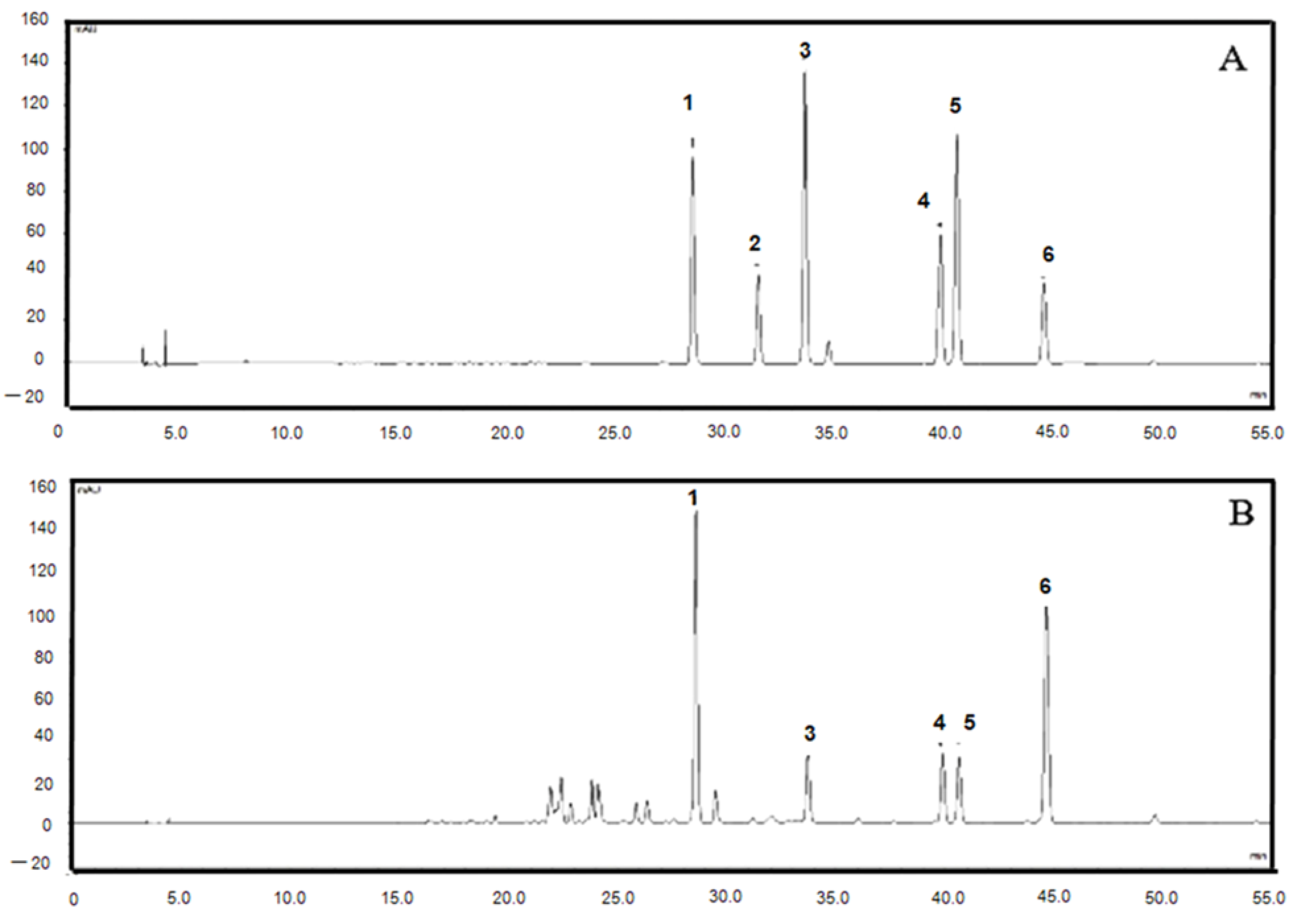
2.1. HPLC Fingerprints of the Root Extract of C. harmandiana
2.2. In Vitro Antioxidant-Activity Evaluations
2.3. In Vitro AChE-Inhibitory Activity Determination
2.4. Investigation of Aβ-Aggregation Inhibition
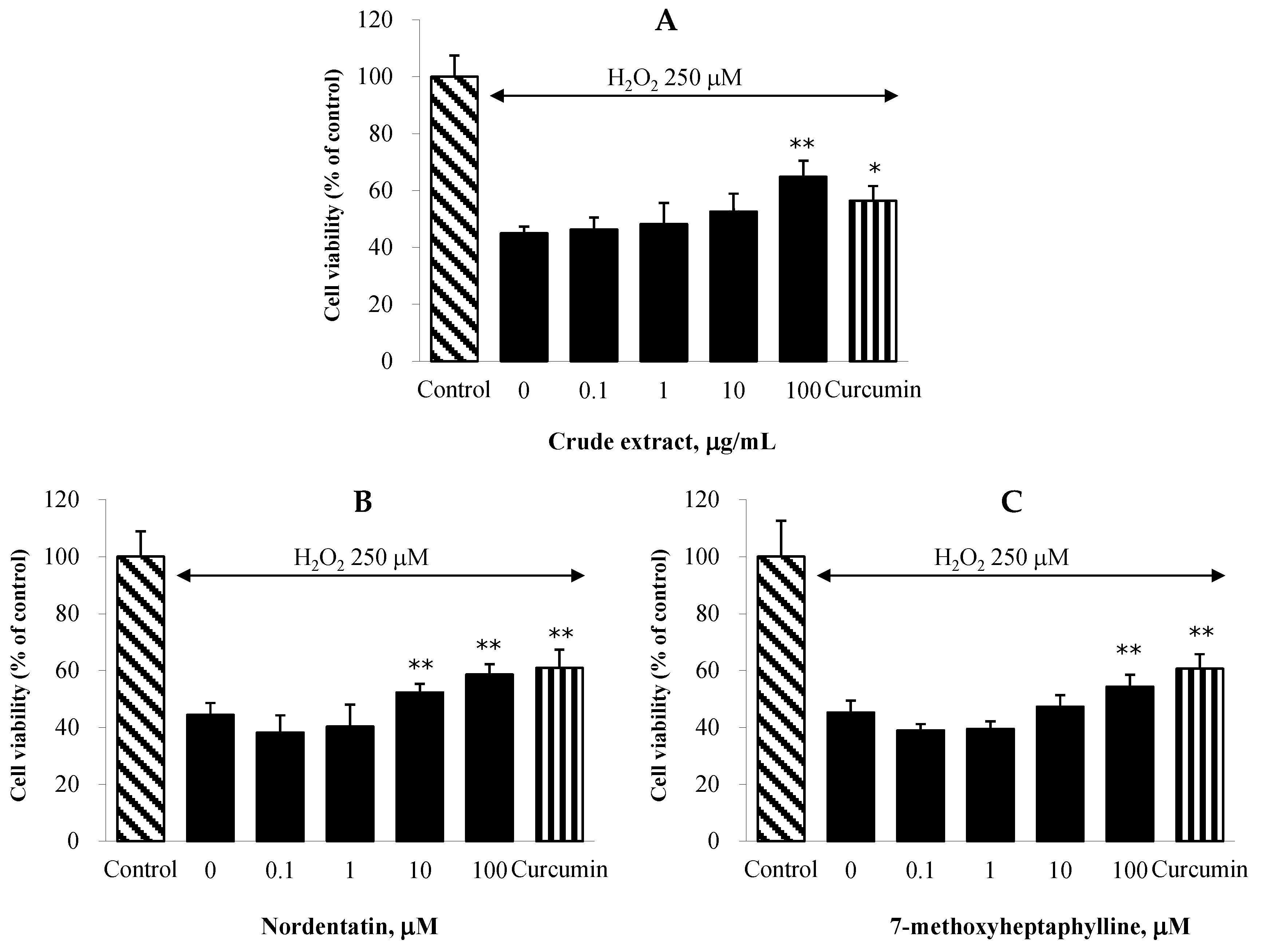
2.5. Effect on H2O2-Induced Neuronal Cell Death in NG108-15 Cells
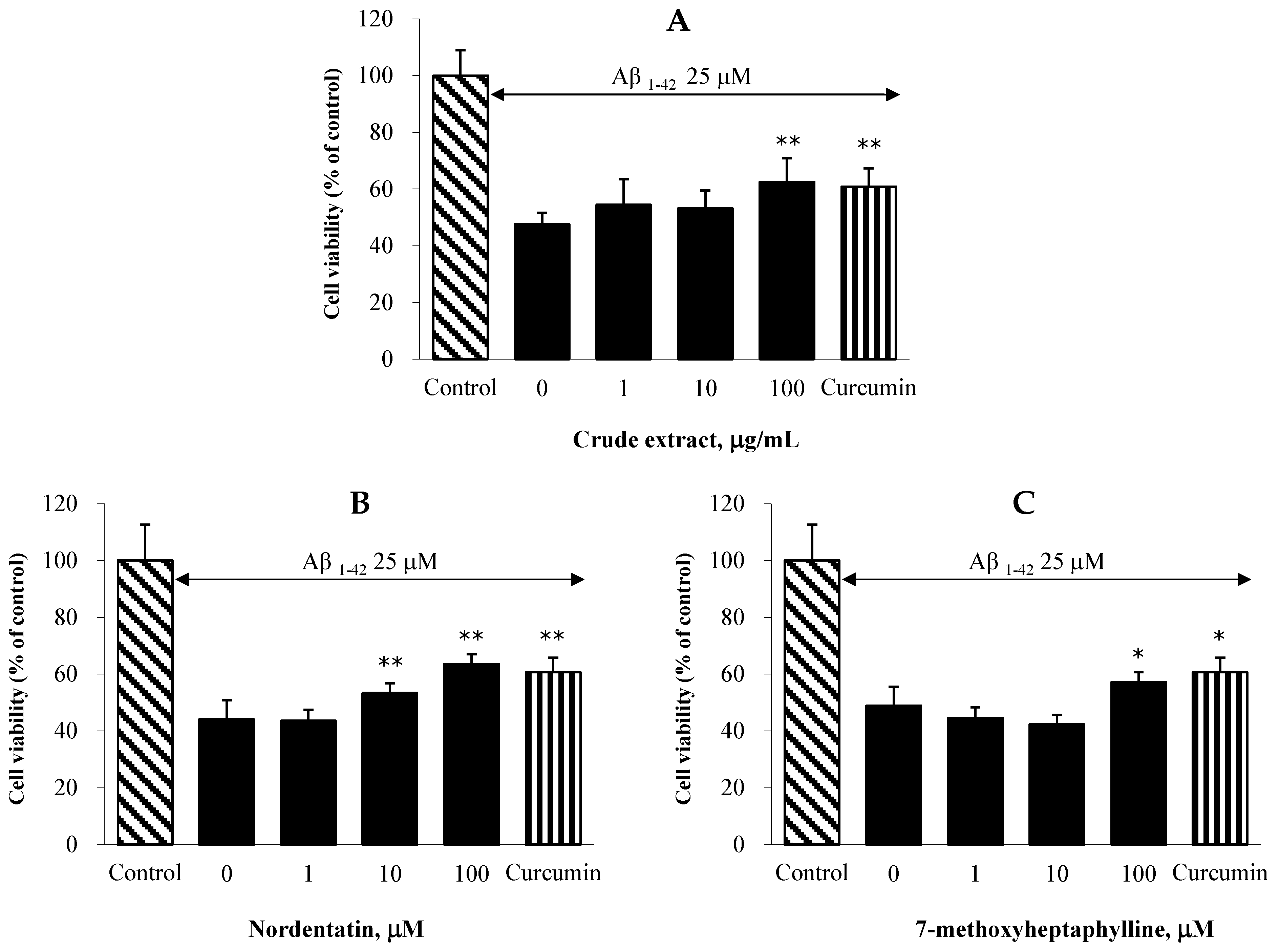
2.6. Effect on Aβ-Induced Neuronal Cell Death in C6 Cells
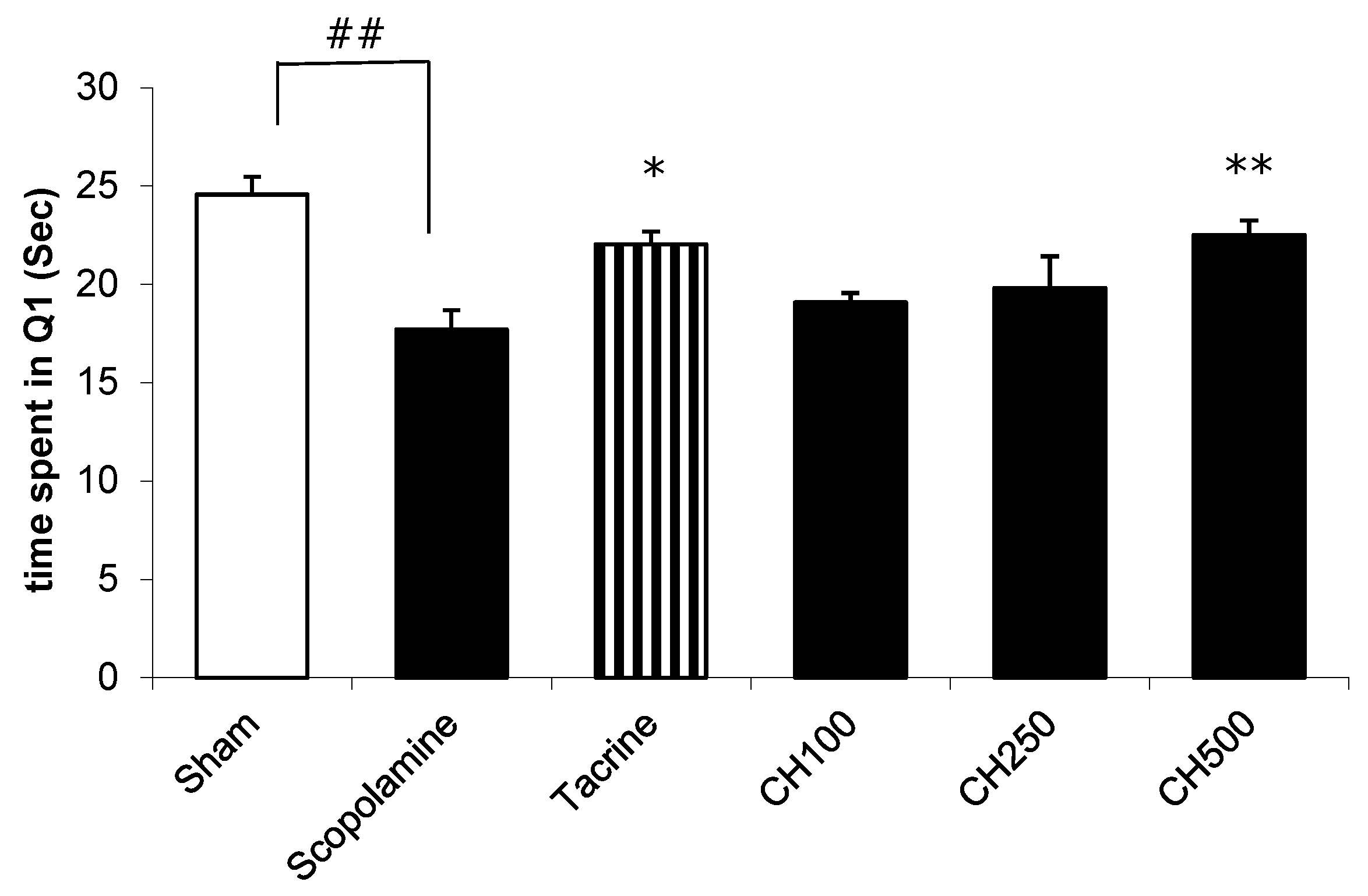
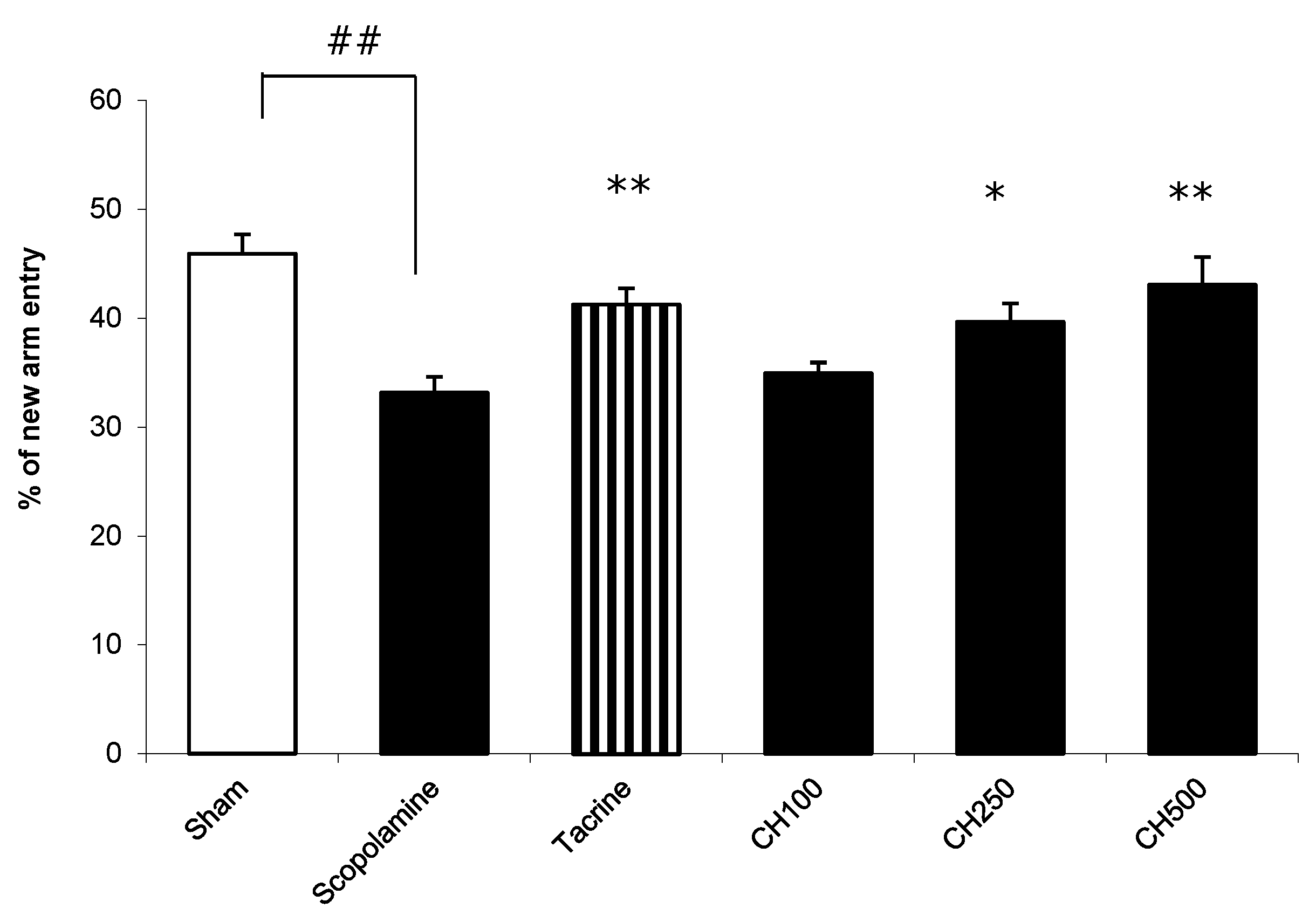
2.7. The Effect of the CH Extract on the Improvement of a Memory Deficit in Mice Induced by Scopolamine
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Plant Materials and Preparation of the Crude Extract
5.3. HPLC Analysis
5.4. In Vitro Antioxidant Activity Assays [7]
5.5. In Vitro Assay of AChE-Inhibitory Activity [45]
5.6. In Vitro Assay for AB-Aggregation Inhibition Using a Thioflavin-T Assay [46]
5.7. Effect on H2O2-Induced Oxidative Cell Damage in NG108-15 Cells [40]
5.8. Effect on β-Amyloid-Induced Cell Damage in C6 Cells [40]
5.9. The Effect of the CH Extract on the Memory Deficit in Mice Induced by Scopolamine
5.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nichols, E.; Steinmetz, J.D.; Vollset, S.E.; Fukutaki, K.; Chalek, J.; Abd-Allah, F.; Abdoli, A.; Abualhasan, A.; Abu-Gharbieh, E.; Akram, T.T.; et al. Estimation of the global prevalence of dementia in 2019 and forecasted prevalence in 2050: An analysis for the Global Burden of Disease Study 2019. Lancet Public Health 2022, 7, e105–e125. [Google Scholar] [CrossRef]
