
Putative Mechanisms Underlying the Beneficial Effects of Polyphenols in Murine Models of Metabolic Disorders in Relation to Gut Microbiota

 ,
,
Abstract
:1. Introduction
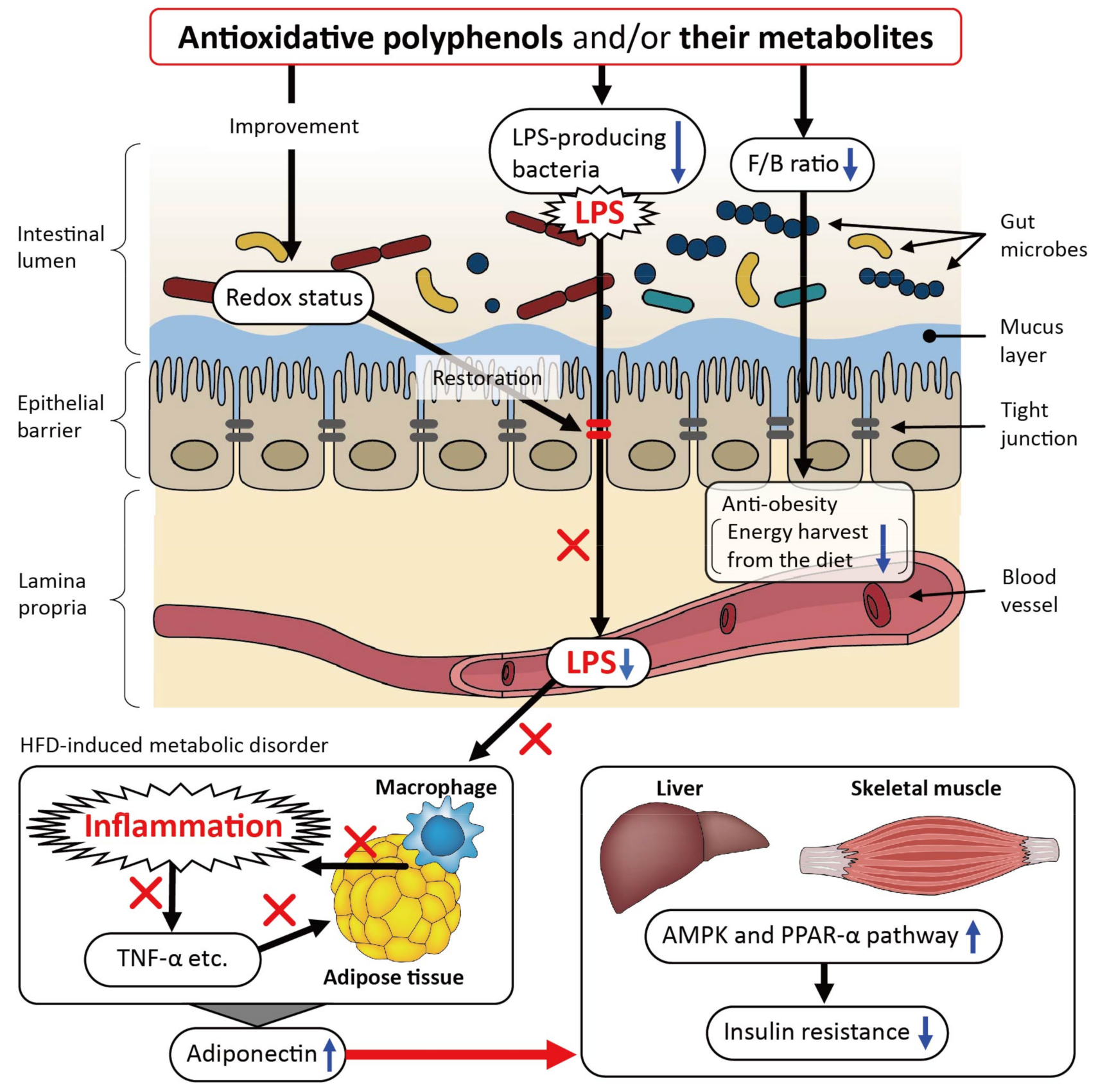
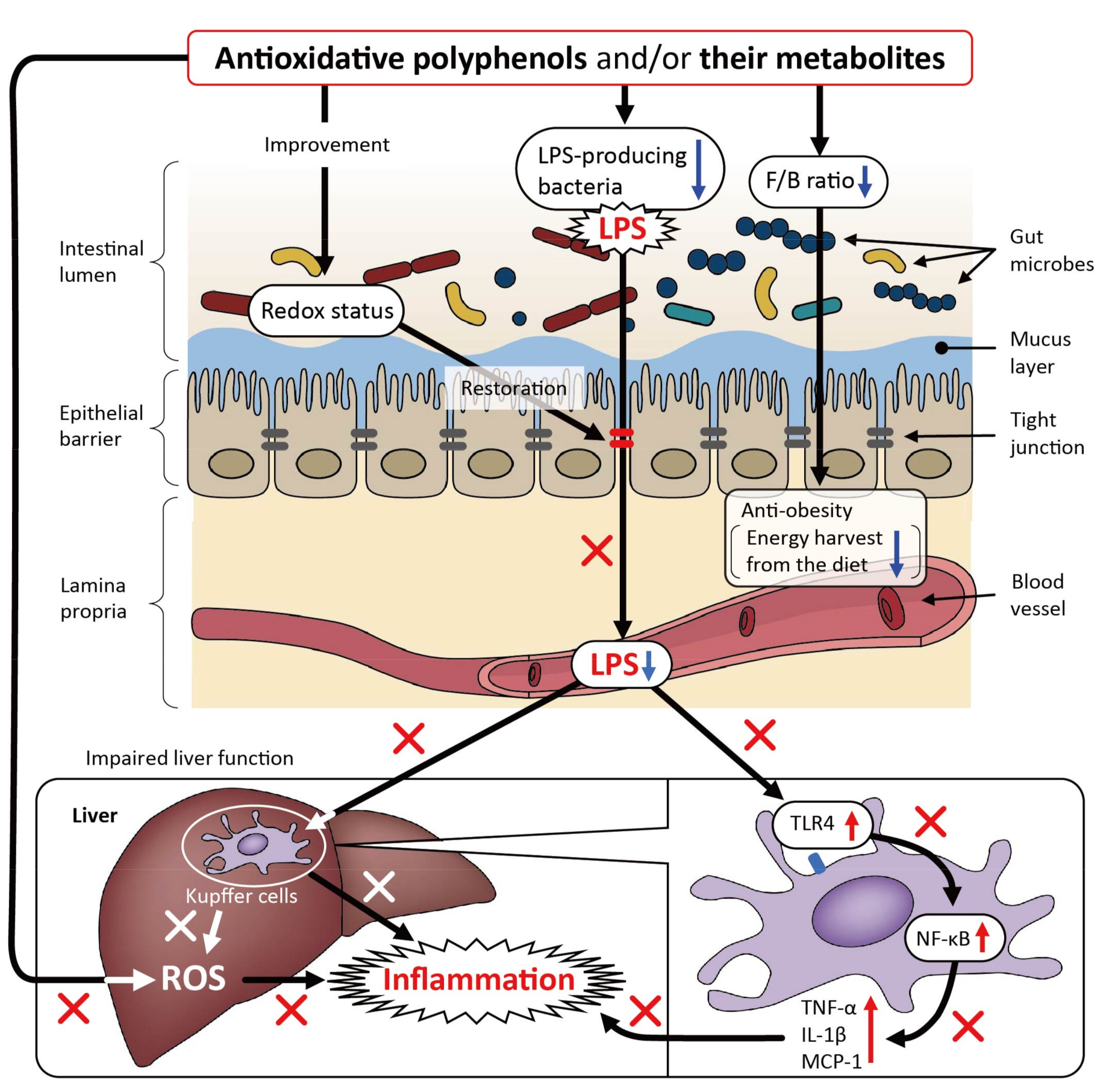
2. Beneficial Effects of Polyphenols on Metabolic Disorders in Relation to the Gut Microbiota in High-Fat Diet (HFD)-Fed Murine Models
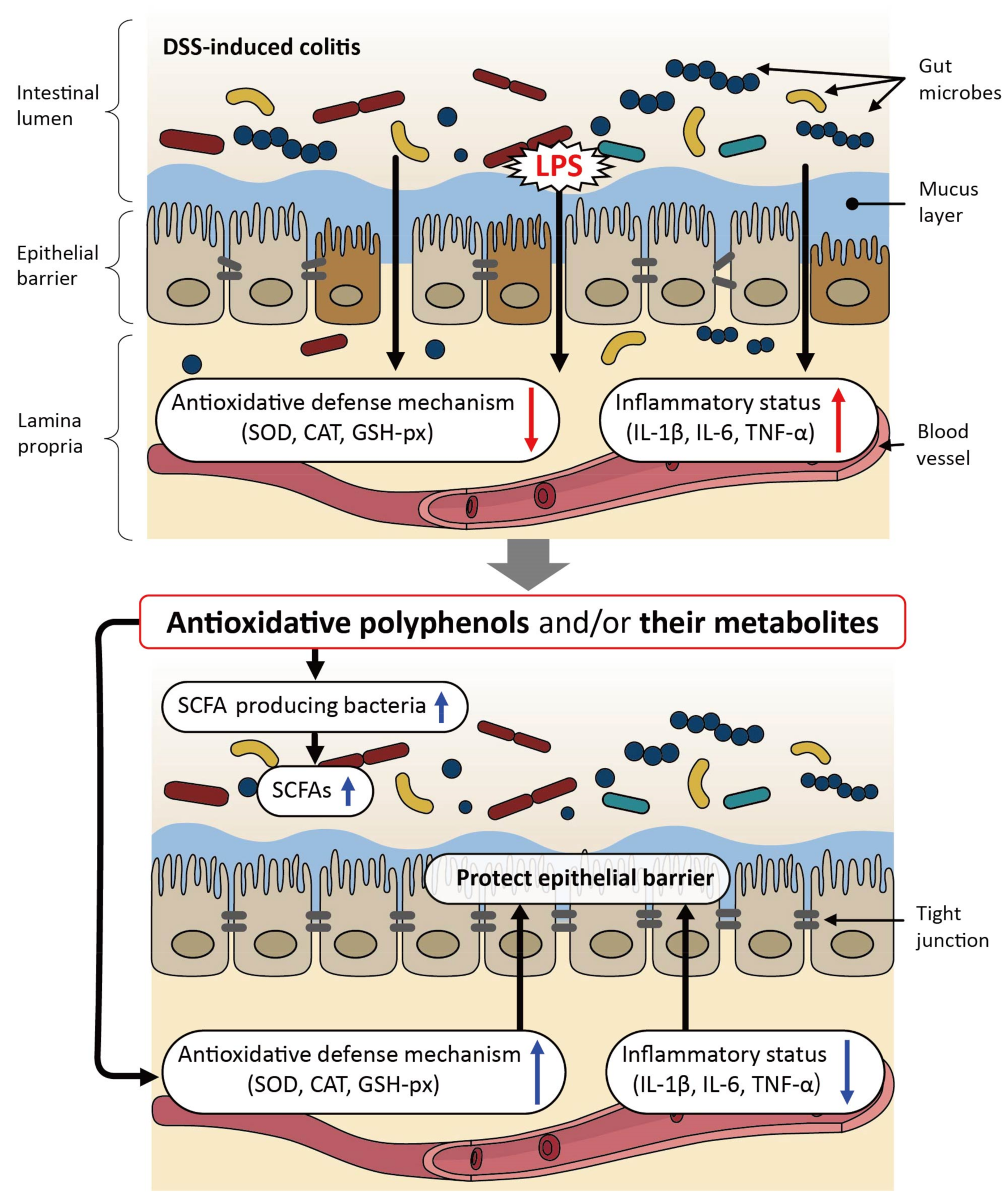
3. Beneficial Effects of Polyphenols on Dextran Sulfate Sodium (DSS)-Induced Colitis in Relation to the Gut Microbiota in Murine Models
4. Beneficial Effects of Polyphenols on Metabolic Disorders Not Associated with HFD or DSS in Relation to the Gut Microbiota in Murine Models
5. Effects of Polyphenols on the Gut Microbiota in Healthy Mice and Rats
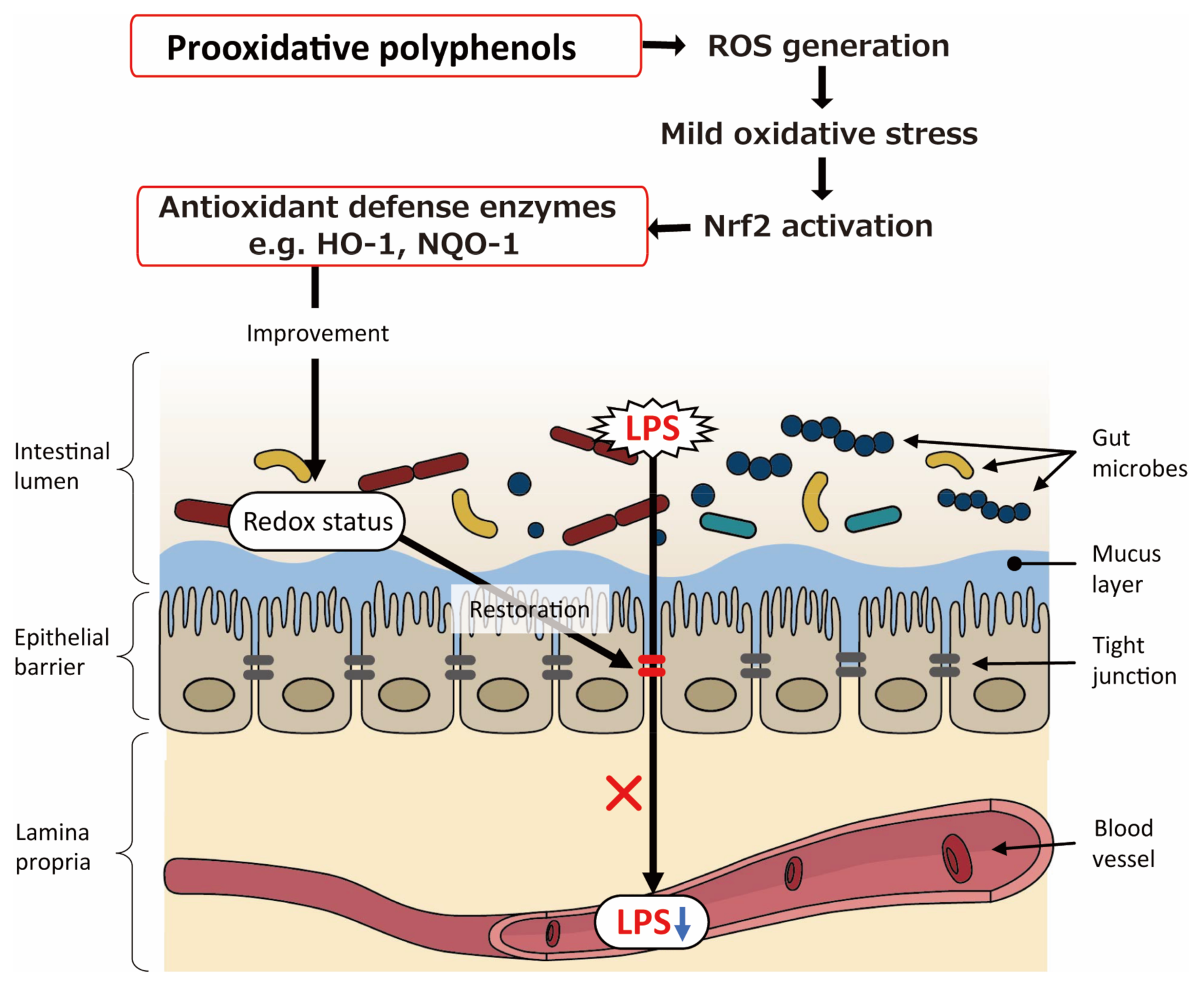
6. Possible Involvement of Prooxidative Potential of Polyphenols in Intestinal Barrier Function
7. Future Perspective on Studies on Interaction of Polyphenols and Gut Microbiota
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marranzano, M.; Rosa, R.L.; Malaguarnera, M.; Palmeri, R.; Tessitori, M.; Barbera, A.C. Polyphenols: Plant sources and food industry applications. Curr. Pharm. Des. 2018, 24, 4125–4130. [Google Scholar] [CrossRef] [PubMed]
- Rajha, H.N.; Paule, A.; Aragonès, G.; Barbosa, M.; Caddeo, C.; Debs, E.; Dinkova, R.; Eckert, G.P.; Fontana, A.; Gebrayel, P.; et al. Recent advances in research on polyphenols: Effects on microbiota, metabolism, and health. Mol. Nutr. Food Res. 2021, 66, e2100670. [Google Scholar] [CrossRef] [PubMed]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, A.; Biagi, M.; Corsini, M.; Baini, G.; Cappellucci, G.; Miraldi, E. Polyphenols: From theory to practice. Foods 2021, 10, 2595. [Google Scholar] [CrossRef]
- Iglesias-Aguirre, C.E.; Cortés-Martín, A.; Ávila-Gálvez, M.; Giménez-Bastida, J.A.; Selma, M.V.; González-Sarrías, A.; Espín, J.C. Main drivers of (poly)phenol effects on human health: Metabolite production and/or gut microbiota-associated metabotypes? Food Funct. 2021, 12, 10324–10355. [Google Scholar] [CrossRef]
- Parida, S.; Sharma, D. The microbiome-estrogen connection and breast cancer risk. Cells 2019, 8, 1642. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Raka, F.; Adeli, K. The role of the gut microbiota in lipid and lipoprotein metabolism. J. Clin. Med. 2019, 8, 2227. [Google Scholar] [CrossRef] [Green Version]
- The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- The Integrative HMP (iHMP) Research Network Consortium. The integrative human microbiome project: Dynamic analysis of microbiome-host omics profiles during periods of human health and disease. Cell Host Microbe 2014, 16, 276–289. [Google Scholar] [CrossRef] [Green Version]
- Saint-Georges-Chaumet, Y.; Edeas, M. Microbiota-mitochondria inter-talk: Consequence for microbiota-host interaction. Pathog. Dis. 2016, 74, ftv096. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Jiang, Q. Roles of the polyphenol-gut microbiota interaction in alleviating colitis and preventing colitis-associated colorectal cancer. Adv. Nutr. 2021, 12, 546–565. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Van Hée, V.F.; Bindels, L.B.; De Backer, F.; Cani, P.D.; Delzenne, N.M. Polyphenol-rich extract of pomegranate peel alleviates tissue inflammation and hypercholesterolaemia in high-fat diet-induced obese mice: Potential implication of the gut microbiota. Br. J. Nutr. 2013, 109, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowan, T.E.; Palmnäs, M.S.; Yang, J.; Bomhof, M.R.; Ardell, K.L.; Reimer, R.A.; Vogel, H.J.; Shearer, J. Chronic coffee consumption in the diet-induced obese rat: Impact on gut microbiota and serum metabolomics. J. Nutr. Biochem. 2014, 25, 489–495. [Google Scholar] [CrossRef]
- Anhê, F.F.; Roy, D.; Pilon, G.; Dudonné, S.; Matamoros, S.; Varin, T.V.; Garofalo, C.; Moine, Q.; Desjardins, Y.; Levy, E.; et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 2015, 64, 872–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, B.; Hoffman, J.; Martinez, K.; Grace, M.; Lila, M.A.; Cockrell, C.; Nadimpalli, A.; Chang, E.; Chuang, C.C.; Zhong, W.; et al. A polyphenol-rich fraction obtained from table grapes decreases adiposity, insulin resistance and markers of inflammation and impacts gut microbiota in high-fat-fed mice. J. Nutr. Biochem. 2016, 31, 150–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zeng, B.; Zhang, X.; Liao, Z.; Gu, L.; Liu, Z.; Zhong, Q.; Wei, H.; Fang, X. The effect of green tea polyphenols on gut microbial diversity and fat deposition in C57BL/6J HFA mice. Food Funct. 2016, 7, 4956–4966. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Gao, X.; Xie, Q.; Kong, P.; Liu, L.; Sun, S.; Xiong, B.; Huang, B.; Yan, L.; Sheng, J.; Xiang, H. Polyphenol- and caffeine-rich postfermented Pu-erh tea improves diet-induced metabolic syndrome by remodeling intestinal homeostasis in mice. Infect. Immun. 2018, 86, e00601-17. [Google Scholar] [CrossRef] [Green Version]
- Van Hul, M.; Geurts, L.; Plovier, H.; Druart, C.; Everard, A.; Ståhlman, M.; Rhimi, M.; Chira, K.; Teissedre, P.L.; Delzenne, N.M.; et al. Reduced obesity, diabetes, and steatosis upon cinnamon and grape pomace are associated with changes in gut microbiota and markers of gut barrier. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E334–E352. [Google Scholar] [CrossRef]
- Wu, S.; Hu, R.; Nakano, H.; Chen, K.; Liu, M.; He, X.; Zhang, H.; He, J.; Hou, D.X. Modulation of gut microbiota by Lonicera caerulea L. berry polyphenols in a mouse model of fatty liver induced by high fat diet. Molecules 2018, 23, 3213. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Deng, Q.; Xu, J.; Wang, X.; Hu, C.; Tang, H.; Huang, F. Sinapic acid and resveratrol alleviate oxidative stress with modulation of gut microbiota in high-fat diet-fed rats. Food Res. Int. 2019, 116, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, J.; Su, Q.; Liu, Y. Sinapine reduces non-alcoholic fatty liver disease in mice by modulating the composition of the gut microbiota. Food Funct. 2019, 10, 3637–3649. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zhang, B.; Hu, Y.; Wang, J.; Liu, J.; Qin, R.; Lv, S.; Wang, S. Correlation analysis of intestinal redox state with the gut microbiota reveals the positive intervention of tea polyphenols on hyperlipidemia in high fat diet fed mice. J. Agric. Food Chem. 2019, 67, 7325–7335. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zheng, Y.; Zhou, J.; Geng, Y.; Zou, P.; Li, Y.; Zhang, C. Polyphenol-rich extracts from brown macroalgae Lessonia trabeculate attenuate hyperglycemia and modulate gut microbiota in high-fat diet and streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2019, 67, 12472–12480. [Google Scholar] [CrossRef]
