
Protective Effects of One 2,4-Dihydro-3H-Pyrazol-3-one Derivative against Posterior Capsular Opacification by Regulation of TGF-β2/SMADs and Non-SMAD Signaling, Collagen I, and Fibronectin Proteins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
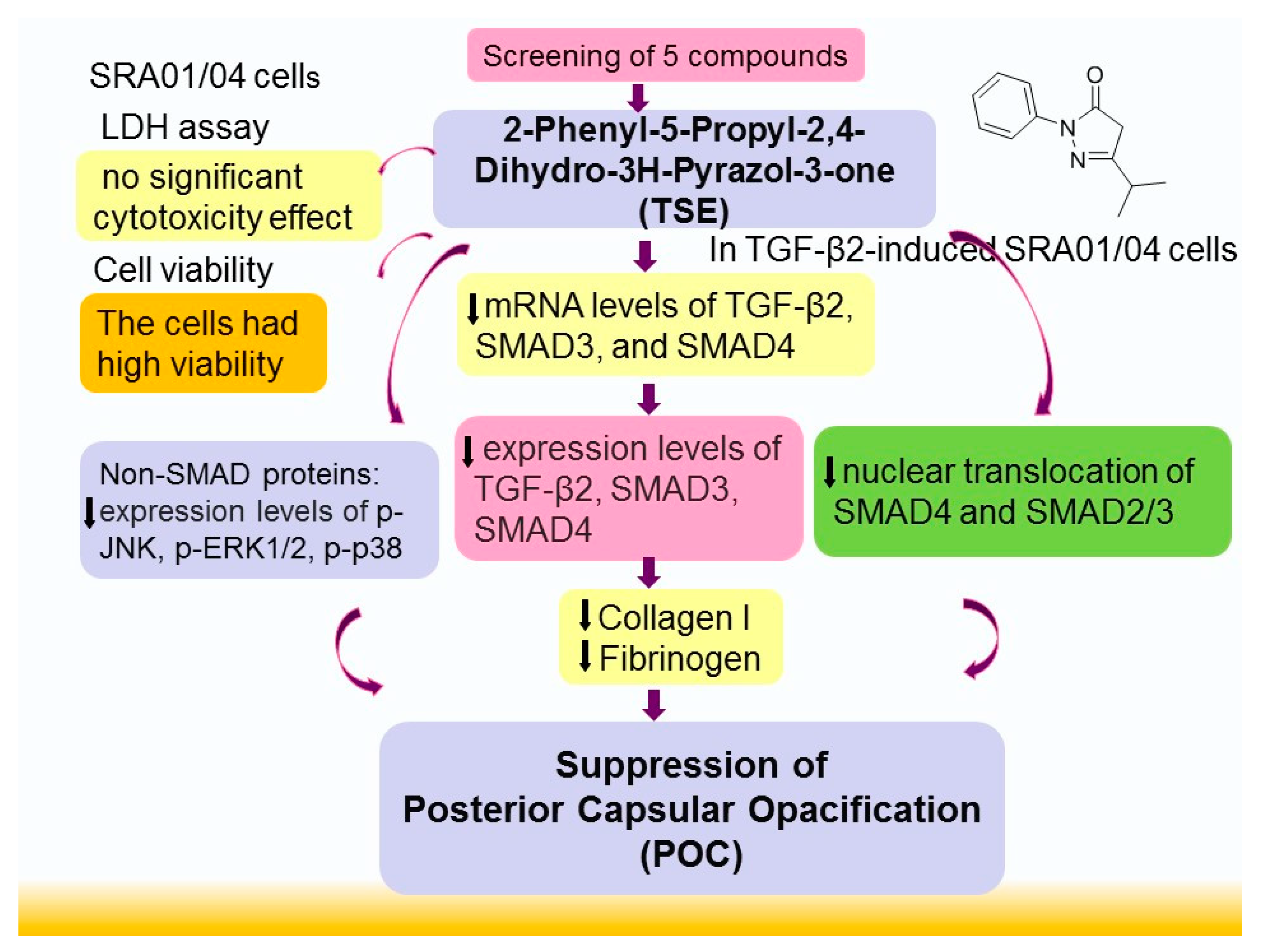
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Lines
2.3. MTT Assay
2.4. Lactate Dehydrogenase (LDH) Activity Assay
2.5. Immunoblotting Analysis
2.6. Confocal Immunofluorescence Microscopy
2.7. Relative Quantification of mRNA
2.8. Statistical Analyses
3. Results
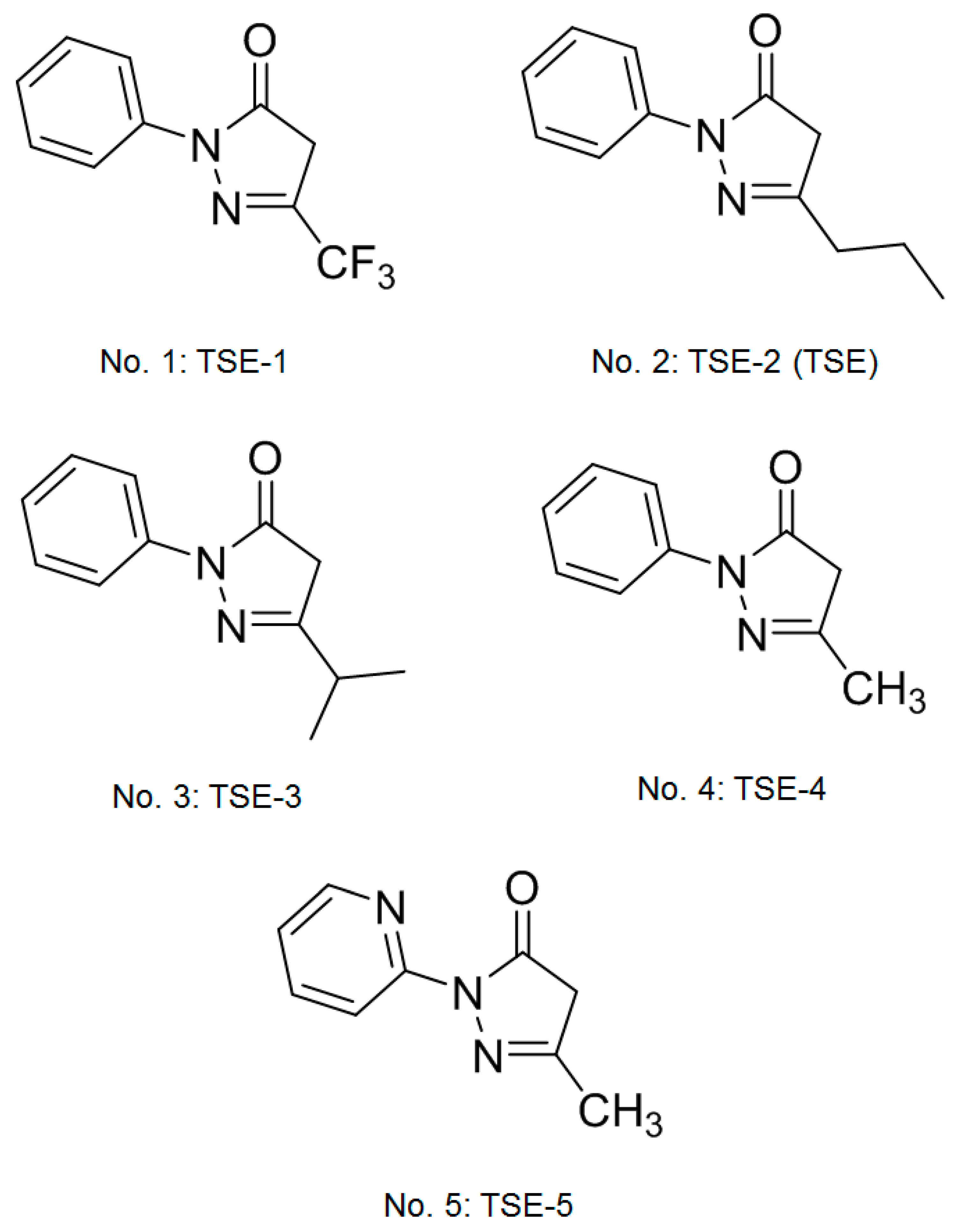
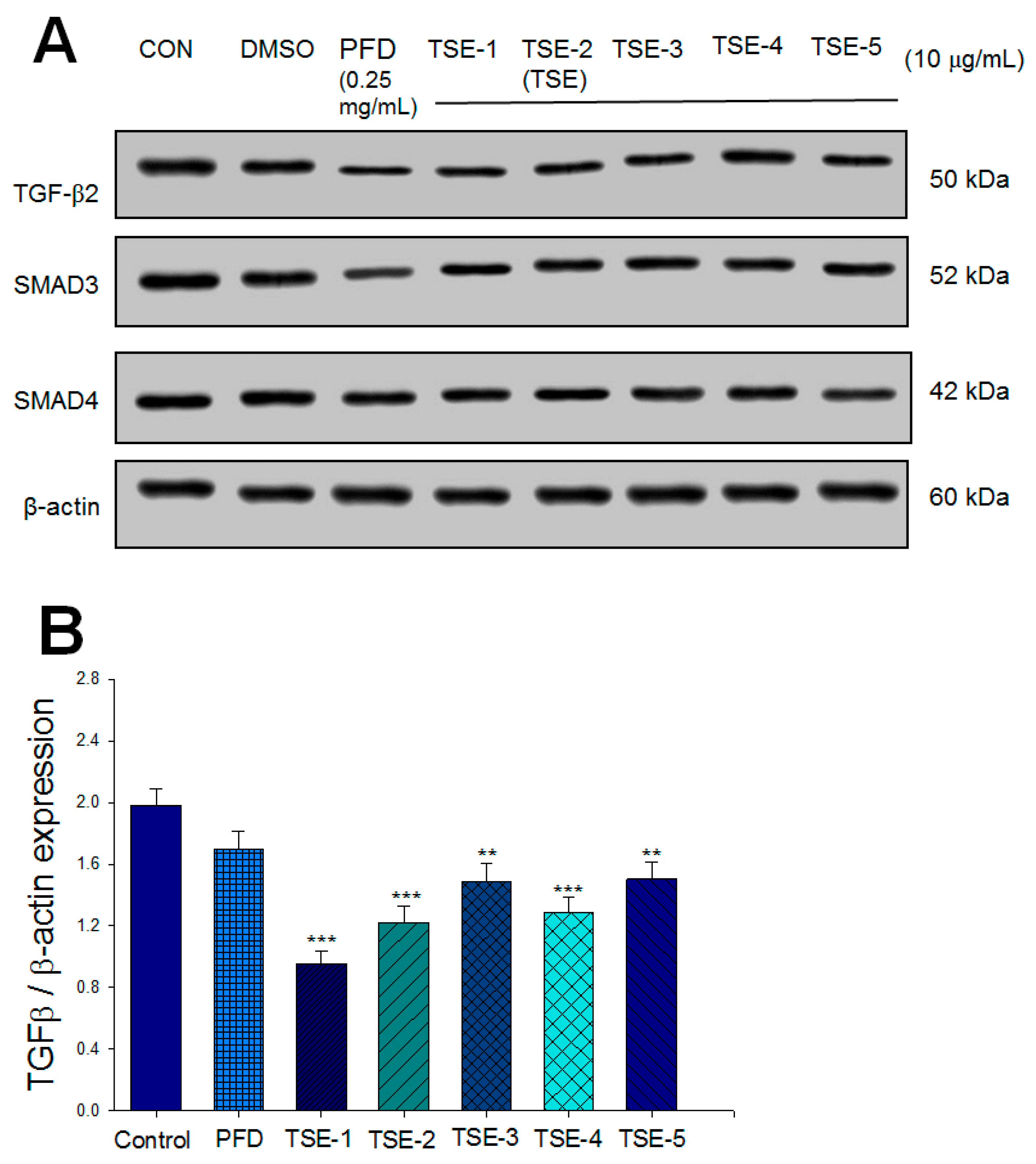
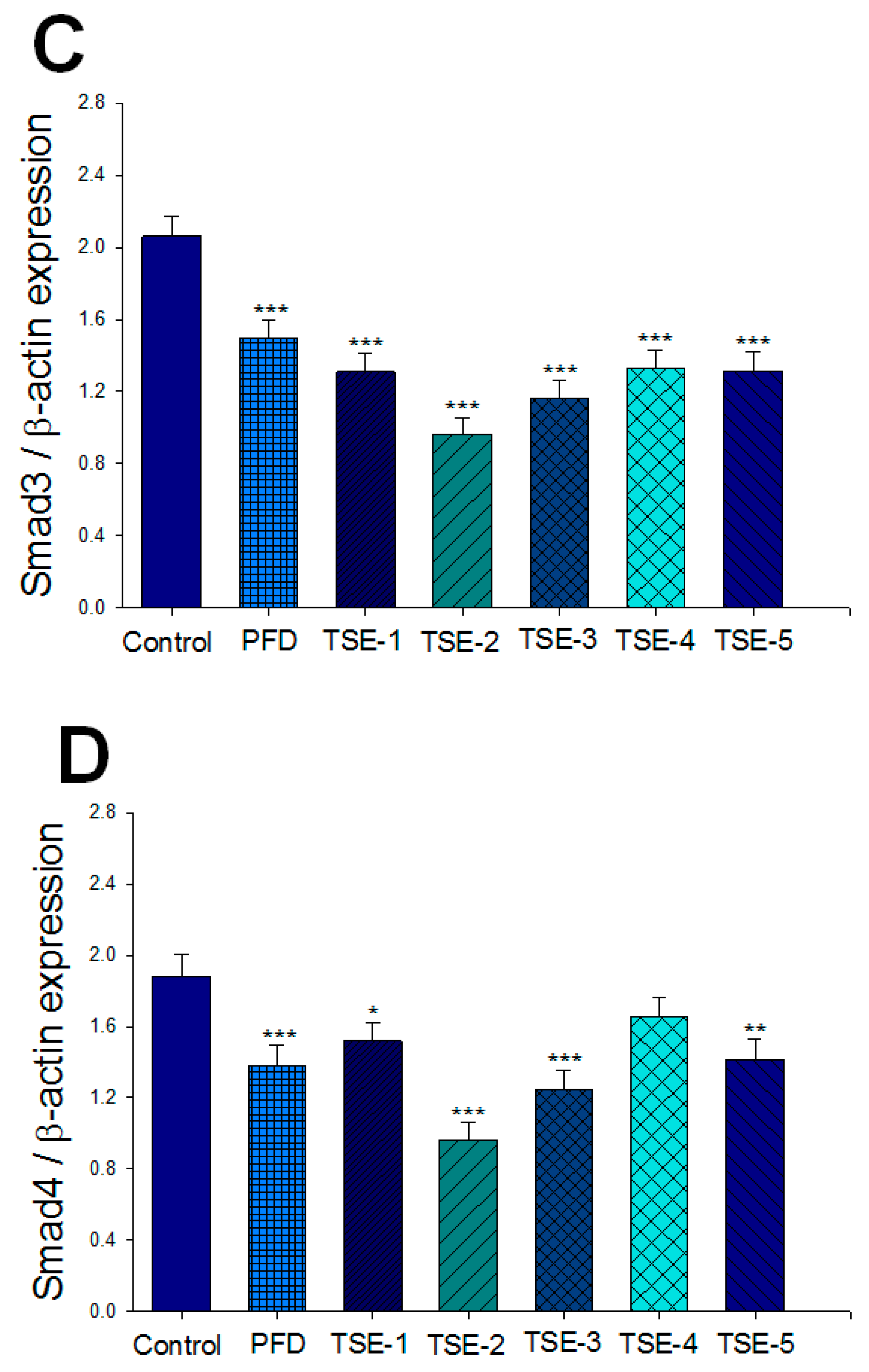
3.1. Effects of Five Compounds on the Expressions of TGF-β2, SMAD3, or SMAD4 in SRA01/04 Cells
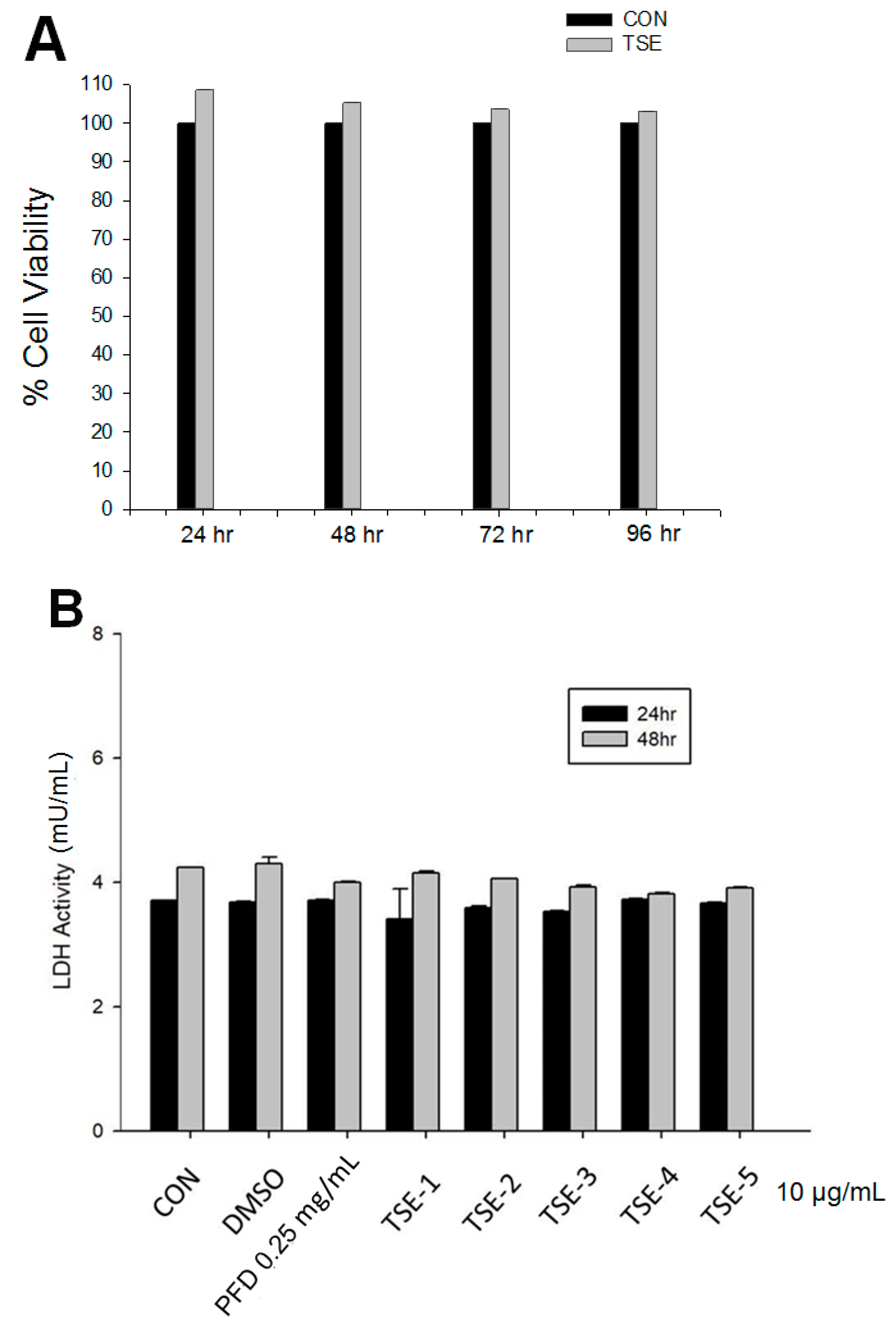
3.2. Cell Viability of Five Compounds
3.3. LDH Assay
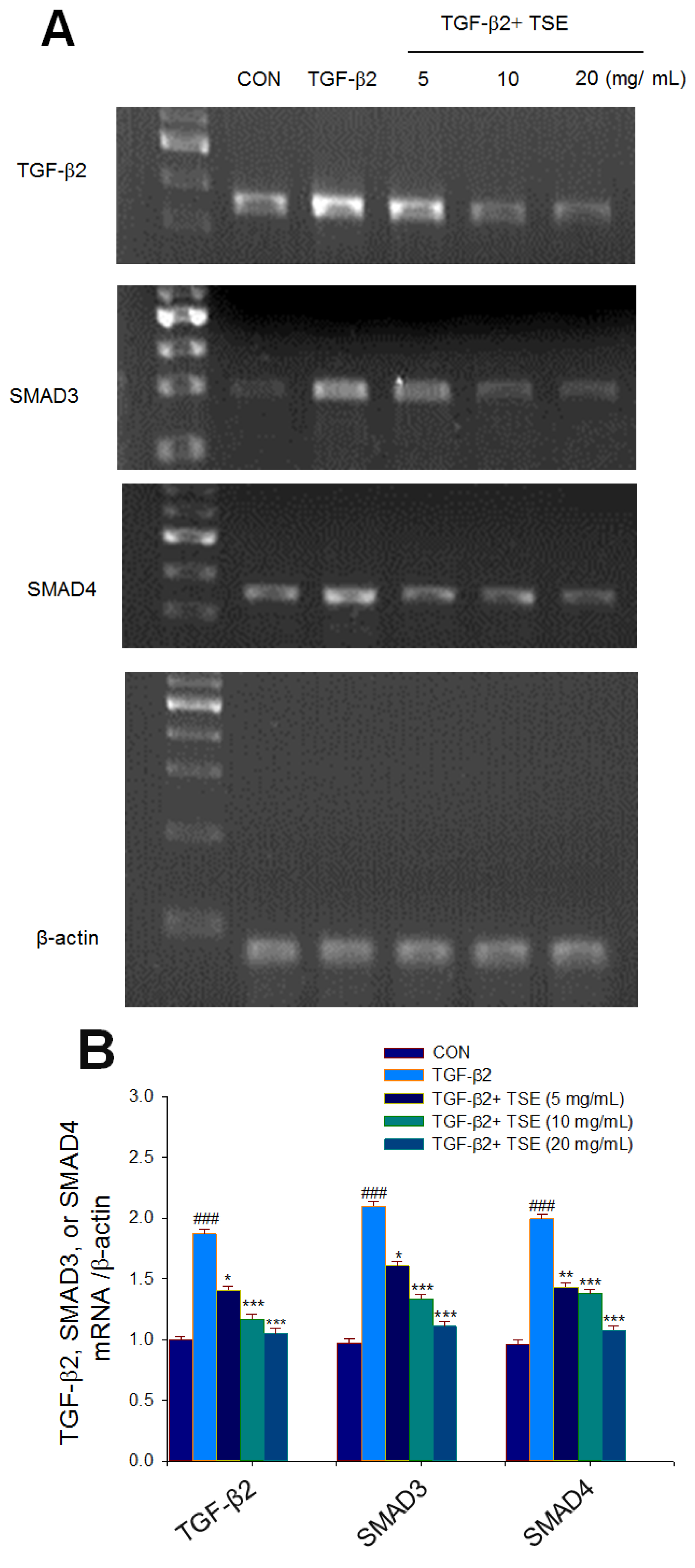
3.4. Effects of TSE on the mRNA Levels of Targeted Genes in Cells
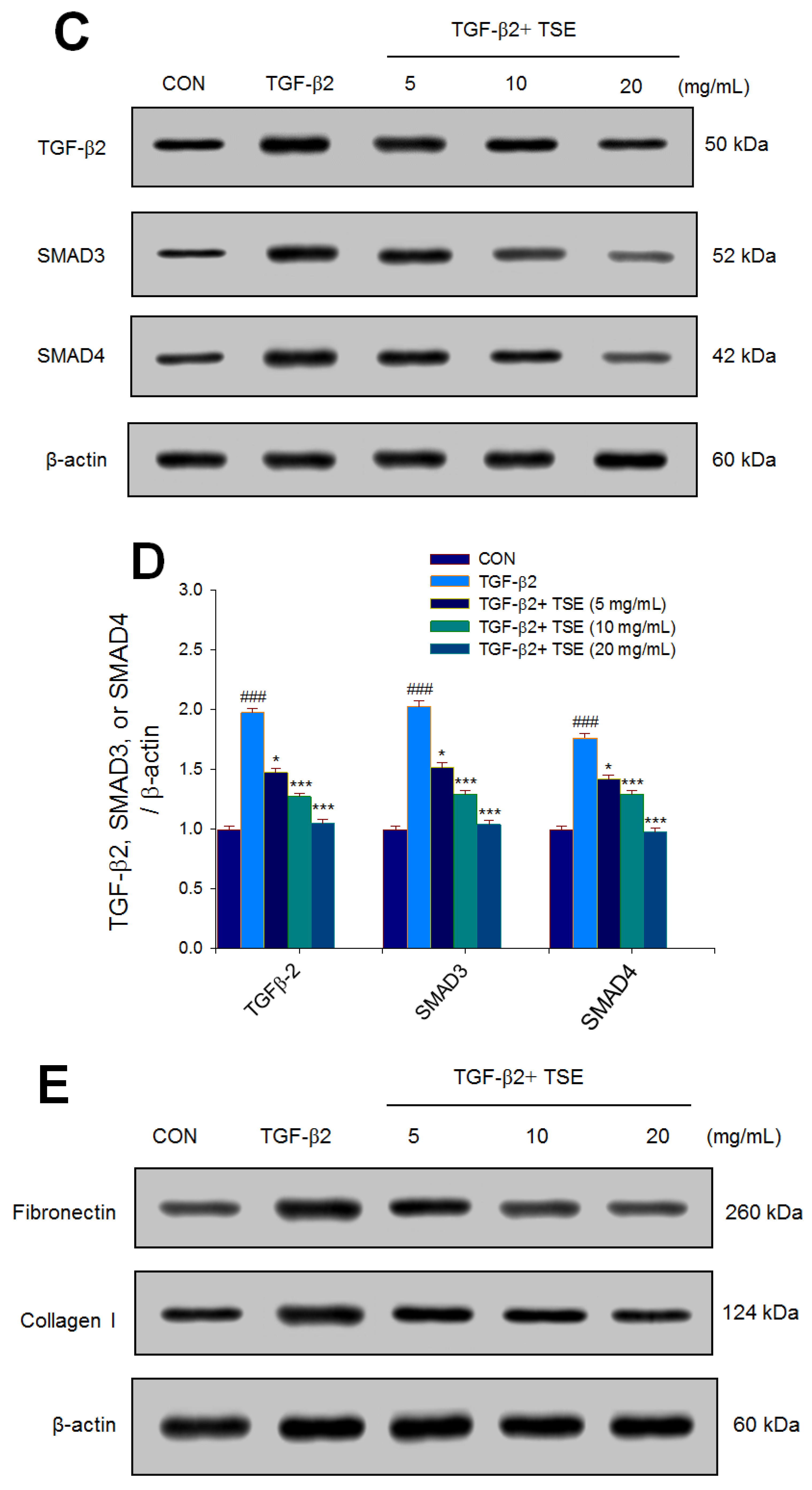
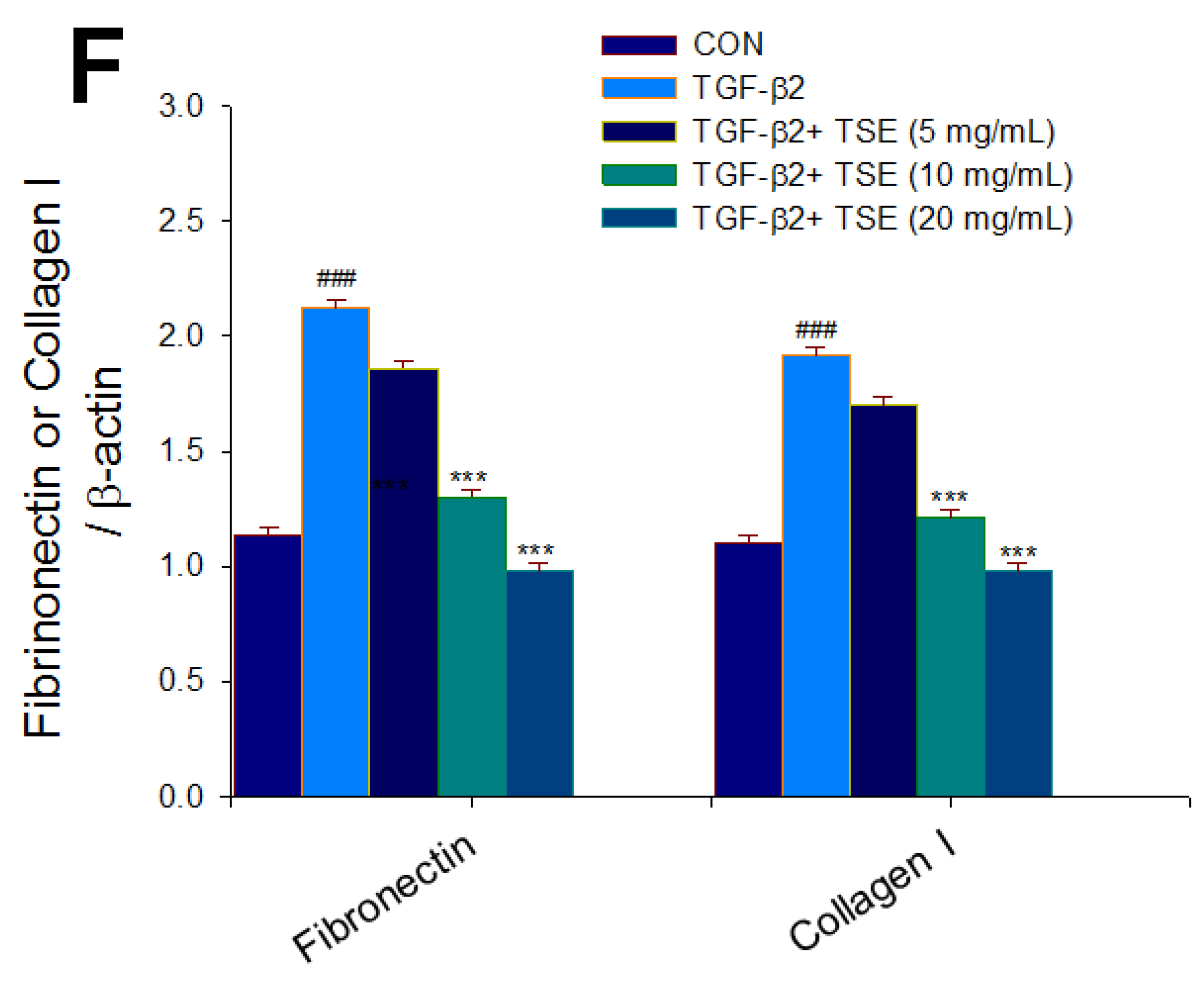
3.5. Effects of TSE on the Expression Levels of TGF-β2, SMADs, Fibronectin, and Collagen I
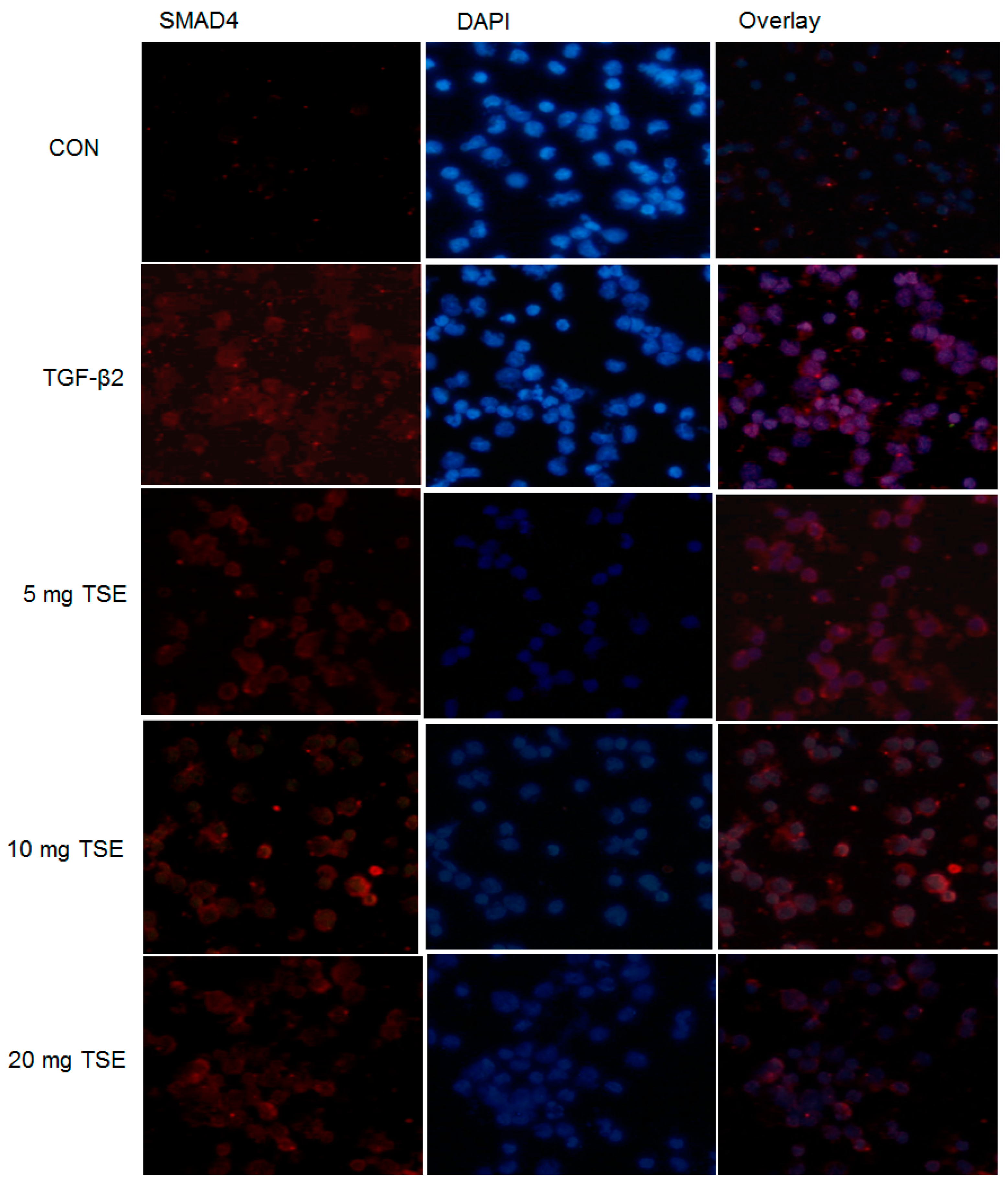
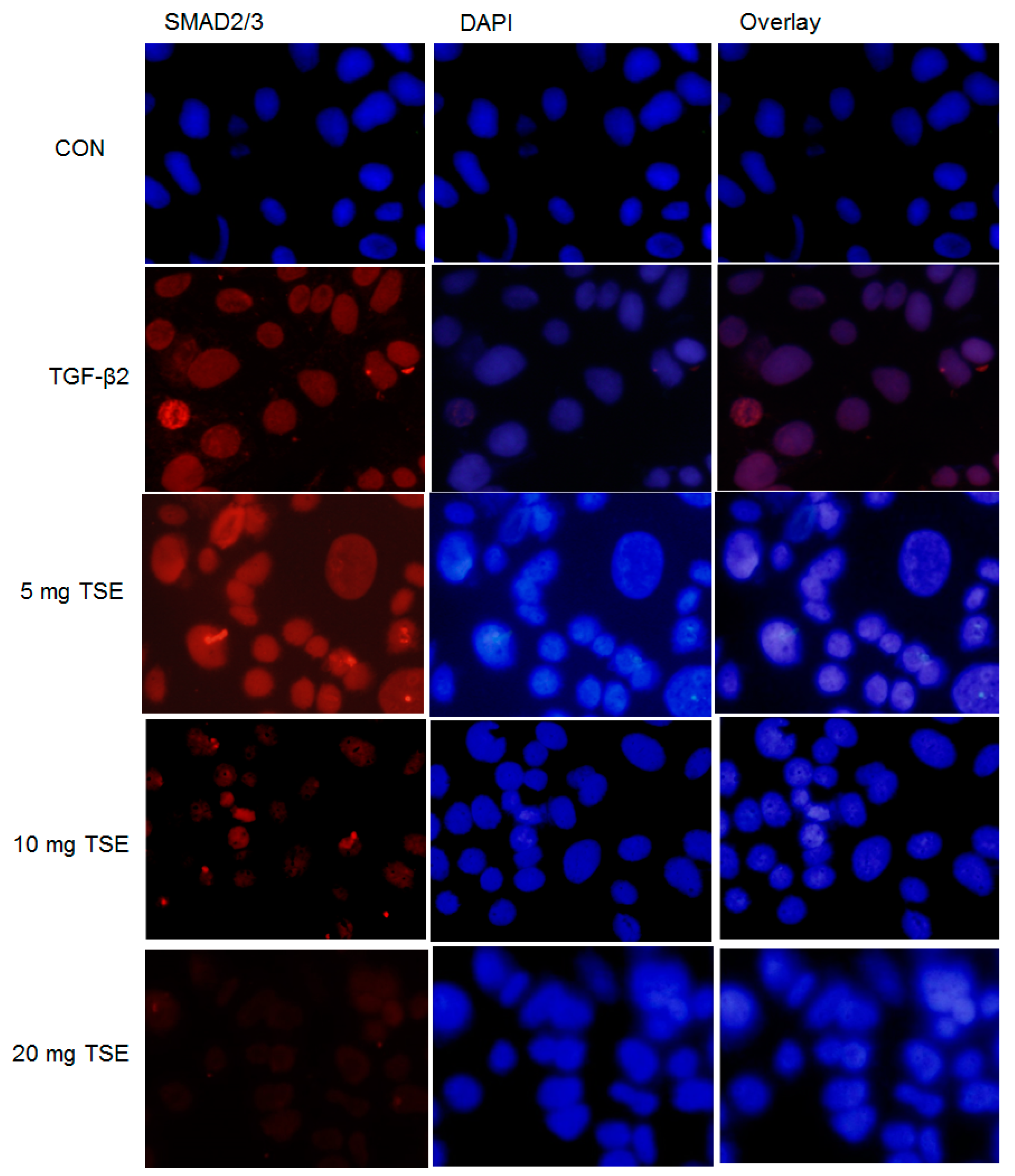
3.6. TSE Inhibits the Nuclear Translocation of SMADs
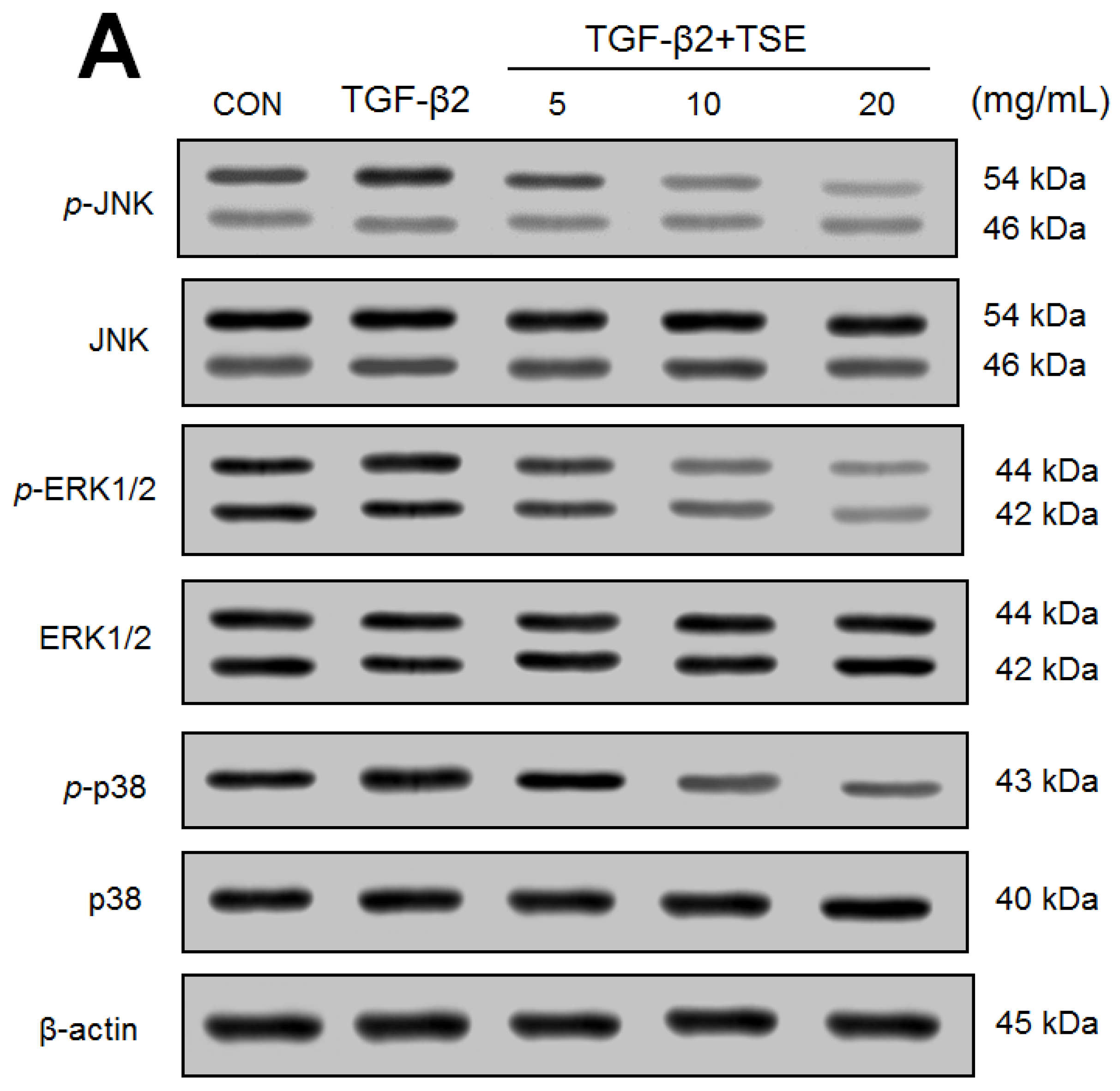
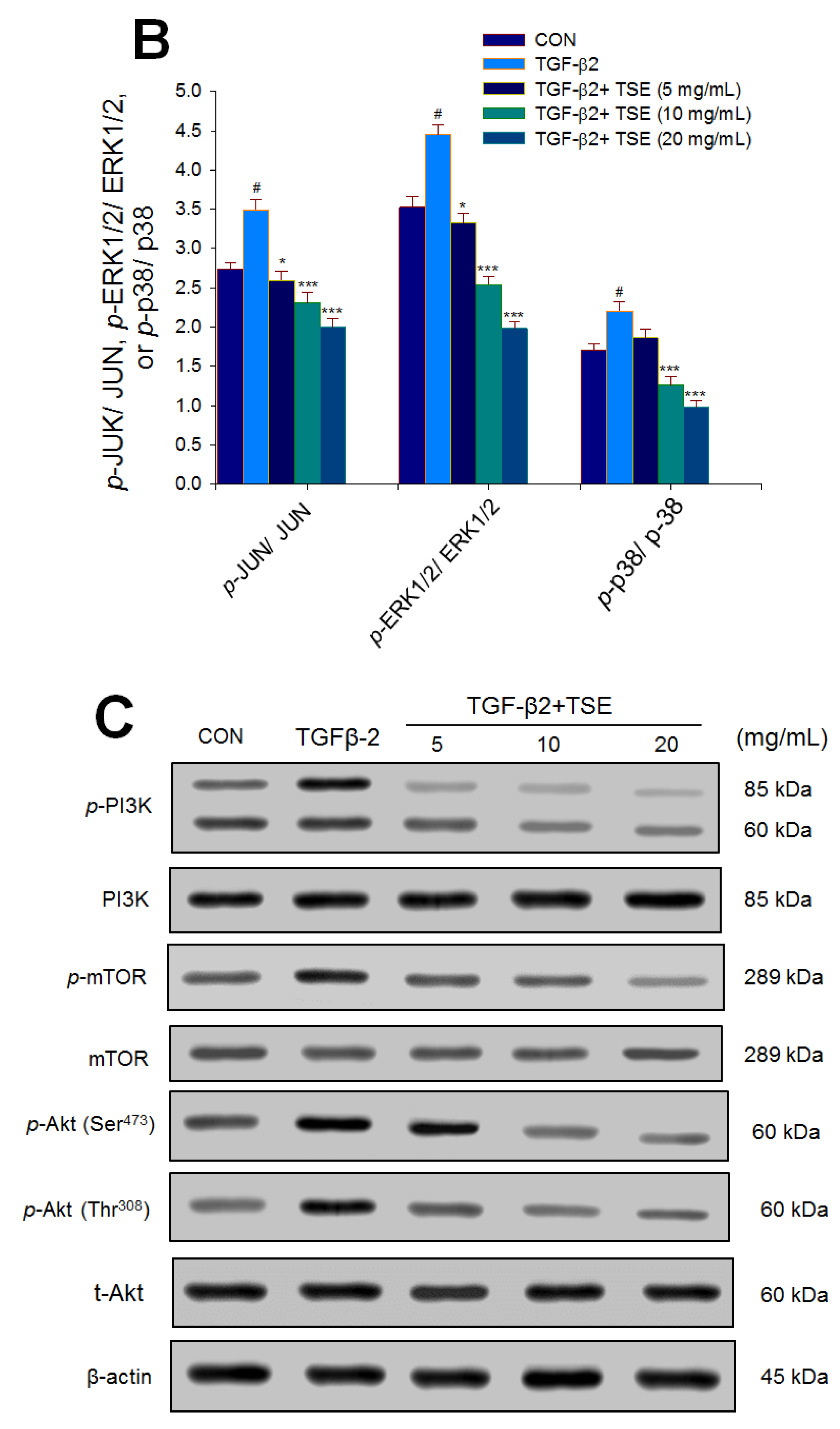
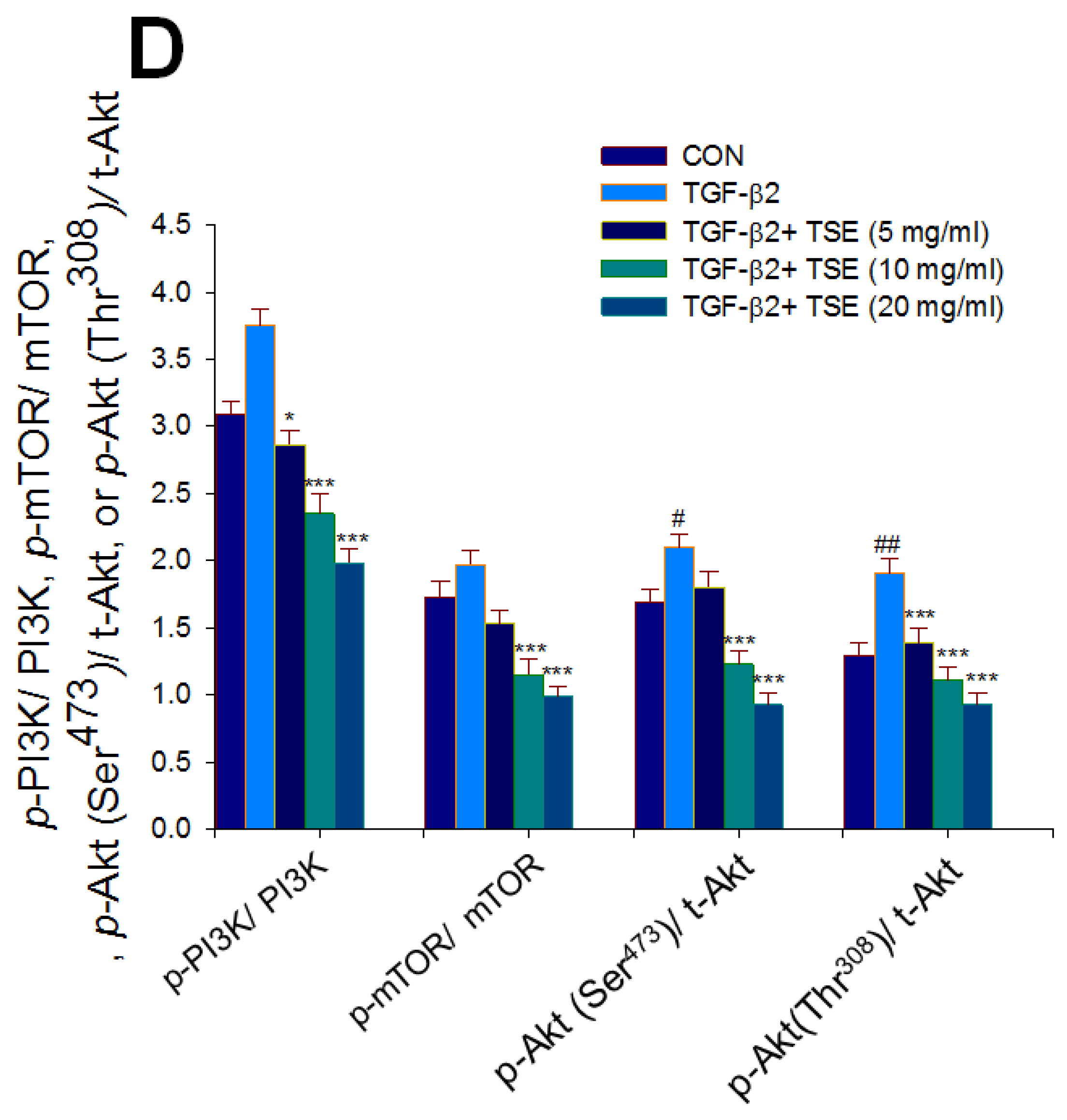