- Hampel, H.; Shen, Y.; Walsh, D.M.; Aisen, P.; Shaw, L.M.; Zetterberg, H.; Trojanowski, J.Q.; Blennow, K. Biological markers of amyloid β-related mechanisms in Alzheimer’s disease. Exp. Neurol. 2010, 223, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.G.; Francis, P.T.; Schwam, E.; Payne-Parrish, J. Cholinesterase inhibitors used in the treatment of Alzheimer’s disease: The relationship between pharmacological effects and clinical efficacy. Drugs Aging 2004, 21, 453–478. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Conde, S.; Rodriguez-Franco, M.I.; Martınez, A. Noncholinergic pharmacotherapy approaches to the future treatment of Alzheimer’s disease. Mini Rev. Med. Chem. 2002, 2, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Rosini, M.; Simoni, E.; Bartolini, M.; Cavalli, A.; Ceccarini, L.; Pascu, N. Inhibition of acetylcholinesterase, β-Amyloid aggregation, and NMDA receptors in Alzheimer’s disease: A promising direction for the multi-target-directed ligands gold rush. J. Med. Chem. 2008, 51, 4381–4384. [Google Scholar] [CrossRef] [PubMed]
- Kozurkova, M.; Hamulakova, S.; Gazova, Z.; Paulikova, H.; Kristian, P. Neuroactive multifunctional tacrine congeners with cholinesterase, anti-amyloid aggregation and neuroprotective properties. Pharmaceuticals 2011, 4, 382–418. [Google Scholar] [CrossRef]
- Thiratmatrakul, S.; Yenjai, C.; Waiwut, P.; Vajragupta, O.; Reubroycharoen, P.; Tohda, M.; Boonyarat, C. Synthesis, biological evaluation and molecular modeling study of novel tacrine-carbazole hybrids as potential multifunctional agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2014, 75, 21–30. [Google Scholar] [CrossRef]
- Sterling, J.; Herzig, Y.; Goren, T.; Finkelstein, N.; Lerner, D.; Goldenberg, W.; Miskolczi, I.; Molnar, S.; Rantal, F.; Tamas, T.; et al. Novel dual inhibitors of AChE and MAO derived from hydroxy aminoindan and phenylethylamine as potential tretament for Alzheimer’s disease. J. Med. Chem. 2002, 45, 5260–5279. [Google Scholar] [CrossRef]
