
Effects of Dietary Inclusion of β-Hydroxy-β-Methylbutyrate on Growth Performance, Fat Deposition, Bile Acid Metabolism, and Gut Microbiota Function in High-Fat and High-Cholesterol Diet-Challenged Layer Chickens

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Measurement of Growth Performance
2.3. Slaughter Indicators and Organ Index
2.4. Measurement of Plasma and Hepatic Parameters
2.5. RNA Extraction and Quantitative Real-Time PCR
2.6. Hematoxylin and Eosin (H&E) Staining
2.7. Gut Microbiome Analysis by 16S rRNA Sequencing
2.8. Statistical Analysis
3. Results
3.1. Effects of Dietary HMB on the Growth Performance of HFHCD−Challenged Layer Chickens
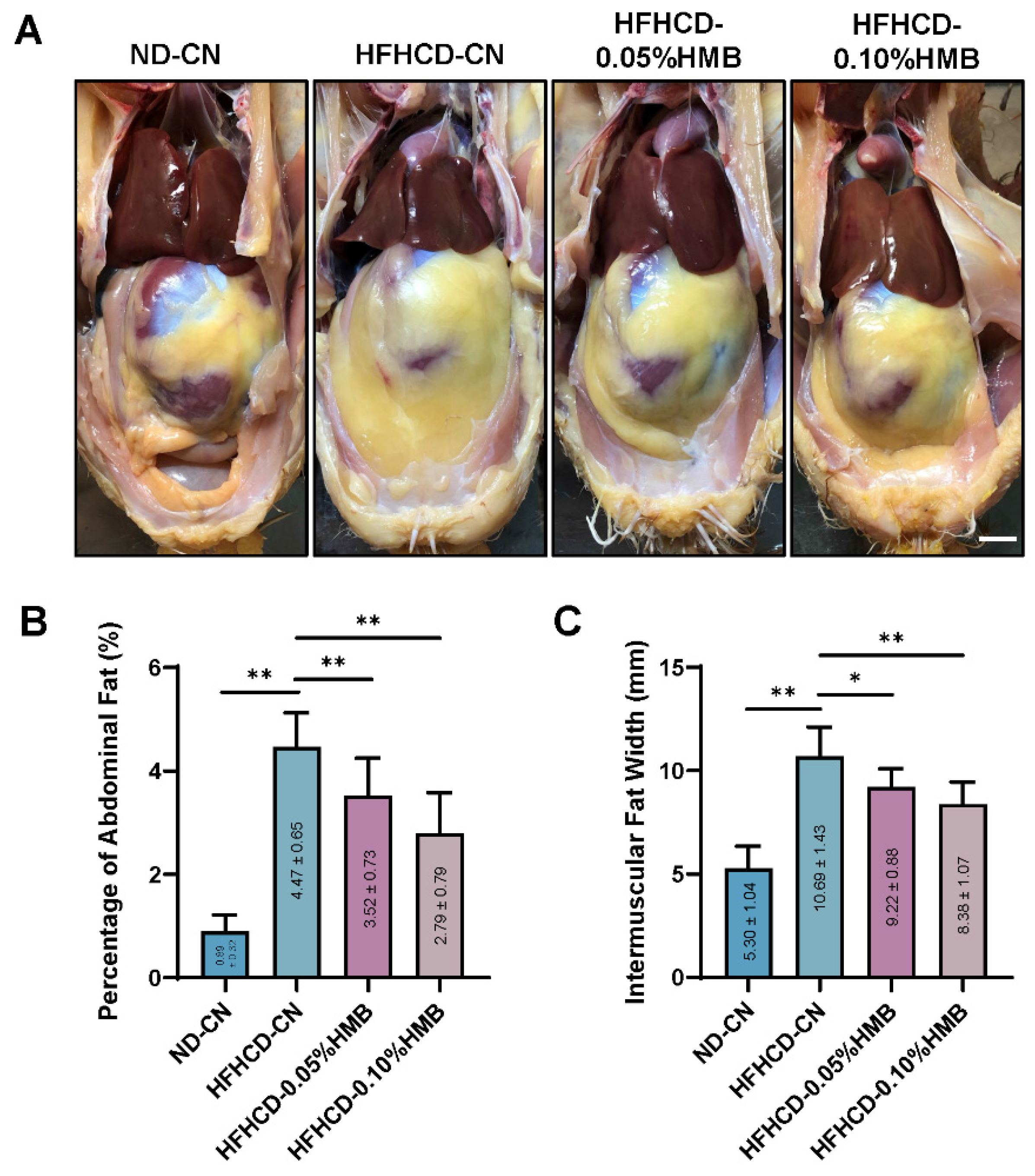
3.2. Effects of Dietary HMB on HFHCD−Induced Lipid Accumulation in Layer Chickens
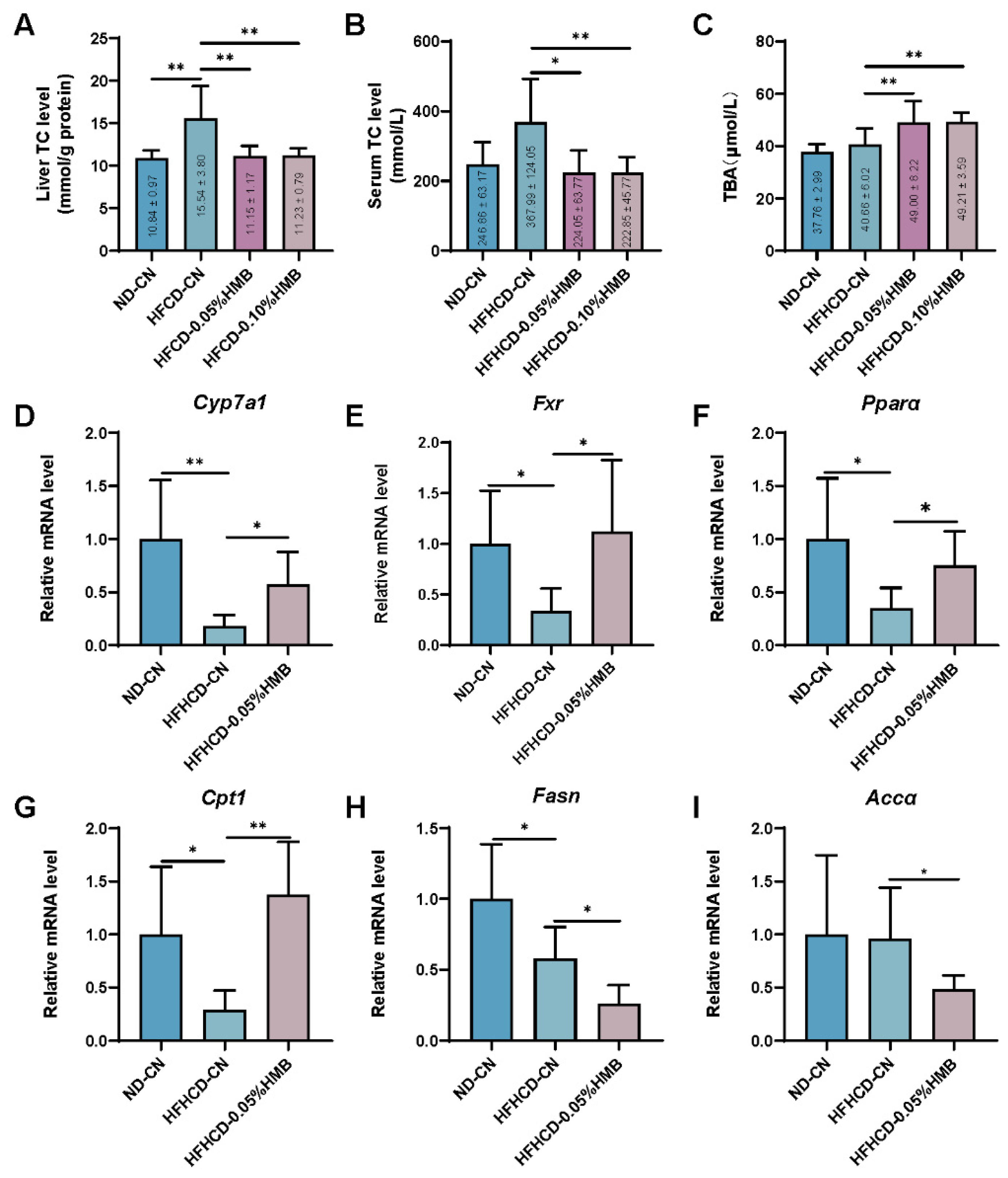
3.3. Effects of Dietary HMB on Hepatic Bile Acid Metabolism in HFHCD−Challenged Layer Chickens
3.4. Effects of Dietary HMB on Gut Microbial Diversity in HFHCD−Challenged Layer Chickens
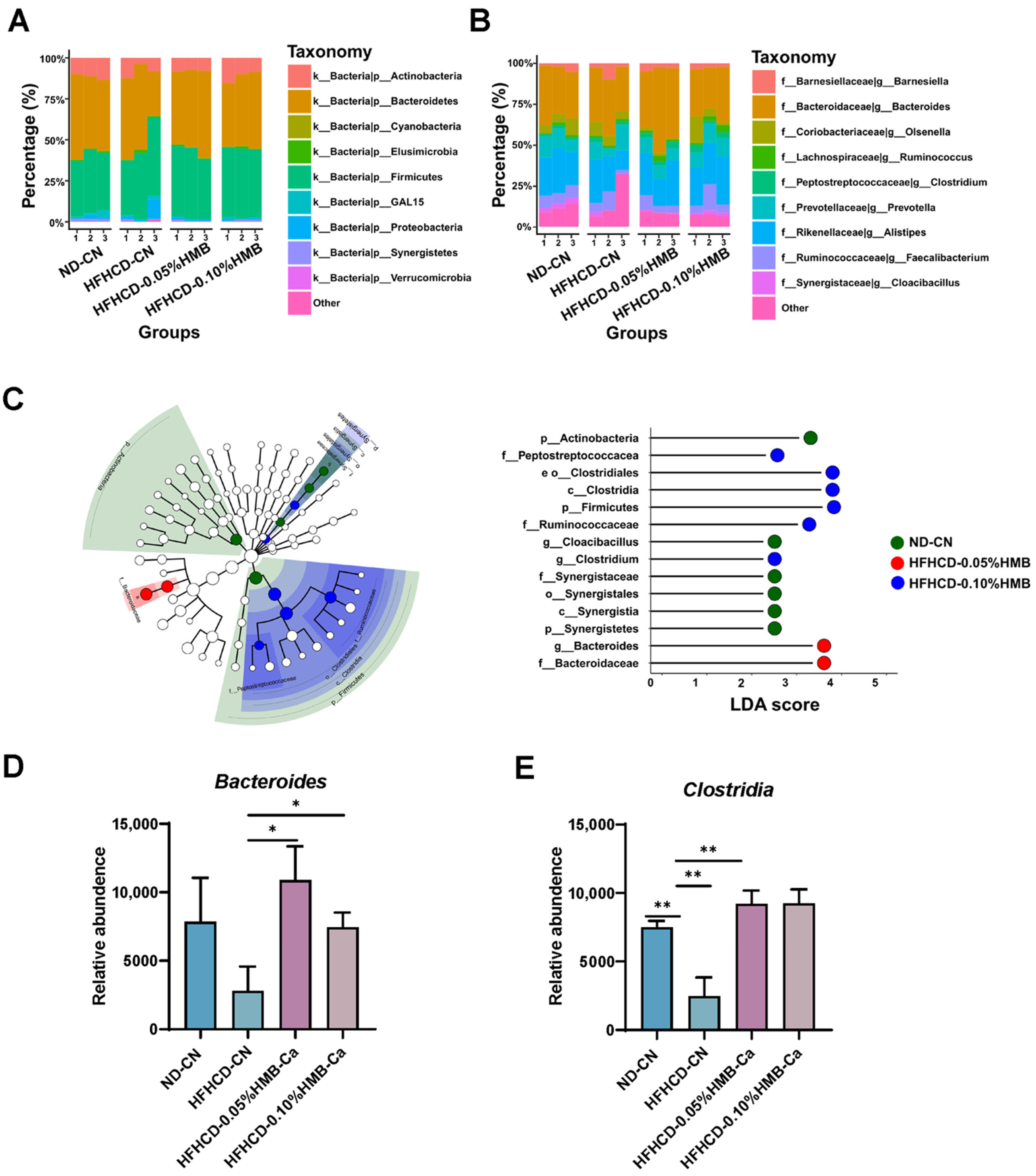
3.5. Effects of Dietary HMB on Gut Microbial Composition in HFHCD–Challenged Layer Chickens
3.6. Effects of HMB on Gut Microbial Function in HFHCD–Challenged Layer Chickens
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Accα | Acetyl-coa carboxylase alpha |
| ADFI | Average daily feed intake |
| ADG | Average daily gain |
| ASV | Amplicon sequence variant |
| Cpt1 | Carnitine palmitoyltransferase 1 |
| Cyp7a1 | Cytochrome P450 family 7 subfamily A member 1 |
| DNL | De novo lipogenesis |
| F/G | Feed-to-gain ratio |
| Fasn | Fatty acid synthase |
| FLHS | Fatty liver haemorrhagic syndrome |
| Fxr | Farnesoid X-activated receptor |
| HFD | High-fat diet |
| HFHCD | High-fat and high-cholesterol diet |
| HMB | Β-hydroxy-β-methylbutyrate |
| LDA | Linear discriminant analysis |
| LEfSe | Linear discriminant analysis (LDA) effect size |
| NAFLD | Non-alchoholic fatty liver disease |
| PAF | Percentage of abdominal fat |
| Ppara | Peroxisome proliferator-activated receptor alpha |
