
Multiplex Detection of Pleurotus ostreatus Mycoviruses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Primer Design
2.3. RNA Extraction
2.4. Reverse Transcription
2.5. PCR Amplification
2.6. Cloning and Sequencing
3. Results
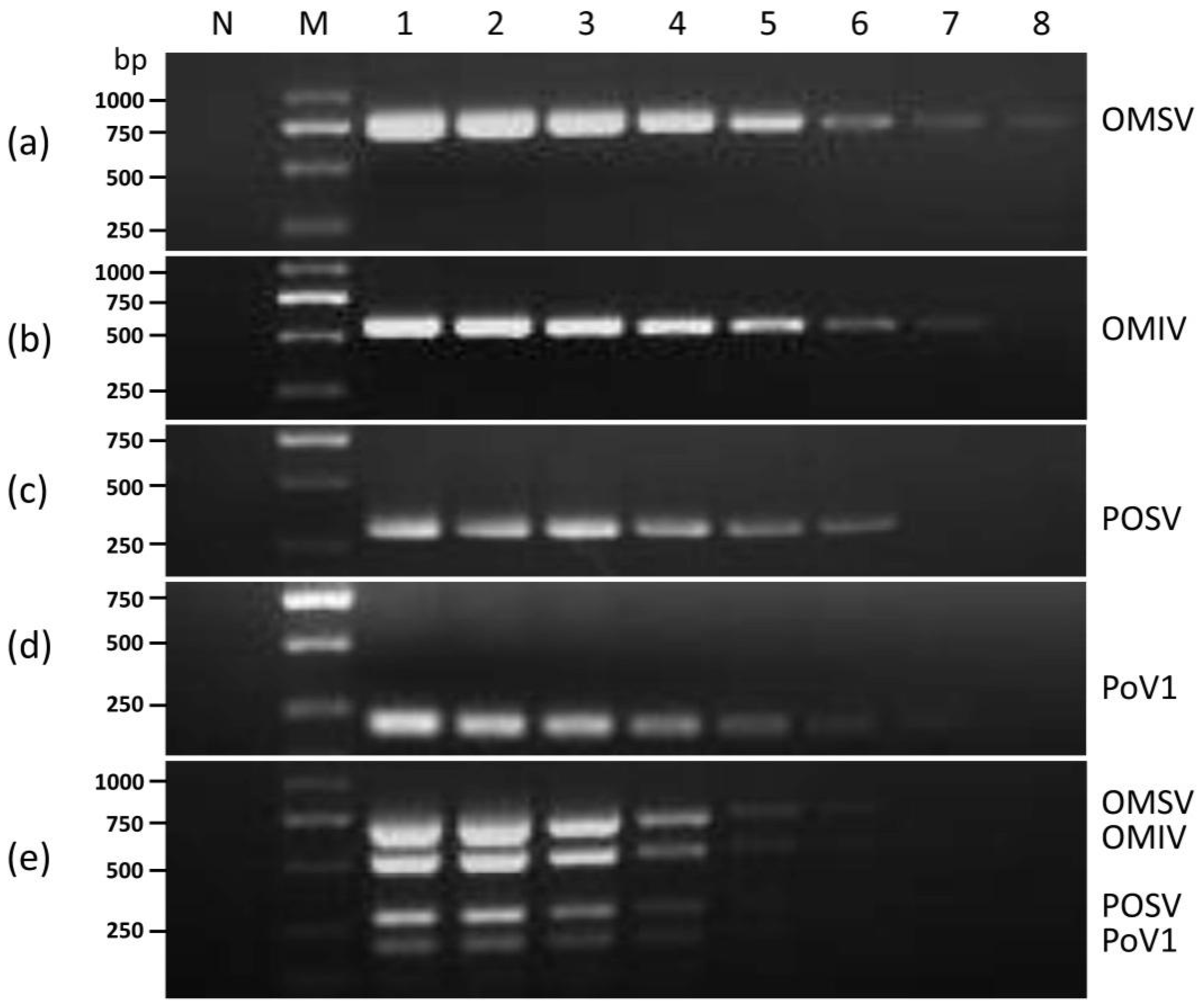
3.1. Establishment of the Multiplex RT-PCR Assay
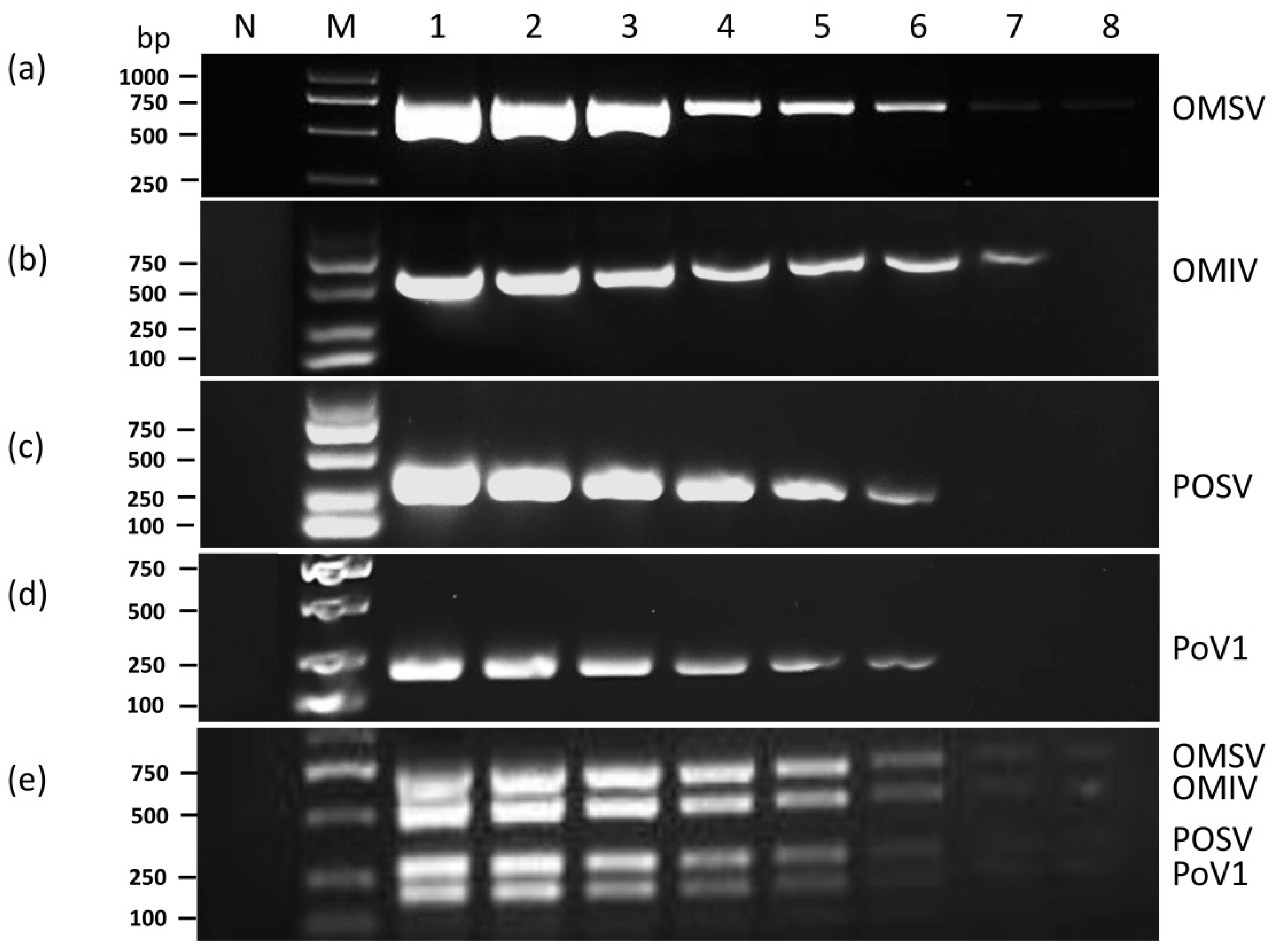
3.2. Sensitivity of Multiplex RT-PCR
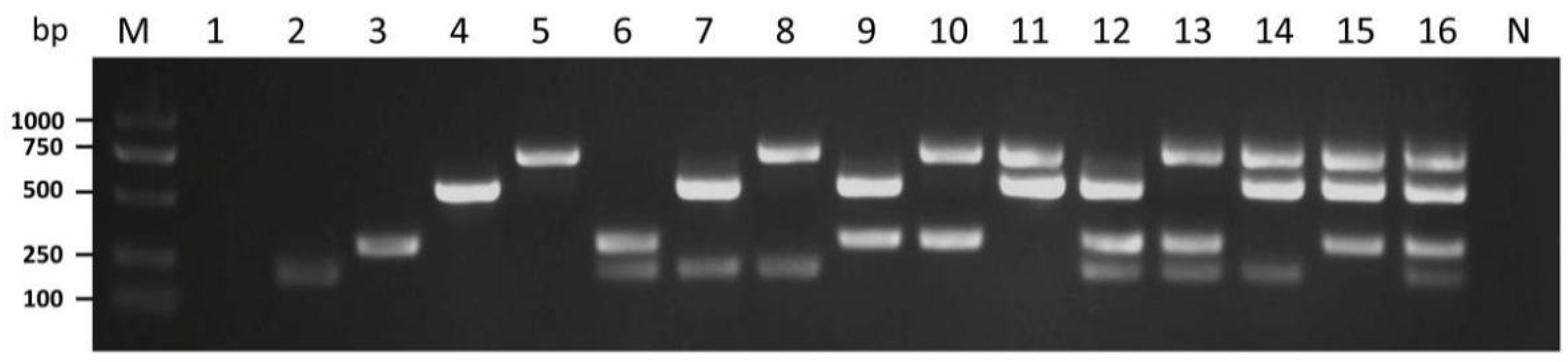
3.3. Specificity of Multiplex RT-PCR
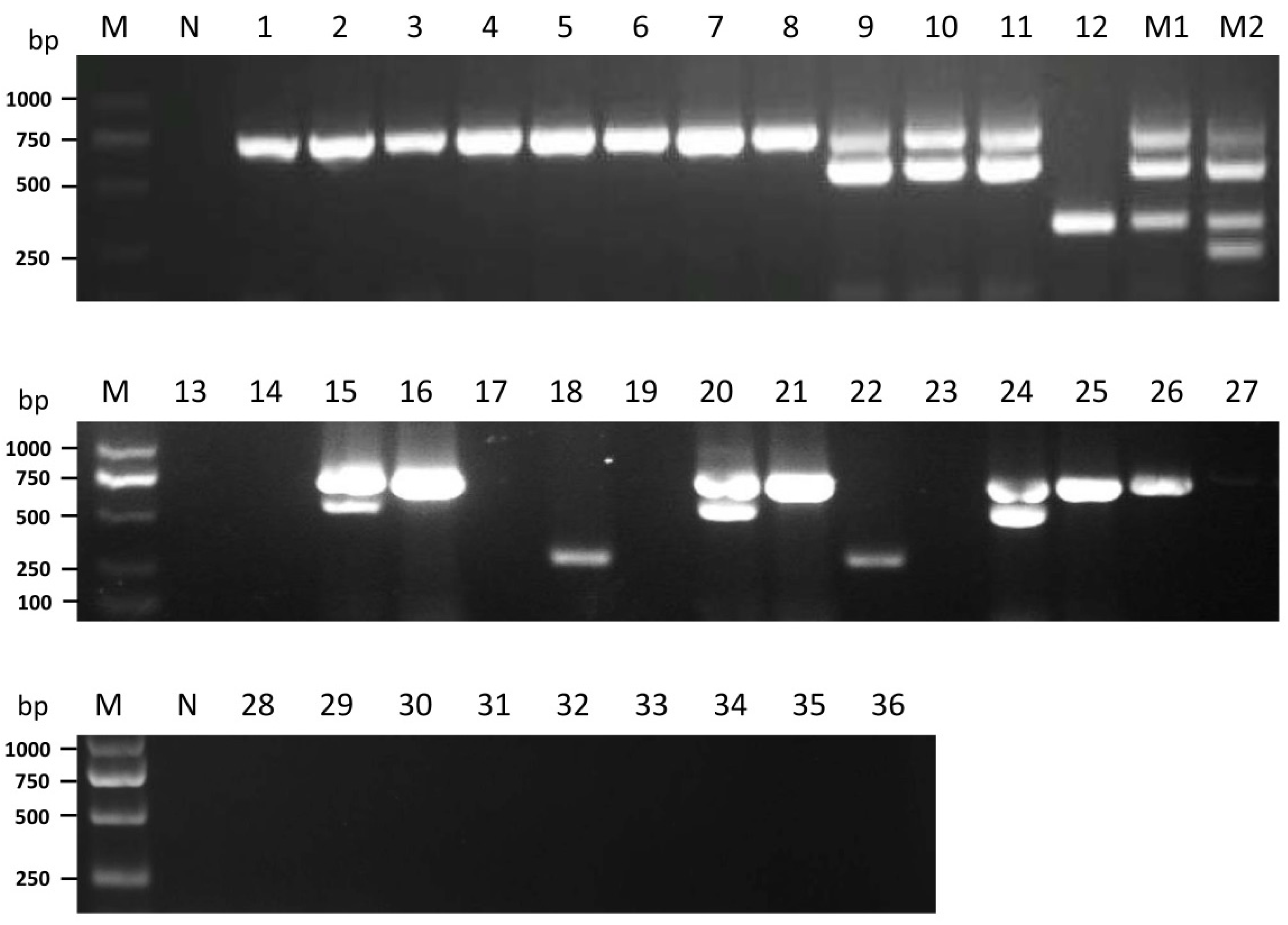
3.4. Detection of the Four Mycoviruses in Different P. ostreatus Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doedt, T.; Krishnamurthy, S.; Bockmuhl, D.P.; Tebarth, B.; Stempel, C.; Russell, C.L.; Brown, A.J.; Ernst, J.F. APSES proteins regulate morphogenesis and metabolism in candida albicans. Mol. Biol. Cell 2004, 15, 3167–3180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollings, M.; Stone, O.M. Viruses that infect fungi. Phytopathology 1971, 9, 93–118. [Google Scholar] [CrossRef]
- Sahin, E.; Akata, I. Viruses infecting macrofungi. Virusdisease 2018, 29, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; Gao, J.; Li, Y. Diversity of mycoviruses in edible fungi. Virus Genes 2022, 58, 377–391. [Google Scholar] [CrossRef]
- Rumbou, A.; Vainio, E.J.; Buttner, C. Towards the forest virome: High-throughput sequencing drastically expands our understanding on virosphere in temperate forest ecosystems. Microorganisms 2021, 9, 1730. [Google Scholar] [CrossRef]
- Hollings, M. Viruses associated with a die-back disease of cultivated mushroom. Nature 1962, 196, 962–965. [Google Scholar] [CrossRef]
- Linnakoski, R.; Sutela, S.; Coetzee, M.P.A.; Duong, T.A.; Pavlov, I.N.; Litovka, Y.A.; Hantula, J.; Wingfield, B.D.; Vainio, E.J. Armillaria root rot fungi host single-stranded RNA viruses. Sci. Rep. 2021, 11, 7336. [Google Scholar] [CrossRef]