3.7. Effects of TSE on the Protein Expression of p- JUN/JUN, p-ERK1/2/ERK1/2, p-p38/p38, p-Akt (Ser473)/t-Akt, and p-Akt (Thr308)/t-Akt in SRA01/04 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Symonds, J.G.; Lovicu, F.J.; Chamberlain, C.G. Posterior capsule opacification-like changes in rat lens explants cultured with TGFbeta and FGF: Effects of cell coverage and reginal differences. Exp. Eye Res. 2006, 82, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Wormstone, I.M.; Tamiya, S.; Anderson, I.; Duncan, G. TGF-β2-induced matrix modification and cell transdifferentiation in the human lens capsular bag. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2301–2308. [Google Scholar]
- Wormstone, I.M.; Wormstone, Y.M.; Smith, A.J.O.; Eldred, J.A. Posterior capsule opacification: What’s in the bag? Prog. Retin. Eye Res. 2020, 22, 100905. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.J.; Magrath, G.N.; Poole Perry, L.J. Rapid anterior capsule contraction after femtosecond laser-assisted cataract surgery in a patient with retinitis pigmentosa. JCRS Online Case Rep. 2019, 7, 23–25. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, J.J.; Jackson, T.L.; Sun, X.; Wu, W.; Marshall, J. Safety and efficacy of intracapsular tranilast microspheres in experimental posterior capsule opacification. J. Cataract. Refract. Surg. 2007, 33, 2122–2128. [Google Scholar] [CrossRef]