| SCFAs | Short-chain fatty acids |
| TBA | Total bile acid |
| TC | Total cholesterol |
| TG | Triglycerides |
| VLDL | Very low-density lipoprotein |
| Vg | Vitellogenin |
References
- Groves, P.J. Non-infectious diseases and laying hen welfare. Anim. Prod. Sci. 2021, 61, 1013–1017. [Google Scholar] [CrossRef]
- Shini, A.; Shini, S.; Bryden, W.L. Fatty liver haemorrhagic syndrome occurrence in laying hens: Impact of production system. Avian Pathol. 2019, 48, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Xing, C.; Cao, H.; Zhang, C.; Luo, J.; Guo, X.; Hu, G. Insulin resistance and metabonomics analysis of fatty liver haemorrhagic syndrome in laying hens induced by a high-energy low-protein diet. Sci. Rep. 2019, 9, 10141. [Google Scholar] [CrossRef] [PubMed]
- Walzem, R.L.; Chen, S.-E. Obesity-induced dysfunctions in female reproduction: Lessons from birds and mammals. Adv. Nutr. 2014, 5, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozenboim, I.; Mahato, J.; Cohen, N.; Tirosh, O. Low protein and high-energy diet: A possible natural cause of fatty liver hemorrhagic syndrome in caged White Leghorn laying hens. Poult. Sci. 2016, 95, 612–621. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [Green Version]
- Fouad, A.M.; El-Senousey, H.K. Nutritional factors affecting abdominal fat deposition in poultry: A review. Asian-Australas. J. Anim. Sci. 2014, 27, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Kim, W.K.; Cline, M.A.; Gilbert, E.R. Factors affecting adipose tissue development in chickens: A review. Poult. Sci. 2017, 96, 3687–3699. [Google Scholar] [CrossRef]
- Desert, C.; Baeza, E.; Aite, M.; Boutin, M.; Le Cam, A.; Montfort, J.; Houee-Bigot, M.; Blum, Y.; Roux, P.F.; Hennequet-Antier, C.; et al. Multi-tissue transcriptomic study reveals the main role of liver in the chicken adaptive response to a switch in dietary energy source through the transcriptional regulation of lipogenesis. BMC Genom. 2018, 19, 187. [Google Scholar] [CrossRef]