- Guo, M.P.; Shen, G.Y.; Wang, J.J.; Liu, M.J.; Bian, Y.B.; Xu, Z.Y. Mycoviral diversity and characteristics of a negative-stranded RNA virus LeNSRV1 in the edible mushroom Lentinula edodes. Virology 2021, 555, 89–101. [Google Scholar] [CrossRef]
- Lin, Y.H.; Fujita, M.; Chiba, S.; Hyodo, K.; Andika, I.B.; Suzuki, N.; Kondo, H. Two novel fungal negative-strand RNA viruses related to mymonaviruses and phenuiviruses in the shiitake mushroom (Lentinula edodes). Virology 2019, 533, 125–136. [Google Scholar] [CrossRef]
- Komatsu, A.; Kondo, H.; Sato, M.; Kurahashi, A.; Nishibori, K.; Suzuki, N.; Fujimori, F.J.M. Isolation and characterization of a novel mycovirus infecting an edible mushroom, Grifola frondosa. Mycoscience 2019, 60, 211–220. [Google Scholar] [CrossRef]
- Qiu, L.Y.; Li, Y.P.; Liu, Y.M.; Gao, Y.Q.; Qi, Y.C.; Shen, J.W. Particle and naked RNA mycoviruses in industrially cultivated mushroom Pleurotus ostreatus in China. Fungal Biol. 2010, 114, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Magae, Y.; Sunagawa, M. Characterization of a mycovirus associated with the brown discoloration of edible mushroom, Flammulina velutipes. Virol. J. 2010, 7, 342. [Google Scholar] [CrossRef] [Green Version]
- Ro, H.S.; Kang, E.J.; Yu, J.S.; Lee, T.S.; Lee, C.W.; Lee, H.S. Isolation and characterization of a novel mycovirus, PeSV, in Pleurotus eryngii and the development of a diagnostic system for it. Biotechnol. Lett. 2007, 29, 129–135. [Google Scholar] [CrossRef]
- Ro, H.S.; Lee, N.J.; Lee, C.W.; Lee, H.S. Isolation of a novel mycovirus OMIV in Pleurotus ostreatus and its detection using a triple antibody sandwich-ELISA. J. Virol. Methods 2006, 138, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Revill, P.A.; Davidson, A.D.; Wright, P.J. The nucleotide sequence and genome organization of mushroom bacilliform virus: A single-stranded RNA virus of Agaricus bisporus (Lange) Imbach. Virology 1994, 202, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Liang, P.; Yu, M.; Chang, S.T. A new double-stranded RNA virus from Volvariella volvacea. Mycologia 1988, 80, 849–853. [Google Scholar] [CrossRef]
