
Anticancer and Apoptotic Activity in Cervical Adenocarcinoma HeLa Using Crude Extract of Ganoderma applanatum

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Crude Extraction
2.2. Cell Culture
2.3. Solution Preparation
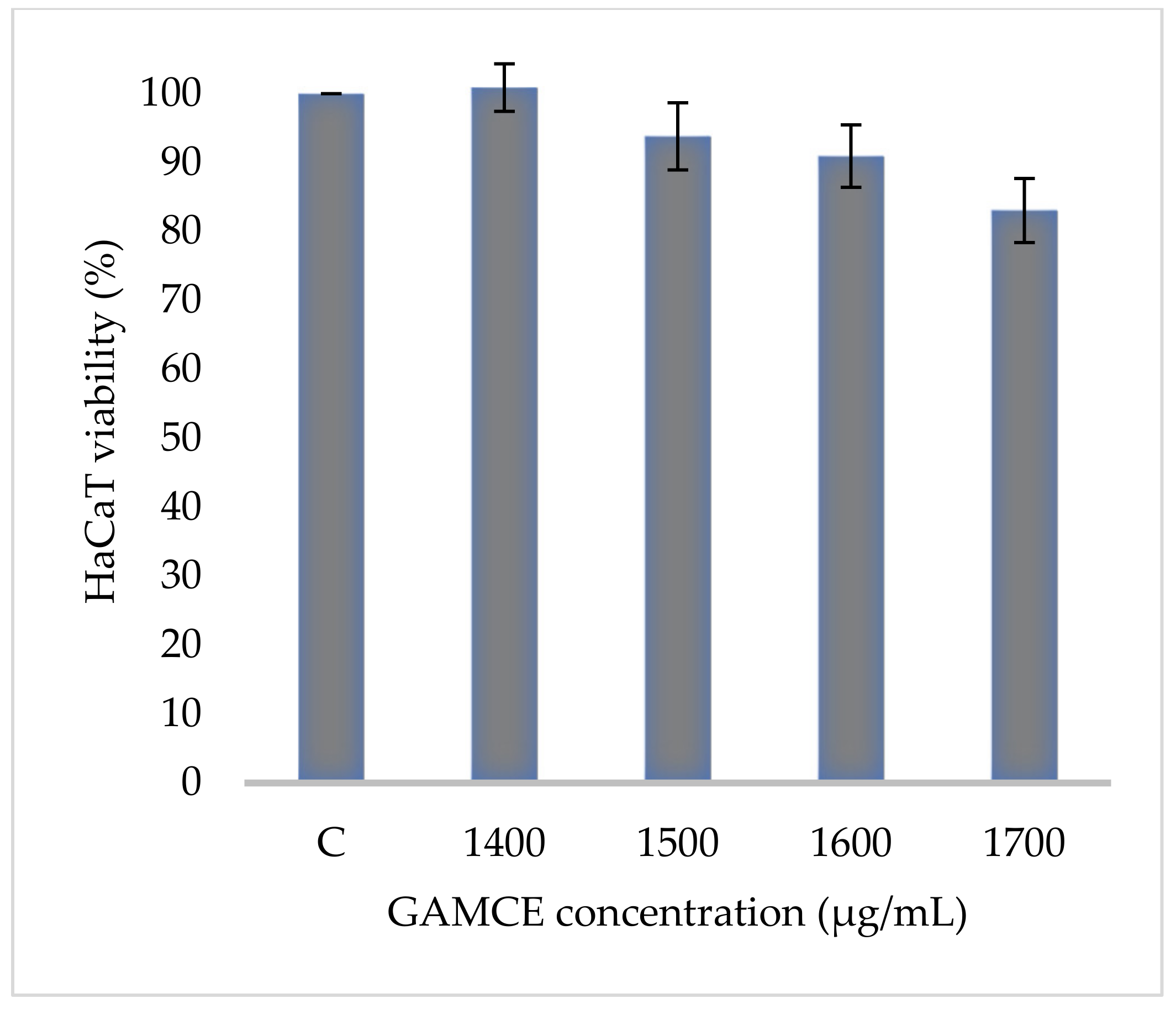
2.4. Cell Viability Assay
2.5. Protein Extraction and Westernblot
2.6. RNA Extraction
2.7. Reverse Transcriptase-PCR
2.8. Real Time-qPCR
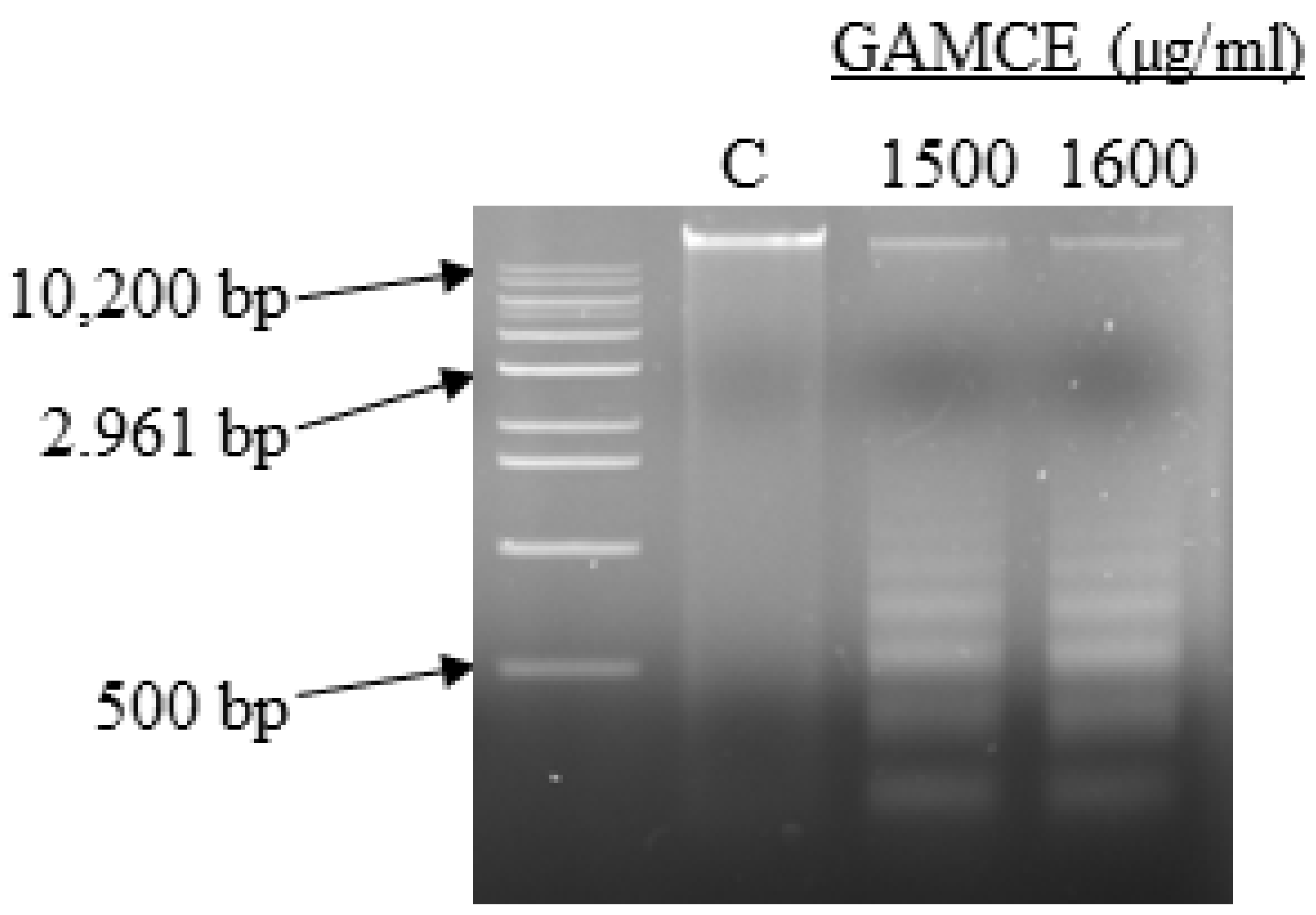
2.9. DNA Fragmentation
2.10. GC–MS
2.11. HPLC
2.12. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Apaf-1 | Apoptotic protease activating factor-1 |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| GAMCE | Crude extract of Ganoderma applanatum |
| GC-MS | Gas chromatography-mass spectrometry |
| HaCaT | Human keratinocyte |
| HeLa | Adenocarcinoma of the cervix |
| HPV | Human papilloma virus |
| IC50 | 50% inhibitory concentration |
| MEM | Minimum Essential Medium |
| PBS | Phosphate buffered saline |
| WHO | World Health Organization |
References
- Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 5 January 2022).