- Zhao, R.; Long, X.; Yang, J.; Du, L.; Zhang, X.; Li, J.; Hou, C. Pomegranate peel polyphenols reduce chronic low-grade inflammatory responses by modulating gut microbiota and decreasing colonic tissue damage in rats fed a high-fat diet. Food Funct. 2019, 10, 8273–8285. [Google Scholar] [CrossRef]
- Wang, P.; Li, D.; Ke, W.; Liang, D.; Hu, X.; Chen, F. Resveratrol-induced gut microbiota reduces obesity in high-fat diet-fed mice. Int. J. Obes. 2020, 44, 213–225. [Google Scholar] [CrossRef]
- Li, B.; Cheng, Z.; Sun, X.; Si, X.; Gong, E.; Wang, Y.; Tian, J.; Shu, C.; Ma, F.; Li, D.; et al. Lonicera caerulea L. polyphenols alleviate oxidative stress-induced intestinal environment imbalance and lipopolysaccharide-induced liver injury in HFD-fed rats by regulating the Nrf2/HO-1/NQO1 and MAPK pathways. Mol. Nutr. Food Res. 2020, 64, e1901315. [Google Scholar] [CrossRef]
- Wang, P.; Wang, J.; Li, D.; Ke, W.; Chen, F.; Hu, X. Targeting the gut microbiota with resveratrol: A demonstration of novel evidence for the management of hepatic steatosis. J. Nutr. Biochem. 2020, 81, 108363. [Google Scholar] [CrossRef]
- Wang, P.; Gao, J.; Ke, W.; Wang, J.; Li, D.; Liu, R.; Jia, Y.; Wang, X.; Chen, X.; Chen, F.; et al. Resveratrol reduces obesity in high-fat diet-fed mice via modulating the composition and metabolic function of the gut microbiota. Free Radic. Biol. Med. 2020, 156, 83–98. [Google Scholar] [CrossRef]
- Du, P.; Zhou, J.; Zhang, L.; Zhang, J.; Li, N.; Zhao, C.; Tu, L.; Zheng, Y.; Xia, T.; Luo, J.; et al. GC × GC-MS analysis and hypolipidemic effects of polyphenol extracts from Shanxi-aged vinegar in rats under a high fat diet. Food Funct. 2020, 11, 7468–7480. [Google Scholar] [CrossRef]
- Hu, D.; Yang, W.; Mao, P.; Cheng, M. Combined amelioration of prebiotic resveratrol and probiotic Bifidobacteria on obesity and nonalcoholic fatty liver disease. Nutr. Cancer 2021, 73, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Sinisgalli, C.; Vezza, T.; Diez-Echave, P.; Ostuni, A.; Faraone, I.; Hidalgo-Garcia, L.; Russo, D.; Armentano, M.F.; Garrido-Mesa, J.; Rodriguez-Cabezas, M.E.; et al. The beneficial effects of red sun-dried Capsicum annuum L. Cv Senise extract with antioxidant properties in experimental obesity are associated with modulation of the intestinal microbiota. Mol. Nutr. Food Res. 2021, 65, e2000812. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca Cardoso, L.M.; de Souza Monnerat, J.A.; de Medeiros Silva, I.W.S.; da Silva Ferreira Fiochi, R.; da Matta Alvarez Pimenta, N.; Mota, B.F.; Dolisnky, M.; do Carmo, F.L.; Barroso, S.G.; da Costa, C.A.S.; et al. Beverages rich in resveratrol and physical activity attenuate metabolic changes induced by high-fat diet. J. Am. Coll. Nutr. 2021, 40, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Li, Y.L.; Zhang, X.; Wang, K.B.; Huang, J.A.; Liu, Z.H.; Zhu, M.Z. Polyphenols from Fu brick tea reduce obesity via modulation of gut microbiota and gut microbiota-related intestinal oxidative stress and barrier function. J. Agric. Food Chem. 2021, 69, 14530–14543. [Google Scholar] [CrossRef]
- Morbidelli, L. Polyphenol-based nutraceuticals for the control of angiogenesis: Analysis of the critical issues for human use. Pharmacol. Res. 2016, 111, 384–393. [Google Scholar] [CrossRef]
- Carrera-Quintanar, L.; López Roa, R.I.; Quintero-Fabián, S.; Sánchez-Sánchez, M.A.; Vizmanos, B.; Ortuño-Sahagún, D. Phytochemicals that influence gut microbiota as prophylactics and for the treatment of obesity and inflammatory diseases. Mediat. Inflamm. 2018, 2018, 9734845. [Google Scholar] [CrossRef]
- Tong, X.; Xu, J.; Lian, F.; Yu, X.; Zhao, Y.; Xu, L.; Zhang, M.; Zhao, X.; Shen, J.; Wu, S.; et al. Structural alteration of gut microbiota during the amelioration of human type 2 diabetes with hyperlipidemia by metformin and a traditional Chinese herbal frmula: A multicenter, randomized, open label clinical trial. mBio 2018, 9, e02392-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, P.; Li, D.; Hu, X.; Chen, F. Beneficial effects of ginger on prevention of obesity through modulation of gut microbiota in mice. Eur. J. Nutr. 2020, 59, 699–718. [Google Scholar] [CrossRef]
- Sellmann, C.; Priebs, J.; Landmann, M.; Degen, C.; Engstler, A.J.; Jin, C.J.; Gärttner, S.; Spruss, A.; Huber, O.; Bergheim, I. Diets rich in fructose, fat or fructose and fat alter intestinal barrier function and lead to the development of nonalcoholic fatty liver disease over time. J. Nutr. Biochem. 2015, 26, 1183–1192. [Google Scholar] [CrossRef]
- Sellmann, C.; Degen, C.; Jin, C.J.; Nier, A.; Engstler, A.J.; Hasan Alkhatib, D.; De Bandt, J.P.; Bergheim, I. Oral arginine supplementation protects female mice from the onset of non-alcoholic steatohepatitis. Amino Acids 2017, 49, 1215–1225. [Google Scholar] [CrossRef] [Green Version]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Xia, X.; Tang, R.; Zhou, J.; Zhao, H.; Wang, K. Development of a real-time PCR method for Firmicutes and Bacteroidetes in faeces and its application to quantify intestinal population of obese and lean pigs. Lett. Appl. Microbiol. 2008, 47, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.S.; Prabhu, S.; Landau, J. Prevention of carcinogenesis by tea polyphenols. Drug Metab. Rev. 2001, 33, 237–253. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Feng, X.; Li, Q.; Wang, Y.; Li, Q.; Hua, M. Adiponectin, TNF-α and inflammatory cytokines and risk of type 2 diabetes: A systematic review and meta-analysis. Cytokine 2016, 86, 100–109. [Google Scholar] [CrossRef]
- Kersten, S.; Desvergne, B.; Wahli, W. Roles of PPARs in health and disease. Nature 2000, 405, 421–424. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef]
- Tomas, E.; Tsao, T.S.; Saha, A.K.; Murrey, H.E.; Zhang, C.C.; Itani, S.I.; Lodish, H.F.; Ruderman, N.B. Enhanced muscle fat oxidation and glucose transport by ACRP30 globular domain: Acetyl-CoA carboxylase inhibition and AMP-activated protein kinase activation. Proc. Natl. Acad. Sci. USA 2002, 99, 16309–16313. [Google Scholar] [CrossRef] [Green Version]