- Barroso, G.; Labarère, J. Evidence for viral and naked double-stranded RNAs in the basidiomycete Agrocybe aegerita. Curr. Genet. 1990, 18, 231–237. [Google Scholar] [CrossRef]
- Huttinga, H.; Wichers, H.J.; Zaayen, D.V. Filamentous and polyhedral virus-like particles in Boletus edulis. Neth. J. Plant Pathol. 1975, 81, 102–106. [Google Scholar] [CrossRef]
- Sahin, E.; Keskin, E.; Akata, I. Molecular characterization of the complete genome of a novel partitivirus hosted by the saprobic mushroom Leucocybe candicans. Arch. Microbiol. 2021, 203, 5825–5830. [Google Scholar] [CrossRef]
- Li, X.F.; Xie, J.T.; Hai, D.; Sui, K.P.; Yin, W.Q.; Sossah, F.L.; Jiang, D.H.; Song, B.; Li, Y. Molecular characteristics of a novel ssRNA virus isolated from Auricularia heimuer in China. Arch. Virol. 2020, 165, 1495–1499. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Keskin, E.; Akata, I. Novel and diverse mycoviruses co-inhabiting the hypogeous ectomycorrhizal fungus Picoa juniperi. Virology 2021, 552, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Sutela, S. Mixed infection by a partitivirus and a negative-sense RNA virus related to mymonaviruses in the polypore fungus Bondarzewia berkeleyi. Virus Res. 2020, 286, 198079. [Google Scholar] [CrossRef]
- Abdoulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses infecting the plant pathogenic fungus Rhizoctonia solani. Viruses 2019, 11, 1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.Y.; Kim, N.; Kim, D.H.; Kim, J.M. The PoV mycovirus affects extracellular enzyme expression and fruiting body yield in the oyster mushroom, Pleurotus ostreatus. Sci. Rep. 2020, 10, 1094. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.P.; Bian, Y.B.; Wang, J.J.; Wang, G.Z.; Ma, X.L.; Xu, Z.Y. Biological and molecular characteristics of a novel partitivirus infecting the edible fungus Lentinula edodes. Plant Dis. 2017, 101, 726–733. [Google Scholar] [CrossRef] [Green Version]