- Pandey, S.K.; Cochener, B.; Apple, D.J.; Colin, J.; Werner, L.; Bougaran, R.; Trivedi, R.H.; Macky, T.A.; Tzak, A.M. Intracapsular ring sustained 5-fluorouracil delivery system for the prevention of posterior capsule opacification in rabbits: A histological study. J. Cataract. Refract. Surg. 2002, 28, 139–148. [Google Scholar] [CrossRef]
- Yu, S.; Lu, C.; Tang, X.; Yuan, X.; Yuan, B.; Yu, Z. Application of spectral domain optical coherence tomography to objectively evaluate posterior capsular opacity in vivo. J. Ophthalmol. 2018, 2018, 5461784. [Google Scholar] [CrossRef] [Green Version]
- Buehl, W.; Findl, O. Intraocular lens design on posterior capsule opacification. J. Cataract. Refract. Surg. 2008, 34, 1976–1985. [Google Scholar] [CrossRef]
- Hazra, S.; Palui, H.; Vemuganti, G.K. Comparison of design of intraocular lens versus the material for PCO prevention. Int. J. Ophthalmol. 2012, 5, 59–63. [Google Scholar]
- Yang, Y.; Ye, Y.; Lin, X.; Wu, K.; Yu, M. Inhibition of pirfenidone on TGF-beta2 induced proliferation, migration and epithelial-mesenchymal transition of human lens epithelial cells line SRA01/04. PLoS ONE 2013, 8, E56837. [Google Scholar] [CrossRef]
- Wu, S.; Tong, N.; Pan, L.; Jiang, X.; Guo, M.; Li, H. Retrospective analyses of potential risk factors for posterior capsule opacification after cataract surgery. J. Ophthalmol. 2018, 2018, 9089285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.H.; Joo, C.K. Role of transforming growth factor-beta in transdifferentiation and fibrosis of lens epithelial cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2025–2032. [Google Scholar]