- Marco-Contelles, J.; Leon, R.; De Los Rios, C.; Guglietta, A.; Terencio, J.; Lopez, M.G.; García, A.G.; Villarroya, M. Novel multipotent tacrine-dihydropyridine hybrids with Improved acetylcholinesterase inhibitory and neuroprotective activities as potential drugs for the treatment of Alzheimer’s disease. J. Med. Chem. 2006, 49, 7607–7610. [Google Scholar] [CrossRef]
- Vajragupta, O.; Boonyarat, C.; Olson, A.J.; Murakami, Y.; Tohda, M.; Matsumoto, K.; Watanabe, H. A novel neuroprotective agent with antioxidant and nitric oxide synthase inhibitory action. Free Radical Research. Free Radic. Res. 2006, 40, 677–687. [Google Scholar] [CrossRef]
- Hage, S.; Kienlen-Campard, P.; Octave, J.; Quetin-Leclercq, J. In vitro screening on β amyloid peptide production of plants used in traditional medicine for cognitive disorders. J. Ethnopharmacol. 2010, 131, 585–591. [Google Scholar] [CrossRef]
- Bastianetto, S.; Ramassamy, C.; Dore, S.; Christen, Y.; Poirier, J.; Quirion, R. The Gingko biloba extract protects hippocampal neurones against cell death induced by beta-amyloid. J. Ethnopharmacol. 2000, 12, 1882–1890. [Google Scholar]
- De Feudis, F.V.; Drieu, K. Ginkgo biloba extract (EGb 761) and CNS functions: Basic studies and clinical applications. Curr. Drug Targets 2000, 1, 25–58. [Google Scholar] [CrossRef]
- Yao, Z.; Drieu, K.; Papadopoulos, V. The Ginkgo biloba extract EGb 761 rescues the PC12 neuronal cells from beta-amyloid-induced cell death by inhibiting the formation of beta-amyloid-derived diffusible neurotoxic ligands. Brain Res. 2001, 889, 181–190. [Google Scholar] [CrossRef]
- Howes, M.R.; Houghton, P.J. Plants used in Chinese and Indian traditional medicine for improvement of memory and cognitive function. Pharmacol. Biochem. Behav. 2003, 75, 513–527. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, Z.; Butko, P.; Christen, Y.; Lambert, P.; Klein, W.L.; Link, C.D.; Luo, Y. Amyloid-beta-induced pathological behaviors are suppressed by Ginkgo biloba extract EGb 761 and Ginkgolides in transgenic Caenorhabditis elegans. J. Neurosci. 2006, 26, 13102–13113. [Google Scholar] [CrossRef]