- Sato, K.; Kamada, T. Regulation of bile acid, cholesterol, and fatty acid synthesis in chicken primary hepatocytes by different concentrations of T0901317, an agonist of liver X receptors. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2011, 158, 201–206. [Google Scholar] [CrossRef]
- Schneider, W.J. Lipid transport to avian oocytes and to the developing embryo. J. Biomed. Res. 2016, 30, 174–180. [Google Scholar] [PubMed]
- Chen, S.; Luo, S.; Yan, C. Gut Microbiota Implications for Health and Welfare in Farm Animals: A Review. Animals 2021, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Boscaini, S.; Leigh, S.J.; Lavelle, A.; Garcia-Cabrerizo, R.; Lipuma, T.; Clarke, G.; Schellekens, H.; Cryan, J.F. Microbiota and body weight control: Weight watchers within? Mol. Metab. 2022, 57, 101427. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Yacyshyn, B.R.; Yacyshyn, M.B. Gut microbiota and obesity: An opportunity to alter obesity through faecal microbiota transplant (FMT). Diabetes Obes. Metab. 2019, 21, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, F.; Li, Y.; Tang, Y.; Kong, X.; Feng, Z.; Anthony, T.G.; Watford, M.; Hou, Y.; Wu, G.; et al. The role of leucine and its metabolites in protein and energy metabolism. Amino Acids 2016, 48, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Holecek, M. Beta-hydroxy-beta-methylbutyrate supplementation and skeletal muscle in healthy and muscle-wasting conditions. J. Cachexia Sarcopenia Muscle 2017, 8, 529–541. [Google Scholar] [CrossRef]
- Zheng, C.; Song, B.; Duan, Y.; Zhong, Y.; Yan, Z.; Zhang, S.; Li, F. Dietary beta-hydroxy-beta-methylbutyrate improves intestinal function in weaned piglets after lipopolysaccharide challenge. Nutrition 2020, 78, 110839. [Google Scholar] [CrossRef]
- Duan, Y.; Zheng, C.; Zhong, Y.; Song, B.; Yan, Z.; Kong, X.; Deng, J.; Li, F.; Yin, Y. Beta-hydroxy beta-methyl butyrate decreases muscle protein degradation via increased Akt/FoxO3a signaling and mitochondrial biogenesis in weanling piglets after lipopolysaccharide challenge. Food Funct. 2019, 10, 5152–5165. [Google Scholar] [CrossRef]