- Schiffman, M.; Castle, P.E.; Jeronimo, J.; Rodriguez, A.C.; Wacholder, S. Human papillomavirus and cervical cancer. Lancet 2007, 370, 890–907. [Google Scholar] [CrossRef]
- Vaccarella, S.; Lortet-Tieulent, J.; Plummer, M.; Franceschi, S.; Bray, F. Worldwide trends in cervical cancer incidence: Impact of screening against changes in disease risk factors. Eur. J. Cancer 2013, 49, 3262–3273. [Google Scholar] [CrossRef] [PubMed]
- Soutter, W.P.; Sasieni, P.; Panoskaltsis, T. Long-term risk of invasive cervical cancer after treatment of squamous cervical intraepithelial neoplasia. Int. J. Cancer 2006, 118, 2048–2055. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.A.; Jhingran, A.; Oaknin, A.; Denny, L. Cervical cancer. Lancet 2019, 393, 169–182. [Google Scholar] [CrossRef]
- Galvez-Olortegui, J.; Astigueta-Perez, J.; Abad-Licham, M.; Galvez-Olortegui, T.; Condor-Rojas, Y. Clinical practice guidelines in oncology. Gac. Mex. Oncol. 2016, 15, 103. [Google Scholar] [CrossRef] [Green Version]
- Small, W.; Bacon, M.A.; Bajaj, A.; Chuang, L.T.; Fisher, B.J.; Harkenrider, M.M.; Jhingran, A.; Kitchener, H.C.; Mileshkin, L.R.; Viswanathan, A.N.; et al. Cervical cancer: A global health crisis. Cancer 2017, 123, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- Strander, B. Cervical Cancer Prevention. Studies on Possible Improvements. Ph.D. Thesis, Göteborg University, Sahlgrenska Academy, Göteborg, Germany, 2008; pp. 1–118. [Google Scholar]
- Vu, M.; Yu, J.; Awolude, O.A.; Chuang, L. Cervical cancer worldwide. Curr. Probl. Cancer 2018, 42, 457–465. [Google Scholar] [CrossRef]
- Denny, L. Cervical cancer: Prevention and treatment. Discov. Med. 2012, 14, 125–131. [Google Scholar]
- Suphachai, C. Antioxidant and anticancer activities of Moringa oleifera leaves. J. Med. Plants Res. 2014, 8, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Kiddane, A.T.; Kim, G. Do Anticancer and Immunomodulatory Effects of Polysaccharides. Nutr. Cancer 2020, 73, 2219–2231. [Google Scholar] [CrossRef]
- Mingyi, Y.; Belwal, T.; Devkota, H.P.; Li, L.; Luo, Z. Trends of Utilizing Mushroom Polysaccharides (MPs) as Potent Nutraceutical Components in Food and Medicine: A Comprehensive Review. Trends Food Sci. Technol. 2019, 92, 94–110. [Google Scholar] [CrossRef]
- Souilem, F.; Fernandes, Â.; Calhelha, R.C.; Barreira, J.C.M.; Barros, L.; Skhiri, F.; Martins, A.; Ferreira, I.C.F.R. Wild mushrooms and their mycelia as sources of bioactive compounds: Antioxidant, anti-inflammatory and cytotoxic properties. Food Chem. 2017, 230, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, C. Reactive oxygen species and antioxidant properties from mushrooms. Synth. Syst. Biotechnol. 2017, 2, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Yang, X.H.; Yang, Z.L.; Tu, Z.C.; Cheng, Y.X. Miscellaneous meroterpenoids from Ganoderma applanatum. Tetrahedron 2016, 72, 4564–4574. [Google Scholar] [CrossRef]