- Ismail, A.A.; Ahmad, B.A.; Mohamed, A.; Siddig, I.A.; Mohamed, Y.I. A review of traditional uses, phytochemical and pharmacological aspects of selected members of Clausena genus (Rutaceae). J. Med. Plants Res. 2012, 6, 5107–5118. [Google Scholar] [CrossRef]
- Thongthoom, T.; Songsiang, U.; Phaosiri, C.; Yenjai, C. Biological activity of chemical constituents from Clausena harmandiana. Arch. Pharm. Res. 2010, 33, 675–680. [Google Scholar] [CrossRef]
- Songsiang, U.; Thongthoom, T.; Zeekpudsa, P.; Kukongviriyapan, V.; Boonyarat, C.; Wangboonskul, J.; Yenjai, C. Antioxidant activity and cytotoxicity against cholangiocarcinoma of carbazoles and coumarins from Clausena harmandiana. ScienceAsia 2012, 38, 75–81. [Google Scholar] [CrossRef]
- Wangboonskul, J.; Prawan, A.; Takthaisong, P.; Sasithornwetchakun, W.; Boonyarat, C.; Yenjai, C.; Mahakunakorn, P. Analgesic, anti-inflammatory, antipyretic activities and acute toxicity of the ethanolic extract of Clausena harmandiana Pierre in animals. J. Asian Assoc. Sch. Pharm. 2012, 1, 159–169. [Google Scholar]
- Wangboonskul, J.; Tunsirikongkon, A.; Sasithornwetchakun, W. Simultaneous determination of nine analytes in Clausena harmandiana Pierre. by new developed high-performance liquid chromatography method and the influence of locations in Thailand on level of nordentatin and dentatin. Pharmacogn. Mag. 2015, 11, 1. [Google Scholar] [CrossRef]
- Yenjai, C.; Sripontan, S.; Sriprajun, P.; Kittakoop, P.; Jintasirikul, A.; Tanticharoen, M.; Thebtaranonth, Y. Coumarins and carbazoles with antiplasmodial activity from Clausena harmandiana. Planta Med. 2000, 66, 277–279. [Google Scholar] [CrossRef]