- Zhang, R.; Li, X.; Liu, Y.; Gao, X.; Zhu, T.; Liu, L. Acceleration of bone regeneration in critical-size defect using BMP-9-loaded nHA/ColI/MWCNTs scaffolds seeded with bone marrow mesenchymal stem cells. Biomed. Res. Int. 2019, 2019, 7343957. [Google Scholar] [CrossRef] [PubMed]
- Dawers, L.J.; Angell, H.; Sleeman, M.; Reddan, J.R.; Wormstone, L.M. TGFbeta isoform dependent SMAD2/3 kinetics in human lens epithelial cells: A cellomics analysis. Exp. Eye Res. 2007, 84, 1009–1012. [Google Scholar] [CrossRef]
- Saika, S.; Yamanaka, O.; Okada, Y.; Tanaka, S.-I.; Miyamoto, T.; Sumioka, T.; Kitano, A.; Shirai, K.; Ikeda, K. TGF beta in fibroproliferative disease in the eye. Front. Biosci. 2009, 1, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Saika, S.; Yamanaka, O.; Flanders, K.C.; Okada, Y.; Miyamoto, T.; Sumioka, T.; Shirai, K.; Kitano, A.; Miyazaki, K.; Tanaka, S.; et al. Epithelial-mesenchymal transition as a therapeutic target for prevention of ocular tissue fibrosis. Endocr. Metab. Immune Disord. Drug Targets 2008, 8, 69–76. [Google Scholar] [CrossRef]
- Katsuno, Y.; Lamouille, S.; Derynck, R. TGF-β signaling and epithelial-mesenchymal transition in cancer progression. Curr. Opin. Oncol. 2013, 25, 76–84. [Google Scholar] [CrossRef]