- Kahn, B.B.; Alquier, T.; Carling, D.; Hardie, D.G. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2005, 1, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada-Iwabu, M.; Yamauchi, T.; Iwabu, M.; Honma, T.; Hamagami, K.; Matsuda, K.; Yamaguchi, M.; Tanabe, H.; Kimura-Someya, T.; Shirouzu, M.; et al. A small-molecule AdipoR agonist for type 2 diabetes and short life in obesity. Nature 2013, 503, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Kostic, A.D.; Xavier, R.J.; Gevers, D. The microbiome in inflammatory bowel disease: Current status and the future ahead. Gastroenterology 2014, 146, 1489–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danese, S.; Fiocchi, C.; Panés, J. Drug development in IBD: From novel target identification to early clinical trials. Gut 2016, 65, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wu, X.; Cao, S.; Cromie, M.; Shen, Y.; Feng, Y.; Yang, H.; Li, L. Chlorogenic acid ameliorates experimental colitis by promoting growth of Akkermansia in mice. Nutrients 2017, 9, 677. [Google Scholar] [CrossRef] [Green Version]
- Ohno, M.; Nishida, A.; Sugitani, Y.; Nishino, K.; Inatomi, O.; Sugimoto, M.; Kawahara, M.; Andoh, A. Nanoparticle curcumin ameliorates experimental colitis via modulation of gut microbiota and induction of regulatory T cells. PLoS ONE 2017, 12, e0185999. [Google Scholar] [CrossRef] [Green Version]
- Scarano, A.; Butelli, E.; De Santis, S.; Cavalcanti, E.; Hill, L.; De Angelis, M.; Giovinazzo, G.; Chieppa, M.; Martin, C.; Santino, A. Combined dietary anthocyanins, flavonols, and stilbenoids alleviate inflammatory bowel disease symptoms in mice. Front. Nutr. 2017, 4, 75. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Jin, X.; Li, Q.; Sawaya, A.; Le Leu, R.K.; Conlon, M.A.; Wu, L.; Hu, F. Propolis from different geographic origins decreases intestinal inflammation and Bacteroides spp. populations in a model of DSS-induced colitis. Mol. Nutr. Food Res. 2018, 62, e1800080. [Google Scholar] [CrossRef]
- Hong, Z.; Piao, M. Effect of quercetin monoglycosides on oxidative stress and gut microbiota diversity in mice with dextran sodium sulphate-induced colitis. BioMed Res. Int. 2018, 2018, 8343052. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, H.; Cheng, N.; Cao, W. Rape bee pollen alleviates dextran sulfate sodium (DSS)-induced colitis by neutralizing IL-1β and regulating the gut microbiota in mice. Food Res. Int. 2019, 122, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Cheng, N.; Zhou, W.; Chen, S.; Wang, Q.; Gao, H.; Xue, X.; Wu, L.; Cao, W. Honey polyphenols ameliorate DSS-induced ulcerative colitis via modulating gut microbiota in rats. Mol. Nutr. Food Res. 2019, 63, e1900638. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Hu, M.; Zhang, L.; Gao, Y.; Ma, L.; Xu, Q. Dietary taxifolin protects against dextran sulfate sodium-induced colitis via NF-κB signaling, enhancing intestinal barrier and modulating gut microbiota. Front. Immunol. 2021, 11, 631809. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Huang, S.; Li, T.; Li, N.; Han, D.; Zhang, B.; Xu, Z.Z.; Zhang, S.; Pang, J.; Wang, S.; et al. Gut microbiota from green tea polyphenol-dosed mice improves intestinal epithelial homeostasis and ameliorates experimental colitis. Microbiome 2021, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Tsao, R.; Li, C.; Wang, X.; Zhang, H.; Jiang, L.; Sun, Y.; Xiong, H. Green pea (Pisum sativum L.) hull polyphenol extracts ameliorate DSS-induced colitis through Keap1/Nrf2 pathway and gut microbiota modulation. Foods 2021, 10, 2765. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, H.; Zhao, Q.; Wang, X.; Zhang, J.; Zhao, X. Polyphenol-rich loquat fruit extract prevents fructose-induced nonalcoholic fatty liver disease by modulating glycometabolism, lipometabolism, oxidative stress, inflammation, intestinal barrier, and gut microbiota in mice. J. Agric. Food Chem. 2019, 67, 7726–7737. [Google Scholar] [CrossRef]
- Xie, K.; He, X.; Chen, K.; Sakao, K.; Hou, D.X. Ameliorative effects and molecular mechanisms of vine tea on western diet-induced NAFLD. Food Funct. 2020, 11, 5976–5991. [Google Scholar] [CrossRef]
- Xia, T.; Duan, W.; Zhang, Z.; Li, S.; Zhao, Y.; Geng, B.; Zheng, Y.; Yu, J.; Wang, M. Polyphenol-rich vinegar extract regulates intestinal microbiota and immunity and prevents alcohol-induced inflammation in mice. Food Res. Int. 2021, 140, 110064. [Google Scholar] [CrossRef]
- Zhao, L.; Mehmood, A.; Soliman, M.M.; Iftikhar, A.; Iftikhar, M.; Aboelenin, S.M.; Wang, C. Protective effects of ellagic acid against alcoholic lver disease in mice. Front. Nutr. 2021, 8, 744520. [Google Scholar] [CrossRef]
- Kong, Y.; Yan, T.; Tong, Y.; Deng, H.; Tan, C.; Wan, M.; Wang, M.; Meng, X.; Wang, Y. Gut microbiota modulation by polyphenols from Aronia melanocarpa of LPS-induced liver diseases in rats. J. Agric. Food Chem. 2021, 69, 3312–3325. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, L.; Chen, R.; Zhao, Y.; Ren, D.; Yang, X. Chlorogenic acid inhibits trimethylamine-N-oxide formation and remodels intestinal microbiota to alleviate liver dysfunction in high L-carnitine feeding mice. Food Funct. 2021, 12, 10500–10511. [Google Scholar] [CrossRef]
- Mezhibovsky, E.; Knowles, K.A.; He, Q.; Sui, K.; Tveter, K.M.; Duran, R.M.; Roopchand, D.E. Grape polyphenols attenuate diet-induced obesity and hepatic steatosis in mice in association with reduced butyrate and increased markers of intestinal carbohydrate oxidation. Front. Nutr. 2021, 8, 675267. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Chen, H.P.; He, Y.Y.; Chen, W.L.; Chen, J.W.; Gao, L.; Hu, H.Y.; Wang, J. Effects of rich-polyphenols extract of Dendrobium loddigesii on anti-diabetic, anti-inflammatory, anti-oxidant, and gut microbiota modulation in db/db mice. Molecules 2018, 23, 3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Ma, Y.; Liu, Z.; Liu, L.; Yang, K.; Wei, Y.; Liu, Y.; Chen, X.; Sun, X.; Wen, D. Hydroxytyrosol prevents PM(2.5)-induced adiposity and insulin resistance by restraining oxidative stress related NF-κB pathway and modulation of gut microbiota in a murine model. Free Radic. Biol. Med. 2019, 141, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Guirro, M.; Gual-Grau, A.; Gibert-Ramos, A.; Alcaide-Hidalgo, J.M.; Canela, N.; Arola, L.; Mayneris-Perxachs, J. Metabolomics elucidates dose-dependent molecular beneficial effects of hesperidin supplementation in rats fed an obesogenic diet. Antioxidants 2020, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Liu, T.; Yin, L.; Man, S.; Ye, S.; Ma, L. Polyphenol-rich extract from Litchi chinensis seeds alleviates hypertension-induced renal damage in rats. J. Agric. Food Chem. 2021, 69, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, N.; Kan, J.; Tang, S.; Sun, R.; Wang, Z.; Chen, M.; Liu, J.; Jin, C. Polyphenols from Arctium lappa L ameliorate doxorubicin-induced heart failure and improve gut microbiota composition in mice. J. Food Biochem. 2021, 46, e13731. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, X.; Zhao, H.; Yang, E.; Wang, Y.; Cheng, N.; Cao, W. Impact of Camellia japonica bee pollen polyphenols on hyperuricemia and gut microbiota in potassium oxonate-induced mice. Nutrients 2021, 13, 2665. [Google Scholar] [CrossRef] [PubMed]
- Da Silva-Maia, J.K.; Batista, A.G.; Correa, L.C.; Lima, G.C.; Bogusz Junior, S.; Maróstica Junior, M.R. Aqueous extract of berry (Plinia jaboticaba) byproduct modulates gut microbiota and maintains the balance on antioxidant defense system in rats. J. Food Biochem. 2019, 43, e12705. [Google Scholar] [CrossRef]
- Alim, A.; Li, T.; Nisar, T.; Ren, D.; Liu, Y.; Yang, X. Consumption of two whole kiwifruit (Actinide chinensis) per day improves lipid homeostasis, fatty acid metabolism and gut microbiota in healthy rats. Int. J. Biol. Macromol. 2020, 156, 186–195. [Google Scholar] [CrossRef]
- Yin, J.; Ren, W.; Wei, B.; Huang, H.; Li, M.; Wu, X.; Wang, A.; Xiao, Z.; Shen, J.; Zhao, Y.; et al. Characterization of chemical composition and prebiotic effect of a dietary medicinal plant Penthorum chinense Pursh. Food Chem. 2020, 319, 126568. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, Y.; Wan, P.; Dong, W.; Huang, K.; Ran, L.; Mi, J.; Lu, L.; Zeng, X.; Cao, Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 108952. [Google Scholar] [CrossRef] [PubMed]
- Likotrafiti, E.; Rhoades, J. Probiotics, prebiotics, synbiotics, and foodborne illness. In Bioactive Foods in Health Promotion: Probiotics, Prebiotics, and Synbiotics; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 469–476. [Google Scholar]
- Hollman, P.C.; Cassidy, A.; Comte, B.; Heinonen, M.; Richelle, M.; Richling, E.; Serafini, M.; Scalbert, A.; Sies, H.; Vidry, S. The biological relevance of direct antioxidant effects of polyphenols for cardiovascular health in humans is not established. J. Nutr. 2011, 141, 989s–1009s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croft, K.D. Dietary polyphenols: Antioxidants or not? Arch. Biochem. Biophys. 2016, 595, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, H.; Maeda, M.; Okubo, S.; Shimamura, T. Role of hydrogen peroxide in bactericidal action of catechin. Biol. Pharm. Bull. 2004, 27, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H.Y.; Zubair, H.; Faisal, M.; Ullah, M.F.; Farhan, M.; Sarkar, F.H.; Ahmad, A.; Hadi, S.M. Plant polyphenol induced cell death in human cancer cells involves mobilization of intracellular copper ions and reactive oxygen species generation: A mechanism for cancer chemopreventive action. Mol. Nutr. Food Res. 2014, 58, 437–446. [Google Scholar] [CrossRef]
- Leonard, M.O.; Kieran, N.E.; Howell, K.; Burne, M.J.; Varadarajan, R.; Dhakshinamoorthy, S.; Porter, A.G.; O’Farrelly, C.; Rabb, H.; Taylor, C.T. Reoxygenation-specific activation of the antioxidant transcription factor Nrf2 mediates cytoprotective gene expression in ischemia-reperfusion injury. FASEB J. 2006, 20, 2624–2626. [Google Scholar] [CrossRef]
- Shelton, L.M.; Park, B.K.; Copple, I.M. Role of Nrf2 in protection against acute kidney injury. Kidney Int. 2013, 84, 1090–1095. [Google Scholar] [CrossRef]
- Smith, R.E.; Tran, K.; Smith, C.C.; McDonald, M.; Shejwalkar, P.; Hara, K. The role of the Nrf2/ARE antioxidant system in preventing cardiovascular diseases. Diseases 2016, 4, 34. [Google Scholar] [CrossRef]
- Milutinović, M.; Dimitrijević-Branković, S.; Rajilić-Stojanović, M. Plant Extracts Rich in Polyphenols as Potent Modulators in the Growth of Probiotic and Pathogenic Intestinal Microorganisms. Front. Nutr. 2021, 8, 688843. [Google Scholar] [CrossRef]
- Cressey, D. UK funders demand strong statistics for animal studies. Nature 2015, 520, 271–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, F.S.; Tabak, L.A. Policy: NIH plans to enhance reproducibility. Nature 2014, 505, 612–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahowald, M.A.; Rey, F.E.; Seedorf, H.; Turnbaugh, P.J.; Fulton, R.S.; Wollam, A.; Shah, N.; Wang, C.; Magrini, V.; Wilson, R.K.; et al. Characterizing a model human gut microbiota composed of members of its two dominant bacterial phyla. Proc. Natl. Acad. Sci. USA 2009, 106, 5859–5864. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [Green Version]
- Jin, G.; Asou, Y.; Ishiyama, K.; Okawa, A.; Kanno, T.; Niwano, Y. Proanthocyanidin-Rich Grape Seed Extract Modulates Intestinal Microbiota in Ovariectomized Mice. J. Food Sci. 2018, 83, 1149–1152. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Yang, J.; Zhuge, A.; Li, L.; Ni, S. Gut microbiota modulates osteoclast glutathione synthesis and mitochondrial biogenesis in mice subjected to ovariectomy. Cell Prolif. 2022, 55, e13194. [Google Scholar] [CrossRef]
- Rowland, I.; Faughnan, M.; Hoey, L.; Wähälä, K.; Williamson, G.; Cassidy, A. Bioavailability of phyto-oestrogens. Br. J. Nutr. 2003, 89 (Suppl. 1), S45–S58. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [Green Version]