- Zheng, J.; Zheng, C.; Song, B.; Guo, Q.; Zhong, Y.; Zhang, S.; Zhang, L.; Duan, G.; Li, F.; Duan, Y. HMB Improves Lipid Metabolism of Bama Xiang Mini-Pigs via Modulating the Bacteroidetes-Acetic Acid-AMPKα Axis. Front. Microbiol. 2021, 12, 736997. [Google Scholar] [CrossRef]
- Duan, Y.; Zhong, Y.; Xiao, H.; Zheng, C.; Song, B.; Wang, W.; Guo, Q.; Li, Y.; Han, H.; Gao, J. Gut microbiota mediates the protective effects of dietary β-hydroxy-β-methylbutyrate (HMB) against obesity induced by high-fat diets. FASEB J. 2019, 33, 10019–10033. [Google Scholar] [CrossRef]
- Tang, Z.; Song, B.; Zheng, C.; Zheng, J.; Yin, Y.; Chen, J. Dietary beta-hydroxy-beta-methyl butyrate supplementation affects growth, carcass characteristics, meat quality, and serum metabolomics profile in broiler chickens. Front. Physiol. 2021, 12, 633964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Tang, Z.; Zheng, C.; Zhong, Y.; Zheng, J.; Duan, G.; Yin, Y.; Duan, Y.; Song, Z. Dietary Beta-Hydroxy-Beta-Methyl Butyrate Supplementation Inhibits Hepatic Fat Deposition via Regulating Gut Microbiota in Broiler Chickens. Microorganisms 2022, 10, 169. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wu, H.; Shi, L.; Zhang, X.; Sheng, R.; Yin, F.; Gooneratne, R. Study of Bacillus subtilis on growth performance, nutrition metabolism and intestinal microflora of 1 to 42 d broiler chickens. Anim. Nutr. 2017, 3, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wu, T.; Fu, Q.; Liao, Q.; Li, Y.; Huang, T.; Li, Y.; Zhou, L.; Song, Z. Maternal High-Fructose Intake Activates Myogenic Program in Fetal Brown Fat and Predisposes Offspring to Diet-Induced Metabolic Dysfunctions in Adulthood. Front. Nutr. 2022, 9, 848983. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hong, Y.; Liao, Y.; Tian, K.; Sun, H.; Liu, X.; Tang, Y.; Hassanin, A.A.; Abdelnour, S.A.; Suthikrai, W. Dietary Lasia spinosa Thw. Improves Growth Performance in Broilers. Front. Nutr. 2021, 8, 775223. [Google Scholar] [CrossRef]