- Noipha, K.; Thongthoom, T.; Songsiang, U.; Boonyarat, C.; Yenjai, C. Carbazoles and coumarins from Clausena harmandiana stimulate glucose uptake in L6 myotubes. Diabetes Res. Clin. Pract. 2010, 90, e67–e71. [Google Scholar] [CrossRef]
- Boonyarat, C.; Boonput, P.; Tongloh, N.; Rawiwun, K.; Chaiwiwatrakul, S.; Yenjai, C.; Waiwut, P. Nordentatin inhibits neuroblastoma cell proliferation and migration through regulation of GSK-3 Pathway. Curr. Issues Mol. Biol. 2022, 44, 1062–1074. [Google Scholar] [CrossRef]
- Boonyarat, C.; Yenjai, C.; Vajragupta, O.; Waiwut, P. Heptaphylline induces apoptosis in human colon adenocarcinoma cells through Bid and Akt/NF-×B (p65) Pathways. Asian Pac. J. Cancer Prev. 2015, 15, 10483–10487. [Google Scholar] [CrossRef]
- Soto-Ortega, D.D.; Murphy, B.P.; Gonzalez-Velasquez, F.J.; Wilson, K.A.; Xie, F.; Wang, Q.; Moss, M.A. Inhibition of amyloid-β aggregation by coumarin analogs can be manipulated by functionalization of the aromatic center. Bioorg. Med. Chem. 2011, 19, 2596–2602. [Google Scholar] [CrossRef]
- Anand, P.; Singh, B.; Singh, N. A review on coumarins as acetylcholinesterase inhibitors for Alzheimer’s disease. Bioorg. Med. Chem. 2012, 20, 1175–1180. [Google Scholar] [CrossRef]
- Nakamura, T.; Kodama, N.; Arai, Y.; Kumamoto, T.; Higuchi, Y.; Chaichantipyuth, C.; Ishikawa, T.; Ueno, K.; Yano, S. Inhibitory effect of oxycoumarins isolated from the Thai medicinal plant Clausena guillauminii on the inflammation mediators, iNOS, TNF-α, and COX-2 expression in mouse macrophage RAW 264.7. J. Nat. Med. 2009, 63, 21–27. [Google Scholar] [CrossRef]
- Wu, T.S.; Huang, S.C.; Wu, P.L.; Kuoh, C.S. Alkaloidal and other constituents from the root bark of Clausena excavata. Phytochemistry 1999, 52, 523–527. [Google Scholar] [CrossRef]