- Hilberg, O.; Simonsen, U.; Du Bois, R.; Bendstrup, E. Pirfenidone: Significant treatment effects in idiopathic pulmonary fibrosis. Clin. Respir. J. 2012, 6, 131–143. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Lin, H.C.; Cheng, K.M.; Su, W.N.; Sung, K.C.; Lin, T.P.; Huang, J.J.; Lin, S.K.; Wong, F.F. Efficient debromination of 5-pyrazolones and 5-hydroxypyrazoles by N-bromobenzamide. Tetrahedron 2009, 65, 9592–9597. [Google Scholar] [CrossRef]
- Pawlowski, P.; Reszec, J.; Eckstein, A.; Johnson, K.; Grzybowski, A.; Chyczewski, J.; Mysliwiec, L. Markers of inflammation and fibrosis in the orbital fat/connective tissue of patients with Graves’ orbitopathy: Clinical implications. Mediat. Inflamm. 2014, 2014, 412158. [Google Scholar] [CrossRef] [Green Version]
- Ibaraki, N.; Chen, S.C.; Lin, L.R.; Okamoto, H.; Pipas, J.M.; Reddy, V.N. Human lens epithelial cell line. Exp. Eye Res. 1998, 67, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Liu, Y.; Yuan, R.R.; Yu, T.; Yang, B.; Wang, W.Y. Antifibrotic effect of pirfenidone on orbital fibroblasts in patients with thyroid-associated ophthalmopathy and its mechanisms. Zhonghua Nei Ke Za Zhi 2019, 58, 185–190. [Google Scholar] [PubMed]
- Olmos-Zuňiga, J.R.; Baltazares-Lipp, M.; Hernándéz-Jimenez, C.; Victoria, R.J.; Gaxiola-Gaxiola, M.; Silva-Martinez, M.; Iňiguez-Garcia, M.A.; González-González, A.I.; Vázquez-Minero, J.C.; Luna-Flores, A.; et al. Treatment with hyaluronic acid and collagen-polyvinylpyrrolidone improves extracellular matrix assembly for scarring after tracheal resection. Biomed. Res. Int. 2020, 2020, 3964518. [Google Scholar] [CrossRef]