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut Microbes 2016, 7, 216–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, S.; Suzuki, T. Isolation and characterization of a novel equol-producing bacterium from human feces. Biosci. Biotechnol. Biochem. 2008, 72, 2660–2666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthies, A.; Blaut, M.; Braune, A. Isolation of a human intestinal bacterium capable of daidzein and genistein conversion. Appl. Environ. Microbiol. 2009, 75, 1740–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setchell, K.D.; Clerici, C. Equol: History, chemistry, and formation. J. Nutr. 2010, 140, 1355s–1362s. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.; Clerici, C.; Lephart, E.D.; Cole, S.J.; Heenan, C.; Castellani, D.; Wolfe, B.E.; Nechemias-Zimmer, L.; Brown, N.M.; Lund, T.D.; et al. S-equol, a potent ligand for estrogen receptor beta, is the exclusive enantiomeric form of the soy isoflavone metabolite produced by human intestinal bacterial flora. Am. J. Clin. Nutr. 2005, 81, 1072–1079. [Google Scholar] [CrossRef] [Green Version]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference, Publication Year, Animal Species, Polyphenol(s), and Dosage | Major Physiological Issues Improved | Mode of Action | |
|---|---|---|---|
| Antioxidative and Anti-Inflammatory Action | Gut Microbiota Modulation | ||
| [12] 2013 Mice, polyphenol-rich pomegranate peel extract (PPE), p.o. in drinking water containing 0.2% PPE (average consumption of 6 mg/d per mouse) for 4 weeks | Reduced serum cholesterol (total and LDL) levels and alleviated tissue (colon and visceral adipose tissue) inflammation | − | Promoted the growth of gut bacteria, in particular, Bifidobacterium spp. |
| [13] 2014 Rats, p.o. as instant caffeinated coffee at a concentration of 20 g/L for 8 weeks (HFD was given for 10 weeks) | Reduced weight gain, adiposity, liver triglycerides, and energy intake | − | Decreased the Firmicutes/Bacteroidetes ratio |
| [14] 2015 Mice, high fat/high sucrose diet (HFHSD), polyphenol-rich cranberry extract, p.o. at 200 mg/kg/day for 8 weeks | Reduced visceral obesity and improved insulin sensitivity | Ameliorated oxidative stress and inflammation in the jejunum and reduced circulating LPS | Increased the relative abundance of Akkermansia spp. |
| [15] 2016 Mice, extractable polyphenol-rich fraction of table grapes (EP), p.o. with diet containing 1.1 g EP/kg for 16 weeks | Reduced white adipose tissue mass and improved glucose tolerance | − | Partially restored the HFD-mediated reduction in diversity |
| [16] 2016 Mice, green tea polyphenols (GTP), p.o. with a diet containing 0.05, 0.2, and 0.8% GTP for 8 weeks | Reduced obesity, and improved hepatic steatosis | − | Partially restored the HFD-mediated reduction in diversity |
| [17] 2017 Rats, a combination of quercetin (Q) and resveratrol (R), p.o. at 30 mg Q + 15 mg R/kg/day for 8 weeks | Reduced obesity | Attenuated serum inflammatory markers | Decreased the Firmicutes/Bacteroidetes ratio |
| [18] 2017 Mice, p.o. polyphenol- and caffeine-rich post-fermented Pu-erh tea, p.o. at 750 mg/kg/day for 12 weeks | Improved glucose and lipid metabolism disorder | Attenuated expression of inflammation genes in the proximal colon, reduced circulating LPS, and restored gut barrier integrity | Restored the HFD-induced gut microbial community structural shift |
| [19] 2018 Mice, polyphenol-rich cinnamon bark, or grape pomace extract (CBE or PBE), p.o. with a diet containing 0.2% CBE or and 0.8% PBE for 8 weeks | Reduced fat mass gain and adipose tissue inflammation, and ameliorated liver steatosis | Reduced adipose tissue inflammation, and improved gut barrier function | Decreased abundance of Desulfovibrio and Lactococcus at the genus level |
| [20] 2018 Mice, Lonicera caerulea L. berry polyphenols (LCBP), p.o. with diet containing 0.5% and 1% LCBP for 45 days | Improved hepatic steatosis | Attenuated serum inflammatory markers, and decreased LPS level in serum and liver | Decreased the Firmicutes/Bacteroidetes ratio |
| [21] 2019 Rats, resveratrol (RSV) and sinapic acid (SA), p.o. at 400 mg RSV/kg/day, 200 mg SA/kg/day, or a combination of RSV and SA for 8 weeks | Reduced fasting blood glucose levels and increased HDL-C levels by RSV | Decreased ROS and MDA levels in the colon, and increased total antioxidant capacity in the liver by SA | Combination of RSV and SA: Improved proportion of butyrate producer Blautia and Dorea from the Lachaospiraceae family and inhibited growth of bacterial species associated with diseases and inflammation, such as Bacteroides and Desulfovibrionaceae sp. |
| [22] 2019 Rats, sinapine (a rapeseed polyphenol), p.o. at 500 mg/kg/day for 12 weeks | Ameliorated NAFLD, reduced body weight and decreased TG and LDL-C levels. | Suppressed expression of NF-κB and TNF-α in the intestine and enhanced expression of IRS-1 in the adipose tissue | Decreased Firmicutes/Bacteroidetes ratio and increased abundance of probiotics, along with SCFA-mediated upregulation of G protein-coupled receptor 43 (GPR43) to inhibit the expression of inflammatory factors |
| [23] 2019 Mice, tea polyphenols (TPs) including EGCG, EGC, and ECG, p.o. at 100, 200, and 400 mg/kg/day for 12 weeks | Ameliorated hyperlipidemia, enhanced expression levels of hepatic lipid metabolism genes, and modulated gut microbiota | Maintenance of intestinal redox state by TPs | Decreased gut microbiota diversity and relative abundance of Proteobacteria, a source of LPS, possibly due to the antimicrobial activity of TPs |
| [24] 2019 Rats (treated with HFD + STZ), polyphenol-rich extracts from brown macroalga Lessonia trabeculata containing phlorotannin derivatives, phenolic acid derivatives, and gallocatechin derivatives, p.o. at 200 mg/kg/day for 4 weeks | Lowered fasting blood glucose and insulin levels, as well as better serum lipid profiles and antioxidant stress parameters | Increased response of antioxidant defense systems (e.g., CAT, SOD, and GSH in the liver) to oxidative stress | A positive effect on regulating the dysbiosis of the microbial ecology in diabetic rats |
| [25] 2019 Mice, pomegranate peel polyphenols including gallic acid, punicalagin, and catechin, p.o. at 150 and 300 mg/kg/day for 12 weeks | Alleviated obesity, decreased circulating proinflammatory cytokines, colonic tissue damage, and enhanced protein expression in the colonic tight junction | Improved oxidative damage and inflammation of the intestinal tissues, thereby reversing the reduced levels of tight junction proteins | Normalized the HFD-induced gut microbiota imbalance by increasing the abundance of beneficial bacteria in the colon |