- Ho, T.C.; Kim, M.H.; Cho, Y.J.; Park, J.S.; Nam, S.Y.; Chun, B.S. Gelatin-sodium alginate based films with Pseuderanthemum palatiferum (Nees) Radlk. freeze-dried powder obtained by subcritical water extraction. Food Packag. Shelf Life 2020, 24, 100469. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pérez, A.G.; Olías, R.; Sanz, C.; Olías, J.M. Furanones in Strawberries: Evolution during Ripening and Postharvest Shelf Life. J. Agric. Food Chem. 1996, 44, 3620–3624. [Google Scholar] [CrossRef]
- Shakeri, R.; Kheirollahi, A.; Davoodi, J. Apaf-1: Regulation and function in cell death. Biochimie 2017, 135, 111–125. [Google Scholar] [CrossRef]
- Albensi, B.C. What is nuclear factor kappa B (NF-κB) doing in and to the mitochondrion? Front. Cell Dev. Biol. 2019, 7, 154. [Google Scholar] [CrossRef]
- Ito, Y.; Kikuchi, E.; Tanaka, N.; Kosaka, T.; Suzuki, E.; Mizuno, R.; Shinojima, T.; Miyajima, A.; Umezawa, K.; Oya, M. Down-regulation of NF kappa B activation is an effective therapeutic modality in acquired platinum-resistant bladder cancer. BMC Cancer 2015, 15, 324. [Google Scholar] [CrossRef] [Green Version]
- Gilmore, T.D.; Herscovitch, M. Inhibitors of NF-κB signaling: 785 and counting. Oncogene 2006, 25, 6887–6899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, M.; Ben-Neriah, Y. Phosphorylation meets ubiquitination: The control of NF-κB activity. Annu. Rev. Immunol. 2000, 18, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Kanarek, N.; Ben-Neriah, Y. Regulation of NF-κB by ubiquitination and degradation of the IκBs. Immunol. Rev. 2012, 246, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Li, W.; Wang, T.; Shu, Y.; Liu, P. Paeoniflorin suppress NF-κB activation through modulation of IκBα and enhances 5-fluorouracil-induced apoptosis in human gastric carcinoma cells. Biomed. Pharmacother. 2008, 62, 659–666. [Google Scholar] [CrossRef]
- Yang, Y.; Kitagaki, J.; Dai, R.M.; Yien, C.T.; Lorick, K.L.; Ludwig, R.L.; Pierre, S.A.; Jensen, J.P.; Davydov, I.V.; Oberoi, P.; et al. Inhibitors of ubiquitin-activating enzyme (E1), a new class of potential cancer therapeutics. Cancer Res. 2007, 67, 9472–9481. [Google Scholar] [CrossRef] [Green Version]
- Khaleel, E.F.; Badi, R.M.; Satti, H.H.; Mostafa, D.G. Exendin-4 exhibits a tumour suppressor effect in SKOVR-3 and OVACR-3 ovarian cancer cells lines by the activation of SIRT1 and inhibition of NF-κB. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1092–1102. [Google Scholar] [CrossRef]
- Kumari, M.; Purohit, M.P.; Patnaik, S.; Shukla, Y.; Kumar, P.; Gupta, K.C. Curcumin loaded selenium nanoparticles synergize the anticancer potential of doxorubicin contained in self-assembled, cell receptor targeted nanoparticles. Eur. J. Pharm. Biopharm. 2018, 130, 185–199. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, S. Modulating Bcl-2 family proteins and caspase-3 in induction of apoptosis by paeoniflorin in human cervical cancer cells. Phyther. Res. 2011, 25, 1551–1557. [Google Scholar] [CrossRef]
- Bratton, S.B.; Salvesen, G.S. Regulation of the Apaf-1-caspase-9 apoptosome. J. Cell Sci. 2010, 123, 3209–3214. [Google Scholar] [CrossRef] [Green Version]