- Mancini, R.; Marucci, L.; Benedertti, A.; Jezeque, A.-M.; Oralndi, F. Immunohistochemical analysis of S-phase cells in normal human and rat liver by PC10 monoclonal antibody. Liver Int. 1994, 14, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kuroda, T.; Hata, S.; Fukagawa, M.; Margolin, S.B.; Kurokawa, K. Pirfenidone improves renal function and fibrosis in the post-obstructed kidney. Kidney Int. 1998, 54, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everett, I.V.T.H.; Olgin, J.E. Atrial fibrosis and the mechanisms of atrial fibrillation. Heart Rhythm 2007, 4, S24–S27. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.N.; Haylor, J. Biological pathways and potential targets for prevention and therapy of chronic allograft nephropathy. Biomed. Res. Int. 2014, 2014, 482438. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.; Lee, K.; Ryu, S.W.; Im, M.; Kook, K.H. Pirfenidone inhibits transforming growth factor-β1-induced fibrogenesis by blocking nuclear translocation of Smads in human retinal pigment epithelial cell line ARPE-19. Mol. Vis. 2012, 18, 1010–1020. [Google Scholar]
- Sutrisno, S.; Sulistyorini, C.; Manungkalit, E.M.; Winarsih, L.; Noorhamdani, N.; Winarsih, S. The effect of genistein on TGF-β signal, dysregulation of apoptosis, cyclooxygenase-2 pathway, and NF-κB pathway in mice peritoneum of endometriosis model. Middle East Fertil. Soc. J. 2017, 22, 295–299. [Google Scholar] [CrossRef]
