
Apigetrin Abrogates Lipopolysaccharide-Induced Inflammation in L6 Skeletal Muscle Cells through NF-κB/MAPK Signaling Pathways

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Cell Culture and Apigetrin Treatment
2.3. Cell Viability Assay
2.4. Western Blotting
2.5. Molecular Docking Analysis
2.6. Statistical Analysis
3. Results
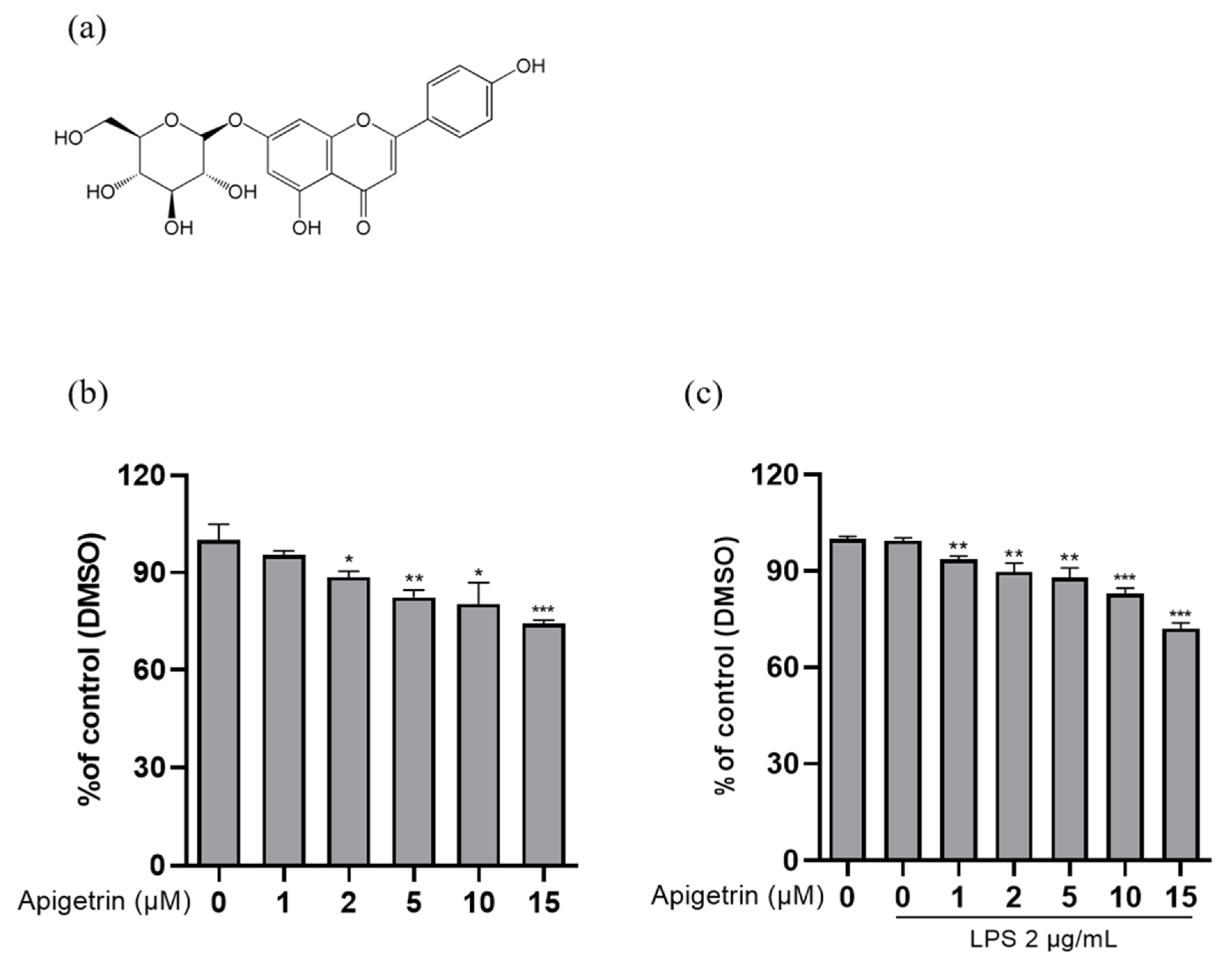
3.1. Effects of Cytotoxicity by Apigetrin on L6 Cells
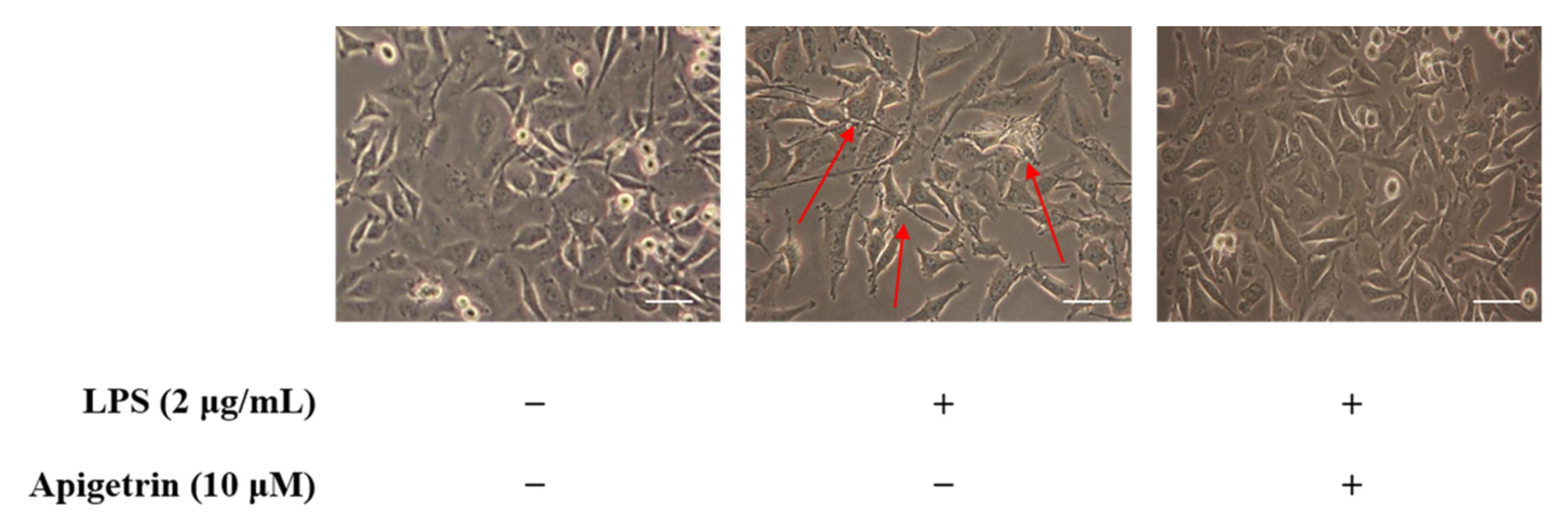
3.2. Observation of Morphological Changes on LPS and Apigetrin-Treated L6 Cells
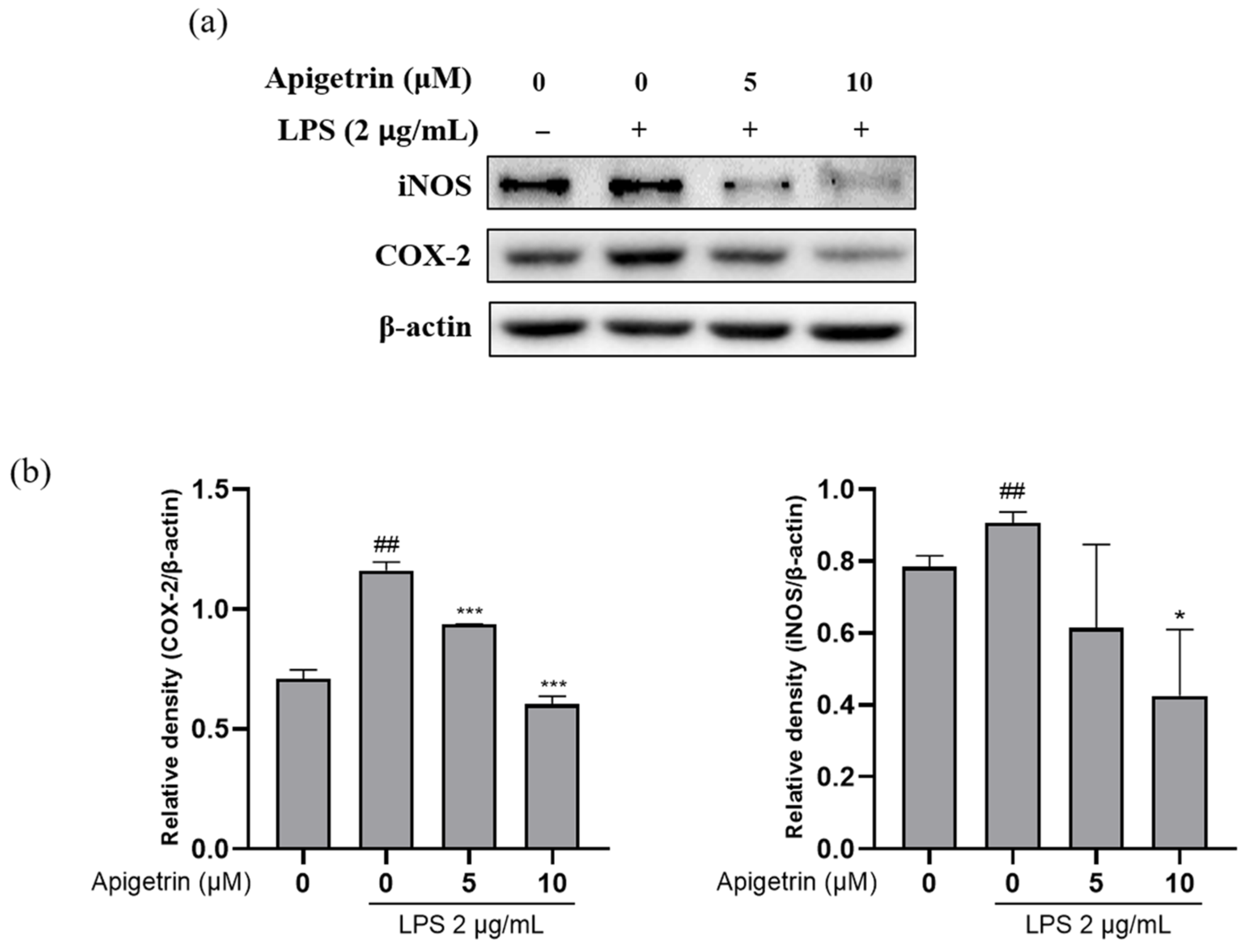
3.3. Apigetrin Suppressed LPS-Induced iNOS and COX-2 Proteins Expression
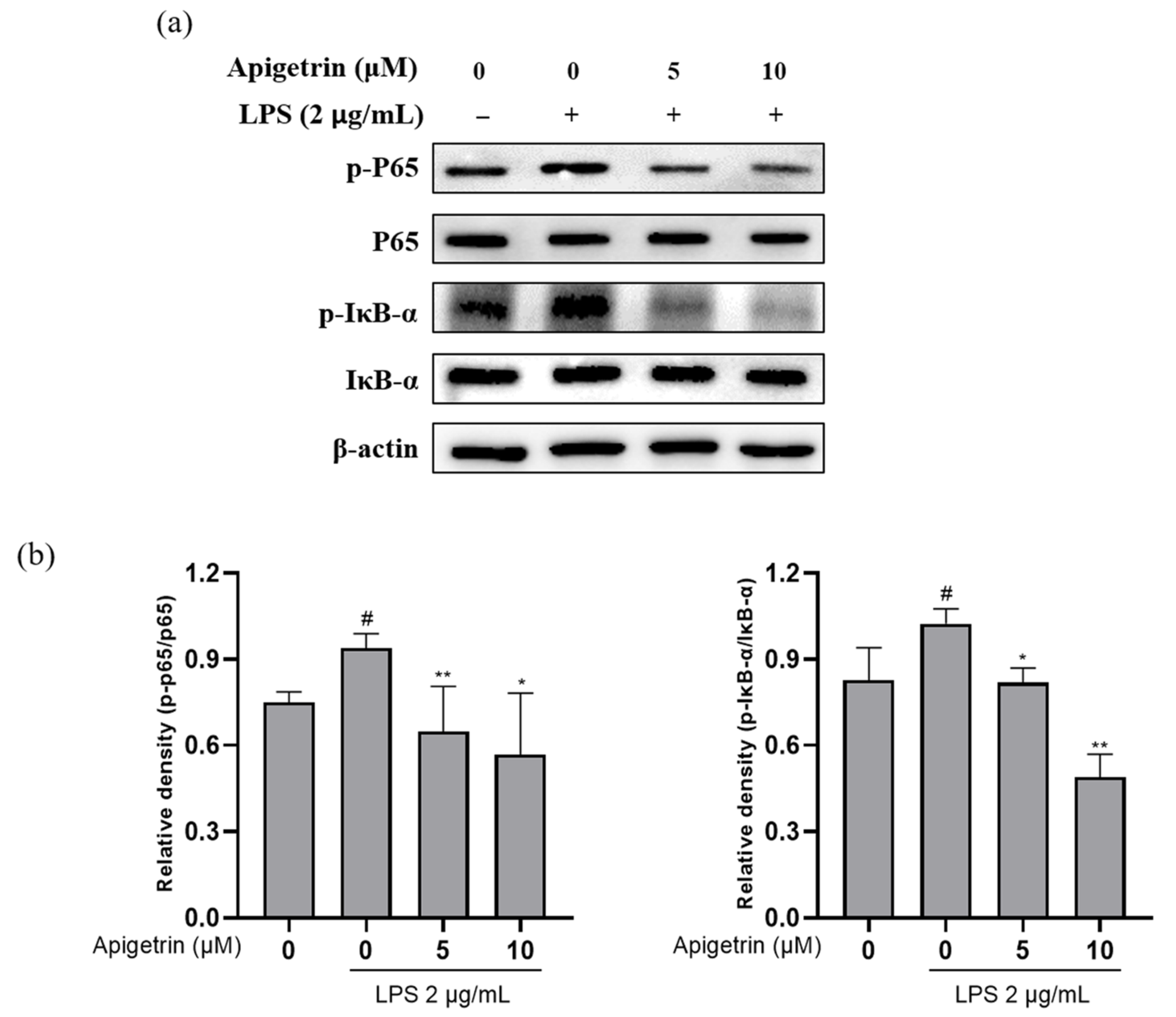
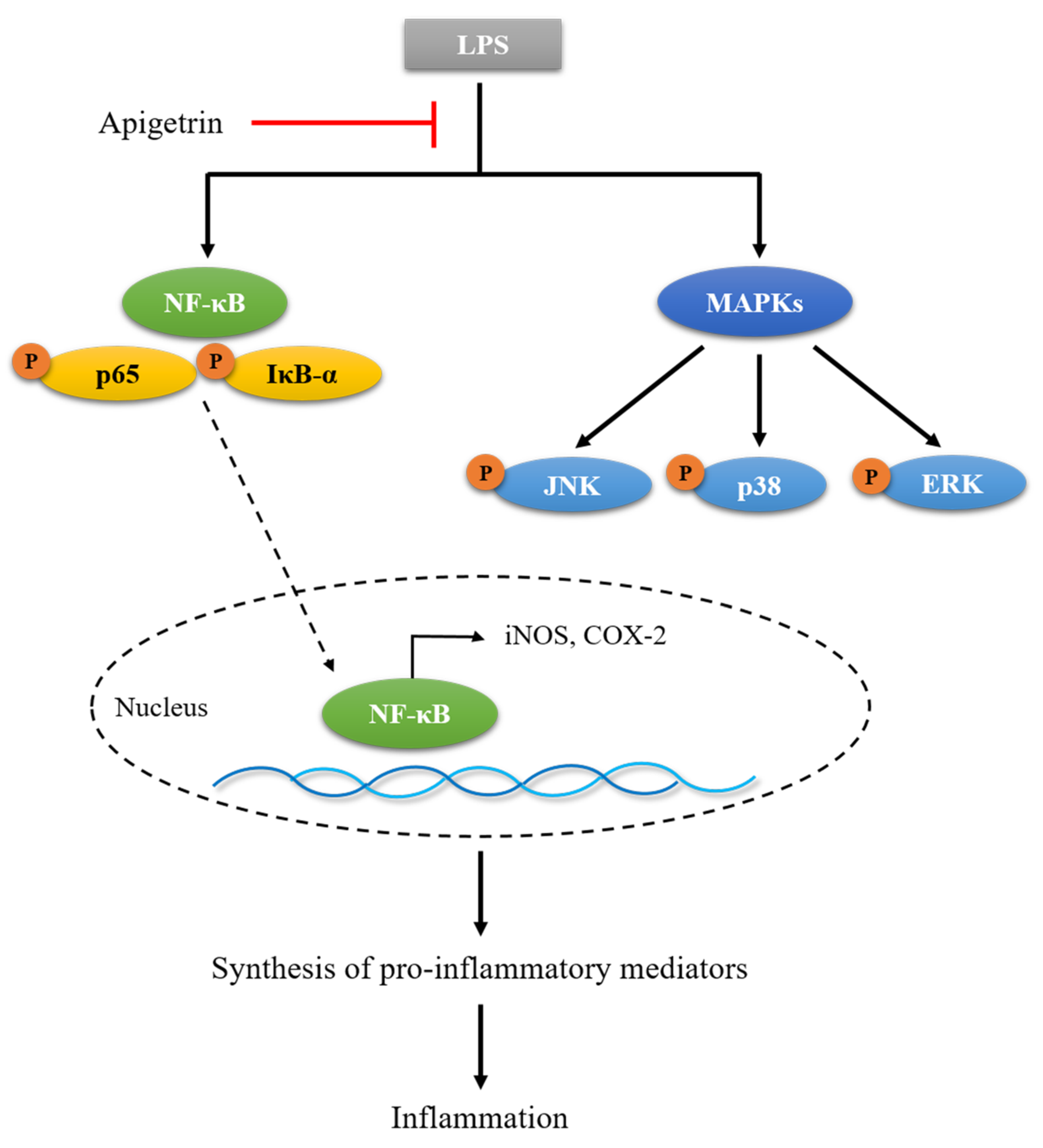
3.4. Effects of Apigetrin on NF-κB Signaling in LPS-Treated L6 Cells
3.5. Effects of Apigetrin on MAPK Phosphorylation in LPS-Treated L6 Cells
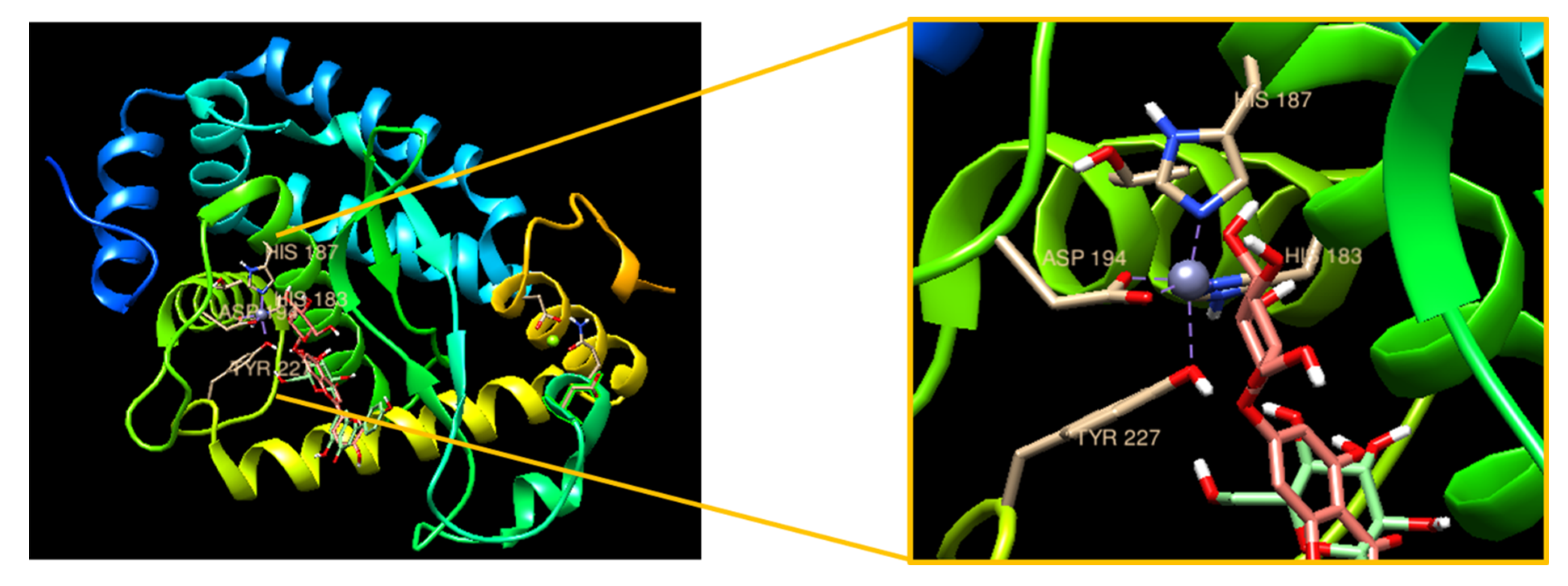
3.6. Molecular Docking with Apigetrin and NF-κB
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Miean, K.H.; Mohamed, S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef] [PubMed]