- Mashek, D.G. Hepatic fatty acid trafficking: Multiple forks in the road. Adv. Nutr. 2013, 4, 697–710. [Google Scholar] [CrossRef] [Green Version]
- Jump, D.B.; Botolin, D.; Wang, Y.; Xu, J.; Christian, B.; Demeure, O. Fatty acid regulation of hepatic gene transcription. Nutr. J. 2005, 135, 2503–2506. [Google Scholar] [CrossRef]
- Russell, D.W. The enzymes, regulation, and genetics of bile acid synthesis. Annu. Rev. Biochem. 2003, 72, 137–174. [Google Scholar] [CrossRef] [Green Version]
- Ge, M.X.; Shao, R.G.; He, H.W. Advances in understanding the regulatory mechanism of cholesterol 7alpha-hydroxylase. Biochem. Pharmacol. 2019, 164, 152–164. [Google Scholar] [CrossRef]
- Stayrook, K.R.; Bramlett, K.S.; Savkur, R.S.; Ficorilli, J.; Cook, T.; Christe, M.E.; Michael, L.F.; Burris, T.P. Regulation of carbohydrate metabolism by the farnesoid X receptor. Endocrinology 2005, 146, 984–991. [Google Scholar] [CrossRef]
- Schmitt, J.; Kong, B.; Stieger, B.; Tschopp, O.; Schultze, S.M.; Rau, M.; Weber, A.; Müllhaupt, B.; Guo, G.L.; Geier, A. Protective effects of farnesoid X receptor (FXR) on hepatic lipid accumulation are mediated by hepatic FXR and independent of intestinal FGF15 signal. Liver Int. 2015, 35, 1133–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massafra, V.; van Mil, S.W. Farnesoid X receptor: A “homeostat” for hepatic nutrient metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.; Ferrell, J.M. Discovery of farnesoid X receptor and its role in bile acid metabolism. Mol. Cell. Endocrinol. 2022, 548, 111618. [Google Scholar] [CrossRef]
- Xi, Y.; Li, H. Role of farnesoid X receptor in hepatic steatosis in nonalcoholic fatty liver disease. Biomed. Pharmacother. 2020, 121, 109609. [Google Scholar] [CrossRef]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A. Gut microbiota and obesity: A role for probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [Green Version]
- Gauffin Cano, P.; Santacruz, A.; Moya, Á.; Sanz, Y. Bacteroides uniformis CECT 7771 ameliorates metabolic and immunological dysfunction in mice with high-fat-diet induced obesity. PLoS ONE 2012, 7, e41079. [Google Scholar]