- Eastwood, D.; Green, J.; Grogan, H.; Burton, K. Viral agents causing brown cap mushroom disease of Agaricus bisporus. Appl. Environ. Microb. 2015, 81, 7125–7134. [Google Scholar] [CrossRef] [Green Version]
- Kwon, Y.C.; Jeong, D.W.; Gim, S.I.; Ro, H.S.; Lee, H.S. Curing viruses in Pleurotus ostreatus by growth on a limited nutrient medium containing cAMP and rifamycin. J. Virol. Methods 2012, 185, 156–159. [Google Scholar] [CrossRef]
- Lim, W.S.; Jeong, J.H.; Jeong, R.D.; Yoo, Y.B.; Yie, S.W.; Kim, K.H. Complete nucleotide sequence and genome organization of a dsRNA partitivirus infecting Pleurotus ostreatus. Virus Res. 2005, 108, 111–119. [Google Scholar] [CrossRef]
- Yu, H.J.; Lim, D.; Lee, H.-S. Characterization of a novel single-stranded RNA mycovirus in Pleurotus ostreatus. Virology 2003, 314, 9–15. [Google Scholar] [CrossRef]
- Hu, H.J.; Wang, J.R.; Cheng, X.H.; Liu, Y.; Zhang, X.Y. Preliminary studies on the effects of oyster mushroom spherical virus China strain on the mycelial growth and fruiting body yield of the edible mushroom Pleurotus ostreatus. Biology 2022, 11, 574. [Google Scholar] [CrossRef] [PubMed]
- Baldauf, S.L. Phylogeny for the faint of heart: A tutorial. Trends Genet. 2003, 19, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.T.; Jiang, D.H. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Won, H.K.; Park, S.J.; Kim, D.K.; Shin, M.J.; Kim, N.; Lee, S.H.; Kwon, Y.C.; Ko, H.K.; Ro, H.S.; Lee, H.S. Isolation and characterization of a mycovirus in Lentinula edodes. J. Microbiol. 2013, 51, 118–122. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, E.; Doyle, S.; Amini, A.; Grogan, H.; Fitzpatrick, D.A. Transmission of mushroom virus X and the impact of virus infection on the transcriptomes and proteomes of different strains of Agaricus bisporus. Fungal Biol. 2021, 125, 704–717. [Google Scholar] [CrossRef]