- Thuan, N.H.; Chaudhary, A.K.; Van Cuong, D.; Cuong, N.X. Engineering co-culture system for production of apigetrin in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2018, 45, 175–185. [Google Scholar] [CrossRef]
- Hadrich, F.; Sayadi, S. Apigetrin inhibits adipogenesis in 3T3-L1 cells by downregulating PPARgamma and CEBP-alpha. Lipids Health Dis. 2018, 17, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Li, M.; Xu, L.J. Apigetrin treatment attenuates LPS-induced acute otitis media though suppressing inflammation and oxidative stress. Biomed. Pharmacother. 2019, 109, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Lu, N.N.; Feng, L. Apigetrin inhibits gastric cancer progression through inducing apoptosis and regulating ROS-modulated STAT3/JAK2 pathway. Biochem. Biophys. Res. Commun. 2018, 498, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Tsolmon, S.; Nakazaki, E.; Han, J.; Isoda, H. Apigetrin induces erythroid differentiation of human leukemia cells K562: Proteomics approach. Mol. Nutr. Food Res. 2011, 55 (Suppl. S1), S93–S102. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Park, B.R.; Moon, S.M.; Shin, S.H.; Kim, J.S.; Kim, D.K.; Kim, C.S. Cynaroside protects human periodontal ligament cells from lipopolysaccharide-induced damage and inflammation through suppression of NF-kappaB activation. Arch. Oral Biol. 2020, 120, 104944. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, X.; Zhang, R.; Koci, M.; Si, D.; Ahmad, B.; Guo, H.; Hou, Y. C-Terminal Amination of a Cationic Anti-Inflammatory Peptide Improves Bioavailability and Inhibitory Activity Against LPS-Induced Inflammation. Front. Immunol. 2020, 11, 618312. [Google Scholar] [CrossRef]
- Podbregar, M.; Lainscak, M.; Prelovsek, O.; Mars, T. Cytokine response of cultured skeletal muscle cells stimulated with proinflammatory factors depends on differentiation stage. Sci. World J. 2013, 2013, 617170. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, H.; Gao, Y.; Meng, Y.; Zhao, X.X.; Pan, S.N. Effects of Dandelion Extract on the Proliferation of Rat Skeletal Muscle Cells and the Inhibition of a Lipopolysaccharide-Induced Inflammatory Reaction. Chin. Med. J. 2018, 131, 1724–1731. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, S.M.; Ha, S.E.; Vetrivel, P.; Saralamma, V.V.G.; Kim, E.H.; Kim, G.S. Sinensetin regulates age-related sarcopenia in cultured primary thigh and calf muscle cells. BMC Complementary Altern. Med. 2019, 19, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santa-Cecilia, F.V.; Socias, B.; Ouidja, M.O.; Sepulveda-Diaz, J.E.; Acuna, L.; Silva, R.L.; Michel, P.P.; Del-Bel, E.; Cunha, T.M.; Raisman-Vozari, R. Doxycycline Suppresses Microglial Activation by Inhibiting the p38 MAPK and NF-kB Signaling Pathways. Neurotox. Res. 2016, 29, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Limtrakul, P.; Yodkeeree, S.; Pitchakarn, P.; Punfa, W. Anti-inflammatory effects of proanthocyanidin-rich red rice extract via suppression of MAPK, AP-1 and NF-kappaB pathways in Raw 264.7 macrophages. Nutr. Res. Pract. 2016, 10, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-kappaB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Amaro-Leal, A.; Shvachiy, L.; Pinto, R.; Geraldes, V.; Rocha, I.; Mota-Filipe, H. Therapeutic effects of IkB kinase inhibitor during systemic inflammation. Int. Immunopharmacol. 2020, 84, 106509. [Google Scholar] [CrossRef]