- Gogada, R.; Amadori, M.; Zhang, H.; Jones, A.; Verone, A.; Pitarresi, J.; Jandhyam, S.; Prabhu, V.; Black, J.D.; Chandra, D. Curcumin induces Apaf-1-dependent, p21-mediated caspase activation and apoptosis. Cell Cycle 2011, 10, 4128–4137. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, D.; Bishayee, K.; Ghosh, S.; Biswas, R.; Kumar Mandal, S.; Rahman Khuda-Bukhsh, A. [6]-Gingerol induces caspase 3 dependent apoptosis and autophagy in cancer cells: Drug-DNA interaction and expression of certain signal genes in HeLa cells. Eur. J. Pharmacol. 2012, 694, 20–29. [Google Scholar] [CrossRef]
- Karp, G. Cell signaling and signal transduction: Communication between cells. In Cell and Molecular Biology: Concepts and Experiments, 7th ed.; Wiley: Hoboken, NJ, USA, 2007; pp. 617–660. [Google Scholar]
- Husain, A.; Khan, S.A.; Iram, F.; Iqbal, M.A.; Asif, M. Insights into the chemistry and therapeutic potential of furanones: A versatile pharmacophore. Eur. J. Med. Chem. 2019, 171, 66–92. [Google Scholar] [CrossRef]
- Hosseinzadeh, Z.; Ramazani, A. An Overview of the Chemistry and Pharmacological Potentials of Furanones Skeletons. Curr. Org. Chem. 2019, 23, 1581–1599. [Google Scholar] [CrossRef]
- Yang, K.; Yang, J.Q.; Luo, S.H.; Mei, W.J.; Lin, J.Y.; Zhan, J.Q.; Wang, Z.Y. Synthesis of N-2(5H)-furanonyl sulfonyl hydrazone derivatives and their biological evaluation in vitro and in vivo activity against MCF-7 breast cancer cells. Bioorg. Chem. 2021, 107, 104518. [Google Scholar] [CrossRef]
- Kim, Y.; Nam, N.; You, Y.; Ahn, B. Synthesis and Cytotoxicity of 3,4-Diaryl-2(5H)-furanones. Bioorg. Med. Chem. Lett. 2002, 12, 719–722. [Google Scholar] [CrossRef]
- Guadalupe, A.; Pa, V.; Ram, T. Synthesis and cytotoxic evaluation of protoanemonin and three brominated derivatives. Rev. Colomb. De Quím. 2020, 49, 13–18. [Google Scholar]
- Byczek-Wyrostek, A.; Kitel, R.; Rumak, K.; Skonieczna, M.; Kasprzycka, A.; Walczak, K. Simple 2(5H)-furanone derivatives with selective cytotoxicity towards non-small cell lung cancer cell line A549—Synthesis, structure-activity relationship and biological evaluation. Eur. J. Med. Chem. 2018, 150, 687–697. [Google Scholar] [CrossRef]
- Wei, M.X.; Zhang, J.; Ma, F.L.; Li, M.; Yu, J.Y.; Luo, W.; Li, X.Q. Synthesis and biological activities of dithiocarbamates containing 2(5H)-furanone-piperazine. Eur. J. Med. Chem. 2018, 155, 165–170. [Google Scholar] [CrossRef]
- Bang, S.C.; Kim, Y.; Yun, M.Y.; Ahn, B.Z. 5-arylidene-2(5H)-furanone derivatives: Synthesis and structure-activity relationship for cytotoxicity. Arch. Pharm. Res. 2004, 27, 485–494. [Google Scholar] [CrossRef]
- Wu, Y.C.; Cao, L.; Mei, W.J.; Wu, H.Q.; Luo, S.H.; Zhan, H.Y.; Wang, Z.Y. Bis-2(5H)-furanone derivatives as new anticancer agents: Design, synthesis, biological evaluation, and mechanism studies. Chem. Biol. Drug Des. 2018, 92, 1232–1240. [Google Scholar] [CrossRef]
- Mancuso, R.; Ziccarelli, I.; Chimento, A.; Marino, N.; Della Ca’, N.; Sirianni, R.; Pezzi, V.; Gabriele, B. Catalytic Double Cyclization Process for Antitumor Agents against Breast Cancer Cell Lines. Food Sci. Hum. Wellness 2018, 3, 279–288. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Product Size | Sequences |