- Xiang, H.; Gan, J.; Zeng, D.; Li, J.; Yu, H.; Zhao, H.; Yang, Y.; Tan, S.; Li, G.; Luo, C. Specific microbial taxa and functional capacity contribute to chicken abdominal fat deposition. Front. Microbiol. 2021, 12, 643025. [Google Scholar] [CrossRef]
- Hou, Q.; Kwok, L.-Y.; Zheng, Y.; Wang, L.; Guo, Z.; Zhang, J.; Huang, W.; Wang, Y.; Leng, L.; Li, H. Differential fecal microbiota are retained in broiler chicken lines divergently selected for fatness traits. Sci. Rep. 2016, 6, 37376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, C.C.; Monteil, M.A.; Davis, E.M. Overweight and Obesity in Children Are Associated with an Abundance of Firmicutes and Reduction of Bifidobacterium in Their Gastrointestinal Microbiota. Child Obes. 2020, 16, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.; Bell, R.; Klag, K.A.; Lee, S.-H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S. T cell–mediated regulation of the microbiota protects against obesity. Science 2019, 365, eaat9351. [Google Scholar] [CrossRef]
- Astbury, S.; Atallah, E.; Vijay, A.; Aithal, G.P.; Grove, J.I.; Valdes, A.M. Lower gut microbiome diversity and higher abundance of proinflammatory genus Collinsella are associated with biopsy-proven nonalcoholic steatohepatitis. Gut Microbes 2020, 11, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Buzała, M.; Janicki, B.; Czarnecki, R. Consequences of different growth rates in broiler breeder and layer hens on embryogenesis, metabolism and metabolic rate: A review. Poult. Sci. 2015, 94, 728–733. [Google Scholar] [CrossRef]
- Park, S.-Y.; Byeon, D.-S.; Kim, G.-W.; Kim, H.-Y. Carcass and retail meat cuts quality properties of broiler chicken meat based on the slaughter age. J. Anim. Sci. Technol. 2021, 63, 180. [Google Scholar] [CrossRef]
- Sandra, S. Life Cycle of a Laying Hen. Available online: https://livestock.extension.wisc.edu/articles/life-cycle-of-a-laying-hen/ (accessed on 10 May 2022).