- Chang, H.C.; Lin, K.H.; Tai, Y.T.; Chen, J.T.; Chen, R.M. Lipoteichoic acid-induced TNF-alpha and IL-6 gene expressions and oxidative stress production in macrophages are suppressed by ketamine through downregulating Toll-like receptor 2-mediated activation oF ERK1/2 and NFkappaB. Shock 2010, 33, 485–492. [Google Scholar] [CrossRef]
- Ha, A.T.; Kim, M.Y.; Cho, J.Y. TAK1/AP-1-Targeted Anti-Inflammatory Effects of Barringtonia augusta Methanol Extract. Molecules 2021, 26, 3053. [Google Scholar] [CrossRef]
- Lim, H.S.; Kim, O.S.; Kim, B.Y.; Jeong, S.J. Apigetrin from Scutellaria baicalensis Georgi Inhibits Neuroinflammation in BV-2 Microglia and Exerts Neuroprotective Effect in HT22 Hippocampal Cells. J. Med. Food 2016, 19, 1032–1040. [Google Scholar] [CrossRef]
- Gutierrez-Venegas, G.; Gonzalez-Rosas, Z. Apigenin reduce lipoteichoic acid-induced inflammatory response in rat cardiomyoblast cells. Arch. Pharm. Res. 2017, 40, 240–249. [Google Scholar] [CrossRef]