| [26] 2020 Mice [fecal microbiota transplantation (FMT) to HFD-fed mice], resveratrol (RSV), p.o. at 300 mg/kg/day for 16 weeks followed by transplantation of the RSV-microbiota to HFD-fed mice (HFD-RSVT) to explore the function of the microbiota | HFD-RSVT decreased weight gain and increased insulin sensitivity | HFD-RSVT reduced the production of ROS and MDA in the intestine | A remarkable alteration in the composition of gut microbiota in mice treated with RSV, for example, enrichment of Bacteroides, Lachnospiraceae_NK4A136_ group, Blautia, Lachnoclostridium, Parabacteroides, and Ruminiclostridium_9, collectively referred to as RSV-microbiota |
| [27] 2020 Rats, Lonicera caerulea L. polyphenols containing anthocyanins, phenolic acids, and flavonoids, p.o. at 250 mg/kg/day for 8 weeks | Ameliorated intestinal permeability and intestinal inflammation; alleviated LPS-induced liver injury | Ameliorated intestinal oxidative stress damage (through regulation of the Nrf2/HO-1/NQO1 pathway) | Increased relative abundance of Bacteroidetes and Tenericutes and decreased relative abundance of Proteobacteria at the phylum level |
| [28] 2020 Mice (FMT from HFD-fed mice to HFD-fed mice), resveratrol (RSV), p.o. at 300 mg/kg/day for 16 weeks followed by transplantation of the HFDR-microbiota to HFD-fed mice (HFD-RSVT) | Alleviated NAFLD; ameliorated liver oxidative stress by HFD + RSV-microbiota treatment | HFD + RSV-microbiota treatment prevented HFD-induced production of ROS and improved antioxidant defense mechanisms (SOD and GSH levels) | The RSV-induced gut microbiota characterized by a decreased abundance of harmful bacteria, including Desulfovibrio, Lachnospiraceae_NK4A316_group, and Alistipes, as well as an increased abundance of SCFA-producing bacteria, such as Allobaculum, Bacteroides, and Blautia |
| [29] 2020 Mice, resveratrol (RSV), p.o. at 300 mg/kg/day for 16 weeks | Improved obesity | A two-part anti-obesity mechanism of RSV through the gut microbiota was proposed:(1) improved composition and function of the gut microbiota as well as the intestinal oxidative state; (2) 3-hydroxyphenylpropionic acid and 4-hydroxyphenylacetic acid (biotransformed from RSV by the gut microbiota), which may be responsible for the beneficial effects of RSV | |
| [30] 2020 Rats, polyphenol extracts from Shanxi-aged vinegar containing at least 41 polyphenols (including 18 phenolic acids), p.o. at 4, 8, and 16 mg/kg/day for 4 weeks | Improved hyperlipidemia | Improved inflammatory stress- and oxidative stress-related indicators | Decreased the Firmicutes/Bacteroidetes ratio; increased the diversity of microorganisms |
| [31] 2021 Mice, resveratrol (RSV) with probiotic Bifidobacteria, p.o. at 100 mg RSV/kg/day and probiotic Bifidobacteria for 3 weeks, starting the fifth week of HFD feeding | Coadministration of B. longum and RSV alleviated obesity and NAFLD | The combination of B. longum and RSV exerted an inhibitory effect on inflammatory cytokines and increased the levels of antioxidants, including SOD and GSH, and decreased the levels of MDA | RSV acted as an excellent prebiotic because most orally administered RSV is located in the bowel lumen |
| [32] 2021 Mice, Capsicum annuum L. ‘Senise’ extract (CAE) containing polyphenols, lycopene, and capsinoid derivatives, p.o. at 1, 10, and 25 mg/kg/day for 6 weeks | Promoted weight loss and improved plasma markers related to glucose and lipid metabolism | Reduced the expression of proinflammatory cytokines possibly due to the antioxidant property of CAE | Decreased the Firmicutes/Bacteroidetes ratio |
| [33] 2021 Rats, polyphenol-rich whole red grape juice, p.o. at 10 mL/day + physical training for 60 days | Lowered the concentration of IL-6 and TBARS | Reduced oxidative stress by activating the body’s antioxidant system, preventing the action of free radicals, and consequently, reducing the expression of inflammatory cytokines | The juice consumption beneficially modulated the gut microbiota |
| [34] 2021 Rats, Fu brick tea polyphenols, including EGCG, EGC, and ECG, p.o. at 100 mg/kg for 12 weeks | Improved the intestinal oxidative stress and intestinal barrier function, including intestinal inflammation and the integrity of the intestinal barrier | Attenuated HFD-induced gut microbiota dysbiosis, characterized by increased phylogenetic diversity and decreased Firmicutes/Bacteroidetes ratio | |
| Reference, Publication Year, Animal Species, Polyphenol(s), and Dosage | Major Physiological Issues Improved | Mode of Action | |
|---|---|---|---|
| Antioxidative and Anti-Inflammatory Action | Gut Microbiota Modulation | ||
| [56] 2017 Mice, chlorogenic acid (CA), p.o. with drinking water containing 1 mM CA for 15 days; 2.5% DSS was given during the last 8 days with CA | Ameliorated DSS-induced colitis and improved mucosal damage | Suppressed the active NF-κB signaling pathway in the colon | Decreased the Firmicutes/Bacteroidetes ratio and increased the relative abundance of Akkermansia |
| [57] 2017 Mice, nanoparticle curcumin (NC), p.o. with diet containing 0.2% NC for 18 days; 3% DSS was given from day 8 to day 18 | Ameliorated DSS-induced colitis and improved mucosal permeability | Suppressed NF-κB activation in colonic epithelial cells | Increased the abundance of butyrate-producing bacteria and fecal butyrate levels |
| [58] 2018 Mice, bronze tomato extract (BTE) rich in anthocyanins, flavonols, and stilbenoids, p.o. with diet containing 1% BTE for 2 weeks; 1% DSS was given from day 14 to day 29 | Ameliorated DSS-induced colitis | Suppressed LPS-mediated production of pro-inflammatory cytokines | Decreased the Firmicutes/Bacteroidetes ratio |
| [59] 2018 Rats, polyphenol-rich Chinese propolis (CP) or Brazilian propolis (BP), p.o. at 300 mg/kg/day for 17 days; 3% DSS was given from day 7 for 1 week | Ameliorated DSS-induced colitis | Alleviated the intestinal oxidative status, and suppressed inflammatory gene expression in the distal colon | Reduced populations of Bacteroides spp. |
| [60] 2018 Mice, quercetin aglycone (Q) or quercetin aglycone with monoglycosides (Q + MQ), p.o. with diet containing 0.21% Q or 0.35% Q + MQ for 7 days; 3% DSS was given from day 8 for 1 week | Ameliorated DSS-induced colitis | Suppressed oxidative stress indicated by MPO, GSH, and MDA | Ameliorated reduced Firmicutes population and increased Proteobacteria population by DSS |