- Polyzos, S.A.; Kountouras, J.; Mantzoros, C.S. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metabolism 2019, 92, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Trauner, M.; Claudel, T.; Fickert, P.; Moustafa, T.; Wagner, M. Bile acids as regulators of hepatic lipid and glucose metabolism. Dig. Dis. 2010, 28, 220–224. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content | |
|---|---|---|
| 1 to 4 Weeks | 4 to 10 Weeks | |
| Ingredients, % | ||
| Corn | 56.80 | 51.50 |
| Wheat bran | 5.00 | 10.00 |
| Soybean meal | 30.00 | 32.00 |
| Fish meal | 3.00 | - |
| Limestone | 1.40 | 1.50 |
| CaHPO4 | 1.20 | 1.00 |
| Lard oil | 0.60 | 2.00 |
| Premix 1 | 2.00 | 2.00 |
| Total | 100.00 | 100.00 |
| Nutrients (calculated value) | ||
| CP | 20.01 | 20.70 |
| EE | 3.44 | 4.42 |
| Ash | 5.26 | 5.12 |
| Ca | 1.27 | 1.28 |
| TP | 0.60 | 0.55 |
| AP | 0.73 | 0.78 |
| Lys | 1.10 | 1.03 |
| Met | 0.48 | 0.43 |
| Genes | Primer Sequences (5′→3′) | GeneBank Accession Number |
|---|---|---|
| Accα | F: AGTCCTGATTGAGCATGGCA | NM_205505.1 |
| R: CTCCAGATGGCGGTAGATTC | ||
| β-actin | F: TGCGTGACATCAAGGAGAAG | NM_205518.1 |
| R: TGCCAGGGTACATTGTGGTA | ||
| Cpt1 | F: GCCAAGTCGCTCGCTGATGAC | DQ314726.1 |
| R: ACGCCTCGTAGGTCAGACAGAAC | ||
| Cyp7a1 | F: CATTCTGTTGCCAGGTGATGTT | AY700578 |
| R: GCTCTCTCTGTTTCCCGCTTT | ||
| Fasn | F: TGAAGGACCTTATCGATTGC | NM_205155.4 |
| R: GCATGGGAAGCATTTTGTTGT | ||
| Fxr | F: AGTAGAAGCCATGTTCCTCCGTT | AF492497 |
| R: GCAGTGCATATTCCTCCTGTGTC | ||
| Ppara | F: GAATGCCACAAGCGGAGAAGGAG | NM_001001464.1 |
| R: GCTCGCAGATCAGCAGATTCAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Q.; Wu, T.; Fu, Q.; Wang, P.; Zhao, Y.; Li, Y.; Xiao, H.; Zhou, L.; Song, Z. Effects of Dietary Inclusion of β-Hydroxy-β-Methylbutyrate on Growth Performance, Fat Deposition, Bile Acid Metabolism, and Gut Microbiota Function in High-Fat and High-Cholesterol Diet-Challenged Layer Chickens. Curr. Issues Mol. Biol. 2022, 44, 3413-3427. https://doi.org/10.3390/cimb44080235
Liao Q, Wu T, Fu Q, Wang P, Zhao Y, Li Y, Xiao H, Zhou L, Song Z. Effects of Dietary Inclusion of β-Hydroxy-β-Methylbutyrate on Growth Performance, Fat Deposition, Bile Acid Metabolism, and Gut Microbiota Function in High-Fat and High-Cholesterol Diet-Challenged Layer Chickens. Current Issues in Molecular Biology. 2022; 44(8):3413-3427. https://doi.org/10.3390/cimb44080235
Chicago/Turabian StyleLiao, Qichao, Tian Wu, Qinghua Fu, Peng Wang, Yameng Zhao, Yan Li, Haihan Xiao, Lei Zhou, and Ziyi Song. 2022. "Effects of Dietary Inclusion of β-Hydroxy-β-Methylbutyrate on Growth Performance, Fat Deposition, Bile Acid Metabolism, and Gut Microbiota Function in High-Fat and High-Cholesterol Diet-Challenged Layer Chickens" Current Issues in Molecular Biology 44, no. 8: 3413-3427. https://doi.org/10.3390/cimb44080235