- Shiota, C.; Abe, T.; Kawai, N.; Ohno, A.; Teshima-Kondo, S.; Mori, H.; Terao, J.; Tanaka, E.; Nikawa, T. Flavones Inhibit LPS-Induced Atrogin-1/MAFbx Expression in Mouse C2C12 Skeletal Myotubes. J. Nutr. Sci. Vitaminol. 2015, 61, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Muniandy, K.; Gothai, S.; Badran, K.M.H.; Suresh Kumar, S.; Esa, N.M.; Arulselvan, P. Suppression of Proinflammatory Cytokines and Mediators in LPS-Induced RAW 264.7 Macrophages by Stem Extract of Alternanthera sessilis via the Inhibition of the NF-kappaB Pathway. J. Immunol. Res. 2018, 2018, 3430684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-kappaB and IkappaB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Hong, G.E.; Kim, J.A.; Nagappan, A.; Yumnam, S.; Lee, H.J.; Kim, E.H.; Lee, W.S.; Shin, S.C.; Park, H.S.; Kim, G.S. Flavonoids Identified from Korean Scutellaria baicalensis Georgi Inhibit Inflammatory Signaling by Suppressing Activation of NF- kappa B and MAPK in RAW 264.7 Cells. Evid. -Based Complementary Altern. Med. 2013, 2013, 912031. [Google Scholar] [CrossRef] [Green Version]