- Luo, K. Signaling cross talk between TGF-β/ Smad and other signaling pathways. Cold Spring Herb. Perspect. Biol. 2017, 9, a022137. [Google Scholar] [CrossRef] [Green Version]
- Shu, D.Y.; Butcher, E.; Saint-Geniez, M. EMT and EndMT: Emerging roles in age-related macular degeneration. Int. J. Mol. Sci. 2020, 21, 4271. [Google Scholar] [CrossRef] [PubMed]
- Longo, C.M.; Higgins, P.J. Molecular biomarkers of Graves’ ophthalmopathy. Exp. Mol. Pathol. 2019, 106, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.-C.; Hsu, L.-S.; Tsai, J.-H.; Yang, H.-L.; Hsiao, M.-W.; Hwang, J.-M.; Lee, C.-J.; Liu, J.-Y. MZF-1/Elk-1/PKCα is associated with poor prognosis in patients with hepatocellular carcinoma. J. Cancer 2017, 8, 3028–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Xu, Z.; Ling, D.; Li, J.; Wang, Y.; Shan, T. The regulatory role of dietary factors in skeletal muscle development, regeneration and function. Crit. Rev. Food Sci. Nutr. 2020, 62, 33021403. [Google Scholar] [CrossRef] [PubMed]
- Atwood, J.J.; Buck, W.R. Recent literature on bryophytes−123(2). Bryologist 2020, 123, 333–362. [Google Scholar] [CrossRef]
- Lin, C.H.; Kuo, Y.H.; Shih, C.C. Effects of Bofu-tsusho-san on diabetes and hyperlipidemia associated with AMP-activated protein kinase and glucose transporter 4 in high-fat-fed mice. Int. J. Mol. Sci. 2014, 15, 20022–20044. [Google Scholar] [CrossRef] [Green Version]