- Song, H.Y.; Choi, H.J.; Jeong, H.; Choi, D.; Kim, D.H.; Kim, J.M. Viral effects of a dsRNA mycovirus (PoV-ASI2792) on the vegetative growth of the edible mushroom Pleurotus ostreatus. Mycobiology 2016, 44, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.J.; Guo, M.P.; Wang, J.J.; Bian, Y.B.; Xu, Z.Y. Curing two predominant viruses occurring in Lentinula edodes by chemotherapy and mycelial fragmentation methods. J. Virol. Methods 2022, 300, 114370. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, M.G.; Kim, J.; Lee, H.S.; Ro, H.S. Detection of the mycovirus OMSV in the edible mushroom, Pleurotus ostreatus, using an SPR biosensor chip. J. Virol. Methods 2008, 148, 120–124. [Google Scholar] [CrossRef]
- Ozkan-Kotiloglu, S.; Coutts, R.H.A. Multiplex detection of Aspergillus fumigatus Mycoviruses. Viruses 2018, 10, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Cheng, J.L.; Huang, T.; Zheng, X.; Wu, Y.F. A multiplex reverse transcription PCR assay for simultaneous detection of five tobacco viruses in tobacco plants. J. Virol. Methods 2012, 183, 57–62. [Google Scholar] [CrossRef]
- Sun, Y.J.; Guo, M.P.; Wang, J.J.; Bian, Y.B.; Xu, Z.Y. Development of multiplex RT-PCR for the detection of two main mycoviruses infecting Chinese Lentinula edodes germplasm resource. Microbiol. China 2019, 46, 1381–1389. (In Chinese) [Google Scholar]
- Shahi, S.; Chiba, S.; Kondo, H.; Suzuki, N. Cryphonectria nitschkei chrysovirus 1 with unique molecular features and a very narrow host range. Virology 2021, 554, 55–65. [Google Scholar] [CrossRef]
- Cornejo, C.; Hisano, S.; Bragança, H.; Suzuki, N.; Rigling, D. A new double-stranded RNA mycovirus in Cryphonectria naterciae is able to cross the species barrier and is deleterious to a new host. J. Fungi 2021, 7, 861. [Google Scholar] [CrossRef]
- Liu, Y.C.; Linder-Basso, D.; Hillman, B.I.; Kaneko, S.; Milgroom, M.G. Evidence for interspecies transmission of viruses in natural populations of filamentous fungi in the genus Cryphonectria. Mol. Ecol. 2003, 12, 1619–1628. [Google Scholar] [CrossRef]