- Peterson, J.M.; Guttridge, D.C. Skeletal muscle diseases, inflammation, and NF-kappaB signaling: Insights and opportunities for therapeutic intervention. Int. Rev. Immunol. 2008, 27, 375–387. [Google Scholar] [CrossRef]
- Wang, K.S.; Li, J.; Wang, Z.; Mi, C.; Ma, J.; Piao, L.X.; Xu, G.H.; Li, X.; Jin, X. Artemisinin inhibits inflammatory response via regulating NF-kappaB and MAPK signaling pathways. Immunopharmacol. Immunotoxicol. 2017, 39, 28–36. [Google Scholar] [CrossRef]
- Kim, S.M.; Lee, S.J.; Venkatarame Gowda Saralamma, V.; Ha, S.E.; Vetrivel, P.; Desta, K.T.; Choi, J.Y.; Lee, W.S.; Shin, S.C.; Kim, G.S. Polyphenol mixture of a native Korean variety of Artemisia argyi H. (Seomae mugwort) and its antiinflammatory effects. Int. J. Mol. Med. 2019, 44, 1741–1752. [Google Scholar] [CrossRef]
- Martineau, L.C.; Gardiner, P.F. Insight into skeletal muscle mechanotransduction: MAPK activation is quantitatively related to tension. J. Appl. Physiol. 2001, 91, 693–702. [Google Scholar] [CrossRef]