|---|---|---|
| NF-kB | 104 bp | Forward 5′-GGGAGATGAACCCTGCCAAA-3′ |
| Reverse 5′-AAAGTGGTGTTCCCAGGCAA-3′ | ||
| Caspase-3 | 102 bp | Forward 5′-CTGTGAACCCTGCATTTGGC-3′ |
| Reverse 5′-ACTTCGGAAGCTGAACCTGG-3′ | ||
| Cytochrome-c | 104 bp | Forward 5′-TGGCTTAATGTGTTCGCCCT-3′ |
| Reverse 5′-AAGCCCAAGCAAAGAGGGAA-3′ | ||
| Apaf-1 | 101 bp | Forward 5′-TGGGTGACTGACCTTTGCTTT-3′ |
| Reverse 5′-GTCTGTGAGGATTCCCCAGTG-3′ | ||
| Bax | 102 bp | Forward 5′-ACGAGGGTGATAGGTGGTACA-3′ |
| Reverse 5′-TGTTCTTCCCTTACCCACACG-3′ | ||
| Caspase-9 | 105 bp | Forward 5′-GAAGAGACCTGGCCAGAACC-3′ |
| Reverse 5′-ATTGCACAGCACGTTCACAC-3′ | ||
| Bak | 101 bp | Forward 5′-GGTTTTCCGCAGCTACGTTTT-3′ |
| Reverse 5′-GTTGCAGAGGTAAGGTGACCA-3′ | ||
| β-actin | 104 bp | Forward 5′-TCTTCCAGCCTTGCTTCCTG-3′ |
| Reverse 5′-GGTGTACAGGTCTTTGCGGA-3′ |
| Peak | Retention Time | Area% | Name |
|---|---|---|---|
| 1 | 3.171 | 6.77 | 2-Propanone, 1-hydroxy-(CAS) |
| 2 | 3.797 | 3.96 | Acetamide (CAS) |
| 3 | 4.422 | 10.25 | Dimethyl sulfoxide-D(6) |
| 4 | 5.372 | 1.43 | 2(5H)-furanone |
| 5 | 5.398 | 1.79 | 2(3H)-furanone, dihyfro-(CAS) |
| 6 | 5.510 | 3.56 | - |
| 7 | 5.589 | 5.65 | 2-Cyclopenten-1-one, 2-hydroxy- |
| 8 | 5.727 | 0.27 | 2(5H)-Furanone, 5-methyl- (identity?) (CAS) |
| 9 | 5.831 | 2.98 | 2-Furancarboxaldehyde, 5-methyl- (CAS) |
| 10 | 6.556 | 2.40 | 2-Hydroxy-gamma-butyrolactone |
| 11 | 6.580 | 1.18 | “5,6–dihydro–pyran–2,5–di–one” (so Pastorova) questiona |
| 12 | 7.078 | 2.07 | - |
| 13 | 7.333 | 1.94 | 4-Heptanone (CAS) |
| 14 | 7.602 | 2.53 | 2,5-Dimethyl-4-hydroxy-3(2H)-furanone |
| 15 | 8.220 | 1.66 | Anhydro-sugar |
| 16 | 8.474 | 1.61 | - |
| 17 | 9.032 | 4.01 | 4H-Pyran-4-one, 2,3-dihydro-3, 5-dihydroxy-6-methyl-(CAS) |
| 18 | 9.458 | 1.48 | 2-Butenethioic acid, 3-(ethylthio)-, S-(1-methylethyl) ester |
| 19 | 9.814 | 0.95 | 1,2-Benzenediol (CAS) |
| 20 | 10.128 | 3.48 | 1,4:3,6-Dianhydro-.alpha.-d-glucopyranose |
| 21 | 10.262 | 4.96 | 5-Hydroxymethylfurfural |
| 22 | 12.725 | 3.37 | - |
| 23 | 14.561 | 2.36 | - |
| 24 | 17.089 | 23.94 | DL-Arabinitol |
| 25 | 17.941 | 5.41 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiddane, A.T.; Kang, M.-J.; Ho, T.C.; Getachew, A.T.; Patil, M.P.; Chun, B.-S.; Kim, G.-D. Anticancer and Apoptotic Activity in Cervical Adenocarcinoma HeLa Using Crude Extract of Ganoderma applanatum. Curr. Issues Mol. Biol. 2022, 44, 1012-1026. https://doi.org/10.3390/cimb44030067
Kiddane AT, Kang M-J, Ho TC, Getachew AT, Patil MP, Chun B-S, Kim G-D. Anticancer and Apoptotic Activity in Cervical Adenocarcinoma HeLa Using Crude Extract of Ganoderma applanatum. Current Issues in Molecular Biology. 2022; 44(3):1012-1026. https://doi.org/10.3390/cimb44030067
Chicago/Turabian StyleKiddane, Anley Teferra, Min-Jae Kang, Truc Cong Ho, Adane Tilahun Getachew, Maheshkumar Prakash Patil, Byung-Soo Chun, and Gun-Do Kim. 2022. "Anticancer and Apoptotic Activity in Cervical Adenocarcinoma HeLa Using Crude Extract of Ganoderma applanatum" Current Issues in Molecular Biology 44, no. 3: 1012-1026. https://doi.org/10.3390/cimb44030067