- Rahimova, N.; Cooke, M.; Zhang, S.; Baker, M.J.; Kazanietz, M.G. The PKC universe keeps expanding: From cancer initiation to metastasis. Adv. Biol. Regul. 2020, 78, 100755. [Google Scholar] [CrossRef]
- Ma, X.; Lin, W.; Lin, Z.; Hao, M.; Gao, X.; Zhang, Y.; Kuang, H. Liraglutide alleviates H2O2-induced retinal ganglion cells injury by inhibiting autophagy through mitochondrial pathways. Peptides 2017, 92, 1–8. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shih, C.-C.; Lee, C.-Y.; Wong, F.-F.; Lin, C.-H. Protective Effects of One 2,4-Dihydro-3H-Pyrazol-3-one Derivative against Posterior Capsular Opacification by Regulation of TGF-β2/SMADs and Non-SMAD Signaling, Collagen I, and Fibronectin Proteins. Curr. Issues Mol. Biol. 2022, 44, 5048-5066. https://doi.org/10.3390/cimb44100343
Shih C-C, Lee C-Y, Wong F-F, Lin C-H. Protective Effects of One 2,4-Dihydro-3H-Pyrazol-3-one Derivative against Posterior Capsular Opacification by Regulation of TGF-β2/SMADs and Non-SMAD Signaling, Collagen I, and Fibronectin Proteins. Current Issues in Molecular Biology. 2022; 44(10):5048-5066. https://doi.org/10.3390/cimb44100343
Chicago/Turabian StyleShih, Chun-Ching, Chia-Yi Lee, Fung-Fuh Wong, and Cheng-Hsiu Lin. 2022. "Protective Effects of One 2,4-Dihydro-3H-Pyrazol-3-one Derivative against Posterior Capsular Opacification by Regulation of TGF-β2/SMADs and Non-SMAD Signaling, Collagen I, and Fibronectin Proteins" Current Issues in Molecular Biology 44, no. 10: 5048-5066. https://doi.org/10.3390/cimb44100343