- Jantakoon, P.; Tadtong, S.; Puthongking, P. Neuritogenic and antioxidant activities of nordentatin from Clausena harmandiana. J. Asian Assoc. Sch. Pharm. 2012, 1, 180–186. [Google Scholar]
- Murray, T.K.; Cross, A.J.; Green, A.R. Reversal by tetrahydroaminoacridine of scopolamine-induced memory and performance deficits in rats. Psychopharmacology 1991, 105, 134–136. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Reed, T.; Newman, S.F.; Sultana, R. Roles of amyloid beta-peptideassociatedoxidative stress and brain protein modifications in the pathogenesis of Alzheimer’s disease and mild cognitive impairment. Free Radic. Biol. Med. 2007, 43, 658–677. [Google Scholar] [CrossRef] [PubMed]
- Yatin, S.M.; Varadarajan, S.; Link, C.D.; Butterfield, D.A. In vitro and in vivo oxidative stress associated with Alzheimer’s amyloid beta-peptide (1–42). Neurobiol. Aging 1999, 20, 325–330. [Google Scholar] [PubMed]
- Lee, V.M. Amyloid binding ligands as Alzheimer’s disease therapies. Neurobiol. Aging 2002, 23, 1039–1042. [Google Scholar] [CrossRef]
- Nie, Q.; Du, X.G.; Geng, M.Y. Small molecule inhibitors of amyloid β peptide aggregation as a potential therapeutic strategy for Alzheimer’s disease. Acta Pharmacol. Sin. 2011, 32, 545–551. [Google Scholar] [CrossRef]
- Kolstoe, S.E.; Wood, S.P. Drug targets for amyloidosis. Biochem. Soc. Trans. 2010, 38, 466–470. [Google Scholar] [CrossRef]
- Ebert, U.; Kirch, W. Scopolamine model of dementia: Electroencephalogram findings and cognitive performance. Eur. J. Clin. Investig. 1998, 28, 944–949. [Google Scholar] [CrossRef]
- Morris, R.G. Development of a water maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Takomthong, P.; Waiwut, P.; Yenjai, C.; Sombatsri, A.; Reubroycharoen, P.; Lei, L.; Lai, R.; Chaiwiwatrakul, S.; Boonyarat, C. Multi-target actions of acridones from Atalantia monophylla towards Alzheimer’s pathogenesis and their pharmacokinetic properties. Pharmaceuticals 2021, 14, 888. [Google Scholar] [CrossRef]