- Cui, Y.; Costa, R.M.; Murphy, G.G.; Elgersma, Y.; Zhu, Y.; Gutmann, D.H.; Parada, L.F.; Mody, I.; Silva, A.J. Neurofibromin regulation of ERK signaling modulates GABA release and learning. Cell 2008, 135, 549–560. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, S.-E.; Bhagwan Bhosale, P.; Kim, H.-H.; Park, M.-Y.; Abusaliya, A.; Kim, G.-S.; Kim, J.-A. Apigetrin Abrogates Lipopolysaccharide-Induced Inflammation in L6 Skeletal Muscle Cells through NF-κB/MAPK Signaling Pathways. Curr. Issues Mol. Biol. 2022, 44, 2635-2645. https://doi.org/10.3390/cimb44060180
Ha S-E, Bhagwan Bhosale P, Kim H-H, Park M-Y, Abusaliya A, Kim G-S, Kim J-A. Apigetrin Abrogates Lipopolysaccharide-Induced Inflammation in L6 Skeletal Muscle Cells through NF-κB/MAPK Signaling Pathways. Current Issues in Molecular Biology. 2022; 44(6):2635-2645. https://doi.org/10.3390/cimb44060180
Chicago/Turabian StyleHa, Sang-Eun, Pritam Bhagwan Bhosale, Hun-Hwan Kim, Min-Yeong Park, Abuyaseer Abusaliya, Gon-Sup Kim, and Jin-A Kim. 2022. "Apigetrin Abrogates Lipopolysaccharide-Induced Inflammation in L6 Skeletal Muscle Cells through NF-κB/MAPK Signaling Pathways" Current Issues in Molecular Biology 44, no. 6: 2635-2645. https://doi.org/10.3390/cimb44060180