- Zhu, J.Z.; Zhu, H.J.; Gao, B.D.; Zhou, Q.; Zhong, J. Diverse, novel mycoviruses from the virome of a hypovirulent Sclerotium rolfsii strain. Front. Plant Sci. 2018, 9, 1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deakin, G.; Dobbs, E.; Bennett, J.M.; Jones, I.M.; Grogan, H.M.; Burton, K.S. Multiple viral infections in Agaricus bisporus—Characterisation of 18 unique RNA viruses and 8 ORFans identified by deep sequencing. Sci. Rep. 2017, 7, 2469. [Google Scholar] [CrossRef] [Green Version]
- Xue, B.; Shang, J.; Yang, J.; Zhang, L.; Du, J.B.; Yu, L.; Yang, W.Y.; Naeem, M. Development of a multiplex RT-PCR assay for the detection of soybean mosaic virus, bean common mosaic virus and cucumber mosaic virus in field samples of soybean. J. Virol. Methods 2021, 298, 114278. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.C.; Xie, A.T.; Wang, X.L.; Wang, S.J.; Liu, X.D.; Zhou, T. Detection of oyster mushroom spherical virus in Beijing and its controlling measures. China Plant Prot. 2016, 36, 9–11. (In Chinese) [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ to 3′) | Target Virus a | Amplicon Size (bp) |
|---|---|---|---|
| OMSV-F | ACCCCCCCAGGATCTCAAGCTTC | OMSV | 672 |
| OMSV-R | GAGATGTAGACRTTGAAAGC | ||
| OMIV-F | AACATTGTTGATCACGCTCT | OMIV | 540 |
| OMIV-R | GGCTTCAGAATAAAGATTGT | ||
| POSV-F | ATCWCATGGCTATCAACCTA | POSV | 310 |
| POSV-R | AGCTGAATTATCGTCACCCA | ||
| PoV1-F | AAACTCGAAGAGTTCCTTTC | PoV1 | 200 |
| PoV1-R | GCGCGTGGGCCACGTTCGGG |
| Number | Strain Name | Source | Presence of Virus | |||
|---|---|---|---|---|---|---|
| OMSV | OMIV | POSV | PoV1 | |||
| 1 | 8129 | Yantai | + | − | − | − |
| 2 | DF5 | Yangzhou | + | − | − | − |
| 3 | DF5-2 | Liaocheng | + | − | − | − |
| 4 | 969 | Liaocheng | + | − | − | − |
| 5 | Kang-2 | Liaocheng | + | − | − | − |
| 6 | Kang-3 | Liaocheng | + | − | − | − |