- Takomthong, P.; Waiwut, P.; Yenjai, C.; Sripanidkulchai, B.; Reubroycharoen, P.; Lai, R.; Kamau, P.; Boonyarat, C. Structure-activity analysis and molecular docking studies of coumarins from Toddalia asiatica as multifunctional agents for Alzheimer’s disease. Biomedicines 2020, 8, 107. [Google Scholar] [CrossRef]
- El-Sherbiny, D.A.; Khalifa, A.E.; Attia, A.S.; Eldenshary, E.E. Hypericum perforatum extract demonstrates antioxidant properties against elevated rat brain oxidative status induced by amnestic dose of scopolamine. Pharmacol. Biochem. Behav. 2003, 76, 525–533. [Google Scholar] [CrossRef]
- Saxena, S.; Thornicroft, G.; Knapp, M.; Whiteford, H. Resources for mental health: Scarcity, inequity, and inefficiency. Lancet 2007, 370, 878–889. [Google Scholar] [CrossRef]
- Sharma, N.; Garg, V.; Arpita, P. Antihyperglycemic, antihyperlipidemic and antioxidative potential of Prosopis cineraria bark. Indian J. Clin. Biochem. 2010, 25, 193–200. [Google Scholar] [CrossRef]
- Shin, C.Y.; Kim, H.S.; Cha, K.; Won, D.H.; Lee, J.; Jang, S.W.; Sohn, U. The effects of donepezil, an acetylcholinesterase inhibitor, on impaired learning and memory in rodents. Biomol. Ther. 2018, 26, 274–281. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Chheng, C.; Waiwut, P.; Plekratoke, K.; Chulikhit, Y.; Daodee, S.; Monthakantirat, O.; Pitiporn, S.; Musigavong, N.; Kwankhao, P.; Boonyarat, C. Multitarget activities of Kleeb Bua Daeng, a Thai traditional herbal formula, against Alzheimer’s disease. Pharmaceuticals 2020, 13, 79. [Google Scholar] [CrossRef]
- Arunrungvichian, K.; Boonyarat, C.; Fokin, V.V.; Taylor, P.; Vajragupta, O. Cognitive improvements in a mouse model with substituted 1,2,3-triazole agonists for nicotinic acetylcholine receptors. ACS Chem. Neurosci. 2015, 6, 1331–1340. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Constituents in CH Root Extract | Structure | Retention Time (min) |