| [61] 2019 Mice, rape bee pollen extract rich in polyphenols including kaempferol, sinapic acid, and rosmarinic acid, p.o. at 10.6 and 21.2 g/kg/day for 15 days; 3% DSS was given from day 8 to day 12 | Ameliorated DSS-induced colitis | Attenuated oxidative stress and downregulated the expression of inflammatory cytokines such as IL-1β | Reduced the abundance of Allobaculum and Bacteroides, and increased the abundance of Lactobacillus |
| [62] 2019 Rats, honey polyphenols including caffeic acid, chlorogenic acid, and rutin, p.o. at 10.5 mg/kg twice daily for 7 days; 3% DSS was given from day 0 to day 5 | Improved DSS-induced colonic apoptosis and reduced the expression of inflammatory cytokines in the colon | Improved the levels of SOD, GSH-Px, NO, and MPO; downregulated the expression of IL-1β, IL-6, TNF-α, and IFN-γ genes, and upregulated the expression of IκB-α gene | Reduced the population of Bacteroides, Corynebacterium, and Proteus species at the genus level |
| [63] 2021 Mice, taxifolin, p.o. at 100 mg/kg/day for 7 days; from day 8, mice received 3% DSS for 7 days | Prevented DSS-induced colitis | Inhibited the secretion of proinflammatory cytokines, and increased the secretion of IL-10, secretory IgA, SOD, and immunoglobulins; increased the expression of intestinal tight junction proteins | Restored the microbiota composition in the colon, including the decrease in the abundance of Bacteroidetes and the Bacteroidetes/Firmicutes ratio at the phylum level |
| [64] 2021 Mice, Exp. I, epigallocatechin-3-gallate (EGCG), p.o. or rectal administration at 50 mg/kg/day for 3 days starting after supplementation with 2.5% DSS for 7 days; Exp. II, EGCG, p.o. at 50 mg/kg/day for 3 weeks with 2.5% DSS for the last 6 days; Exp. III, 2.5% DSS for 7 days followed by FMT for 3 days | Exp. I: oral, but not rectal, EGCG alleviated DSS-induced colitis; Exp. II: oral EGCG prevented DSS-induced colitis; Exp. III: FMT alleviated DSS-induced colitis | Oral, but not rectal, EGCG attenuated oxidative stress, and exerted an anti-inflammatory effect along with enhanced integrity of the colonic barrier; oral EGCG pre-supplementation induced beneficial outcomes similar to those achieved with oral EGCG administration | Attenuation of colitis by oral EGCG suggests an intimate involvement of SCFA-producing bacteria, of the genus Akkermansia. |
| [65] 2021 Mice, green pea (Pisum sativum L.) hull polyphenol extracts containing quercetin and its derivatives, kaempferol trihexanside, and catechin and its derivatives, p.o. at 100 and 600 mg/kg for 14 days; 3% DSS was given for 7 weeks from day 8 | Alleviated DSS induced colitis | Restored oxidative balance, and regulated inflammatory factors along with repaired colonic function | Increased the Firmicutes/Bacteroidetes ratio, promoted the growth of Lactobacillaceae and Lachnospiraceae, and improved the level of SCFAs |
| Reference, Publication Year, Animal Model of the Disorder, Polyphenol(s), and Dosage | Major Physiological Issues Improved | Mode of Action | |
|---|---|---|---|
| Antioxidative and Anti-Inflammatory Action | Gut Microbiota Modulation | ||
| [66] 2019 Mice with fructose-induced NAFLD, loquat fruit extract rich in polyphenols including chlorogenic acid, cryptochlorogenic acid, and oleanolic acid, p.o. at 25 and 50 mg/kg/day for 8 weeks | Prevented fructose-induced NAFLD with mitigation of abnormal body weight and improved lipid metabolism | Mitigated oxidative stress and inflammation; reduced the endotoxin content and improved fructose-induced breakage of the intestinal barrier | Maintained normal Firmicutes/Bacteroidetes ratio |
| [67] 2020 Mice with western diet-induced NAFLD, vine tea polyphenol extracted from Ampelopsis grossedentata, p.o. in drinking water (0.5, 1, and 2%) for 12 weeks | Decreased the serum levels of cholesterol and triglycerides, and reduced the accumulation of hepatic lipid droplets | Activated Nrf2-mediated expression of hemeoxygenase-1 and quinone oxidoreductase, and reduced hepatic TBARS levels to prevent hepatic oxidative stress | Increased the relative abundance of Akkermansia, and reduced the Firmicutes/Bacteroidetes ratio |
| [68] 2021 Mice with alcohol-induced liver inflammation, Zhenjiang aromatic vinegar (a traditional fermented food in China) rich in polyphenols including catechin, p-hydroxybenzoic acid, and vanillic acid, p.o. at 200 and 800 mg/kg/day for 30 days; EtOH was given p.o. after 32 h, every day | Protected against alcohol-induced liver injury | Inhibited oxidative stress (reduced the levels of ROS, iNOS, MDA, 4-HNE, and 8-OHdG) and LPS-mediated inflammation | Modulated the composition of the gut microbiota and improved gut immunity and intestinal homeostasis; decreased the Firmicutes/Bacteroidetes ratio |
| [69] 2021 Mice with alcoholic liver disease, ellagic acid, p.o. at 50 and 100 mg/kg/day for 4 weeks to mice that received 15% alcohol | Alleviated alcohol-induced liver injury | Alleviated hepatic antioxidant activities (GSH-Px, CAT, MDA, SOD, and GSH), and proinflammatory cytokines levels (IL-6, IL-1β, and TNF-α) | Improved the alcohol-induced gut microbiota dysbiosis; restored the relative abundance of microbiota, such as Firmicutes, Verrucomicrobia, Actinobacteria, Bacteroidetes, and Proteobacteria at the phylum level |
| [70] 2021 Rats with LPS-induced liver disease, Aronia melanocarpa polyphenols containing anthocyanins, flavonols, and hydroxycinnamates, p.o. at 50, 100, and 200 mg/kg/day with LPS (p.o. at 200 μg/kg/day) for 4 weeks | Alleviated the degree of LPS-induced liver disease | Alleviated LPS-induced oxidative stress in the liver (reduced ROS and increased GSH levels). | Modulated the composition of the gut microbiota and improved the intestinal barrier function. At the phylum level, the enrichment of Verrucomicrobia microflora was alleviated and the abundance of Actinobacteria was decreased |
| [71] 2021 Mice with L-carnitine-induced liver injury, chlorogenic acid, p.o. at 200 and 400 mg/kg/day for 12 weeks with 3% L-carnitine in drinking water | Prevented L-carnitine-induced liver injury | Inhibited free radical production and improved the antioxidant defense system; inhibited the inflammatory reaction (i.e., IL-1, IL-6, TNF-α, and TNF-β levels). | Inhibited the L-carnitine-induced increase in the abundance of Firmicutes and Proteobacteria, and promoted Bacteroidetes at the phylum level |