| 7 | P89 | Beijing | + | − | − | − |
| 8 | P99 | Beijing | + | − | − | − |
| 9 | PG-0122-1 | Yantai | + | + | − | − |
| 10 | Heiping | Yantai | + | + | − | − |
| 11 | PG-ZP20 | Yantai | + | + | − | − |
| 12 | TD300 | Linyi | − | − | + | − |
| 13 | PG-2203 | Jinan | − | − | − | − |
| 14 | PG-2204 | Jinan | − | − | − | − |
| 15 | P2108 | Dezhou | + | + | − | − |
| 16 | LD-0701 | Weifang | + | − | − | − |
| 17 | LD-0704 | Weifang | − | − | − | − |
| 18 | LD-0707 | Weifang | − | − | + | − |
| 19 | LD-0719 | Weifang | − | − | − | − |
| 20 | LD-1011 | Qingdao | + | + | − | − |
| 21 | LD-1015 | Qingdao | + | − | − | − |
| 22 | PGH-1011 | Zibo | − | − | + | − |
| 23 | PGH-1012 | Zibo | − | − | − | − |
| 24 | PGH-1014 | Zibo | + | + | − | − |
| 25 | PGZ-1020 | Weihai | + | − | − | − |
| 26 | PGZ-1021 | Weihai | + | − | − | − |
| 27 | PGZ-1022 | Weihai | − | − | − | − |
| 28 | Huimei | Liaocheng | − | − | − | − |
| 29 | Xianfeng-1 | Liaocheng | − | − | − | − |
| 30 | PG-0324 | Yantai | − | − | − | − |
| 31 | WPG-1107 | Yantai | − | − | − | − |
| 32 | 8105 | Yangzhou | − | − | − | − |
| 33 | Luping-0417-5 | Yantai | − | − | − | − |
| 34 | F803 | Linyi | − | − | − | − |
| 35 | PG-ZP17 | Yantai | − | − | − | − |
| 36 | LD-0328 | Yantai | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Hu, H.; Wang, Y.; Yan, J.; Liu, Y.; Wang, J.; Cheng, X. Multiplex Detection of Pleurotus ostreatus Mycoviruses. Curr. Issues Mol. Biol. 2022, 44, 5778-5787. https://doi.org/10.3390/cimb44110392
Zhang X, Hu H, Wang Y, Yan J, Liu Y, Wang J, Cheng X. Multiplex Detection of Pleurotus ostreatus Mycoviruses. Current Issues in Molecular Biology. 2022; 44(11):5778-5787. https://doi.org/10.3390/cimb44110392
Chicago/Turabian StyleZhang, Xiaoyan, Haijing Hu, Yifan Wang, Junjie Yan, Yu Liu, Jianrui Wang, and Xianhao Cheng. 2022. "Multiplex Detection of Pleurotus ostreatus Mycoviruses" Current Issues in Molecular Biology 44, no. 11: 5778-5787. https://doi.org/10.3390/cimb44110392