|---|---|---|
| xanthoxyletin |  | 28.60 |
| nordentatin |  | 33.73 |
| 7-methoxyheptaphylline |  | 39.92 |
| heptaphylline |  | 40.67 |
| dentatin |  | 44.66 |
| Compounds | ABTS Assay a (IC50) | AChE Assay b (IC50) | Aβ-Aggregation Assay c (% Inhibition) |
|---|---|---|---|
| Tacrine | nd | 0.36 ± 0.16 | nd |
| Trolox (µM) | 23.67 ± 1.41 | nd | nd |
| Curcumin (10 µM) | nd | nd | 44.86 ± 6.90 |
| CH extract (µg/mL) | 42.21 ± 2.11 | 86.71 ± 5.23 | 65.28 ± 7.54 |
| 7-methoxyheptaphyline (µM) | 10.78 ± 0.11 | >100 µM | 18.01 ± 5.72 |
| Nordentatin (µM) | 3.78 ± 0.13 | 67.79 ± 4.19 | 75.34 ± 4.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonyarat, C.; Yenjai, C.; Monthakantirat, O.; Kaewamatawong, R.; Poonsawas, P.; Wangboonskul, J.; Chaiwiwatrakul, S.; Waiwut, P. Multifunctionality of Clausena harmandiana Extract and Its Active Constituents against Alzheimer’s Disease. Curr. Issues Mol. Biol. 2022, 44, 3681-3694. https://doi.org/10.3390/cimb44080252
Boonyarat C, Yenjai C, Monthakantirat O, Kaewamatawong R, Poonsawas P, Wangboonskul J, Chaiwiwatrakul S, Waiwut P. Multifunctionality of Clausena harmandiana Extract and Its Active Constituents against Alzheimer’s Disease. Current Issues in Molecular Biology. 2022; 44(8):3681-3694. https://doi.org/10.3390/cimb44080252
Chicago/Turabian StyleBoonyarat, Chantana, Chavi Yenjai, Orawan Monthakantirat, Rawiwun Kaewamatawong, Pattaporn Poonsawas, Jinda Wangboonskul, Suchada Chaiwiwatrakul, and Pornthip Waiwut. 2022. "Multifunctionality of Clausena harmandiana Extract and Its Active Constituents against Alzheimer’s Disease" Current Issues in Molecular Biology 44, no. 8: 3681-3694. https://doi.org/10.3390/cimb44080252