| [72] 2021 Mice with obesity and hepatic steatosis induced by a western diet (WD), low in fiber but high in fats and sugars, p.o. as the WD supplemented with 1% grape polyphenols rich in B-type proanthocyanidins | Higher lean mass and lower fat mass, body weight, and hepatic steatosis | Reduced the intestinal oxidative stress | Increased the abundance of Akkermansia muciniphila, a gut microbe reported to increase energy expenditure |
| [73] 2018 Diabetic db/db mice, polyphenol-rich extract of Dendrobium loddigesi, p.o. at 50 and 100 mg/kg/day for 8 weeks | Decreased blood glucose, LDL-C, and body weight | Inhibited oxidative stress (reduced MDA and increased SOD, CAT, and GSH) in liver and kidney, attenuated serum inflammatory markers (IL-6 and TNF-α) | Decreased the Firmicutes/Bacteroidetes ratio |
| [74] 2019 Mice exposed to PM2.5 by instillation, hydroxytyrosol, p.o. at 50 mg/kg/day for 4 weeks | Alleviated PM2.5-induced visceral adiposity and insulin resistance | Inhibited PM2.5-induced oxidative stress-mediated activation of NF-κB | Enrichment of gut microbiota, and reduction of pathogenic bacteria |
| [75] 2020 Rats fed an obesogenic cafeteria diet, hesperidin (a citrus polyphenol), p.o. at 40 and 100 mg/kg/day for 8 weeks | Decreased the total cholesterol, LDL-C, and free fatty acids; ameliorated blood pressure and insulin sensitivity, and decreased the markers of arterial stiffness and inflammation | Metabolomics revealed an improvement in lower excretion of inflammation- and oxidative stress-related metabolites | Excretion of higher amounts of microbe-derived metabolites, which positively correlated with the Bacteroidaceae family |
| [76] 2021 Spontaneously hypertensive rats, Litchi chinensis seed extract, rich in polyphenols, including procyanidins, cinnamtannins, and rutin, p.o. at 30 and 60 mg/kg/day for 10 weeks | Reduced blood pressure and alleviated hypertension-induced renal damage | Attenuated oxidative stress and inflammation | Increased the relative abundance of Lactobacillus and production of SCFAs in the intestine |
| [77] 2021 Mice with doxorubicin (DOX)-induced heart failure, purified polyphenols from Arctium lappa L. including arctiin, dicaffeoyl succinoylquinic acid, and luteolin, p.o. at 50 and 150 mg/kg/day for 29 days; on day 22, 24, and 26, DOX was i.p. injected | Reduced heart failure syndrome and reduced serum activities of casein kinase and lactate dehydrogenase | Alleviated serum oxidative stress and reduced serum levels of inflammatory indices (TNF-α and NO). | Increased the abundance of Lactobacillaceae, Muribaculaceae, and Ruminococcaceae and decreased the abundance of Proteobacteria, Enterobacteria, and Escherichia_Shigella; enhanced the abundance of bacteria producing SCFAs |
| [78] 2021 Mice with potassium oxonate-induced hyperuricemia, Camellia japonica bee pollen extract containing polyphenols including kaempferol, quercetin, and gallic acid, p.o. at 2 and 4 g/kg/day for 3 weeks; starting on the 15th day, mice received potassium oxonate for the following 7 days | Reduced serum uric acid by inhibiting XOD activity and improved renal function | Increased antioxidant biomarkers, SOD activity, and GSH content, and decreased MDA content in the liver | Increased the abundance of Lactobacillus that has an anti-hyperuricemia effect; decreased the Firmicutes/Bacteroidetes ratio |
| Reference, Publication Year, Animal Species, Polyphenol(s), and Dosage | Observations | |
|---|---|---|
| Antioxidative and Anti-Inflammatory Action | Gut Microbiota Modulation | |
| [79] 2019 Rats, Jaboticaba (Plinia jaboticaba) peel extract containing gallic acid, flavonols, and anthocyanins, p.o. in drinking water (5 g/L) for 15 and 49 days | No disturbance of the antioxidant system | Increased the counts of Lactobacillus, Bifidobacterium, and Enterobacteriaceae |
| [80] 2020 Rats, freeze-dried whole golden kiwifruit (Actinidia chinensis) containing polyphenols, including quercetin, catechin, and chlorogenic acid, p.o. 4.6 g/kg/day for 28 days | Effectively improved the hepatic lipid profile, lipid peroxidation, long-chain fatty acid profile, and intestinal homeostasis | Reduced the Firmicutes/Bacteroidetes ratio at the phylum level; increased the abundance of beneficial bacteria (Lactobacillus and Barnesiella) and reduced the abundance of harmful bacteria (Enterococcus, Escherichia, and Staphylococcus) |
| [81] 2020 Mice, Penthorum chinense extract containing polyphenols, including pinocembrin-7-O-glucoside, thonningianin A, and brevifolin carboxylic acid, p.o. at 0.4 g/kg/day for 5 days | Increased antioxidant capacity (increased plasma SOD and hepatic CAT and GSH-px levels) | Increased microbiota diversity, elevated the Bacteroidetes/Firmicutes ratio at the phylum level, and enriched gut health-promoting bacteria |
| [82] 2020 Mice, anthocyanin-rich Lycium ruthenicum Murray (ACN), p.o. at 200 mg/kg/day for 12 weeks | Increased the antioxidant status in the liver (increased AOC, SOD, CAT, GSH, and GSH-Px levels and decreased the MDA levels); ameliorated the anti-inflammatory status in the colon (reduced the expression of iNOS, COX-2, TNF-α, IL-6, IL-1β, and IFN-γ mRNAs) and increased the intestinal barrier function (increased the expression of ZO-1, Occludin, Claudin-1, and Muc1 mRNAs) | Increased the proliferation of Barnesiella, Alistipes, Eisenbergiella, Coprobacter, and Odoribacter; increased SCFAs in the cecal content and feces. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niwano, Y.; Kohzaki, H.; Shirato, M.; Shishido, S.; Nakamura, K. Putative Mechanisms Underlying the Beneficial Effects of Polyphenols in Murine Models of Metabolic Disorders in Relation to Gut Microbiota. Curr. Issues Mol. Biol. 2022, 44, 1353-1375. https://doi.org/10.3390/cimb44030091
Niwano Y, Kohzaki H, Shirato M, Shishido S, Nakamura K. Putative Mechanisms Underlying the Beneficial Effects of Polyphenols in Murine Models of Metabolic Disorders in Relation to Gut Microbiota. Current Issues in Molecular Biology. 2022; 44(3):1353-1375. https://doi.org/10.3390/cimb44030091
Chicago/Turabian StyleNiwano, Yoshimi, Hidetsugu Kohzaki, Midori Shirato, Shunichi Shishido, and Keisuke Nakamura. 2022. "Putative Mechanisms Underlying the Beneficial Effects of Polyphenols in Murine Models of Metabolic Disorders in Relation to Gut Microbiota" Current Issues in Molecular Biology 44, no. 3: 1353-1375. https://doi.org/10.3390/cimb44030091