Decreased Brain Serotonin in rbfox1 Mutant Zebrafish and Partial Reversion of Behavioural Alterations by the SSRI Fluoxetine

 , and
, and
Abstract
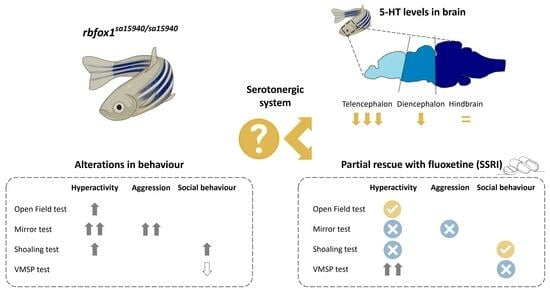
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Neurotransmitter Alterations in rbfox1sa15940/sa15940 Zebrafish
2.2. Partial Reversal of Behavioural Alterations in rbfox1sa15940/sa15940 Zebrafish with Fluoxetine
2.2.1. Open Field
2.2.2. Aggressive Behaviour of rbfox1sa15940/sa15940 Zebrafish under the Influence of Fluoxetine
2.2.3. Social Behaviour of rbfox1sa15940/sa15940 Zebrafish under the Influence of Fluoxetine
Social Novelty Preference Test
Shoaling
3. Discussion
3.1. Serotonin Imbalances Associated with the rbfox1sa15940 Mutation
3.2. Influence of Fluoxetine on Zebrafish Behaviour
3.3. Limitations of This Study
4. Materials and Methods
4.1. Zebrafish Lines and Husbandry
4.2. Quantification of Basal Serotonin and Tryptophan Levels
4.3. Drugs
4.4. Behavioural Experiments
4.4.1. Open Field
4.4.2. Mirror Test
4.4.3. Social Novelty Preference Test
4.4.4. Shoaling
4.5. Statistical Methods and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fogel, B.L.; Wexler, E.; Wahnich, A.; Friedrich, T.; Vijayendran, C.; Gao, F.; Parikshak, N.; Konopka, G.; Geschwind, D.H. RBFOX1 Regulates Both Splicing and Transcriptional Networks in Human Neuronal Development. Hum. Mol. Genet. 2012, 21, 4171–4186. [Google Scholar] [CrossRef]
- Conboy, J.G. Developmental Regulation of RNA Processing by Rbfox Proteins. WIREs RNA 2017, 8, e1398. [Google Scholar] [CrossRef]
- Bill, B.R.; Lowe, J.K.; DyBuncio, C.T.; Fogel, B.L. Orchestration of Neurodevelopmental Programs by RBFOX1. In International Review of Neurobiology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 113, pp. 251–267. ISBN 978-0-12-418700-9. [Google Scholar]
- Zhao, W.-W. Intragenic Deletion of RBFOX1 Associated with Neurodevelopmental/Neuropsychiatric Disorders and Possibly Other Clinical Presentations. Mol. Cytogenet. 2013, 6, 26. [Google Scholar] [CrossRef]
- Zhang-James, Y.; Fernàndez-Castillo, N.; Hess, J.L.; Malki, K.; Glatt, S.J.; Cormand, B.; Faraone, S.V. An Integrated Analysis of Genes and Functional Pathways for Aggression in Human and Rodent Models. Mol. Psychiatry 2019, 24, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.H.; Anttila, V.; Won, H.; Feng, Y.-C.A.; Rosenthal, J.; Zhu, Z.; Tucker-Drob, E.M.; Nivard, M.G.; Grotzinger, A.D.; Posthuma, D.; et al. Genomic Relationships, Novel Loci, and Pleiotropic Mechanisms across Eight Psychiatric Disorders. Cell 2019, 179, 1469–1482.e11. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, A.; Fernàndez-Castillo, N.; Gan, G.; Yang, Y.; Yotova, A.Y.; Kranz, T.M.; Grünewald, L.; Freudenberg, F.; Antón-Galindo, E.; Cabana-Domínguez, J.; et al. Behavioural and Functional Evidence Revealing the Role of RBFOX1 Variation in Multiple Psychiatric Disorders and Traits. Mol. Psychiatry 2022, 27, 4464–4473. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-A.; Damianov, A.; Lin, C.-H.; Fontes, M.; Parikshak, N.N.; Anderson, E.S.; Geschwind, D.H.; Black, D.L.; Martin, K.C. Cytoplasmic Rbfox1 Regulates the Expression of Synaptic and Autism-Related Genes. Neuron 2016, 89, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Grotzinger, A.D.; Mallard, T.T.; Akingbuwa, W.A.; Ip, H.F.; Adams, M.J.; Lewis, C.M.; McIntosh, A.M.; Grove, J.; Dalsgaard, S.; Lesch, K.-P.; et al. Genetic Architecture of 11 Major Psychiatric Disorders at Biobehavioral, Functional Genomic and Molecular Genetic Levels of Analysis. Nat. Genet. 2022, 54, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Ren, Y.; Fan, L.; Zhao, S.; Feng, J.; Zhong, Q.; Tu, D.; Wu, W.; Chen, J.; Xie, P. TCF4 and RBFOX1 as Peripheral Biomarkers for the Differential Diagnosis and Treatment of Major Depressive Disorder. J. Affect. Disord. 2024, 345, 252–261. [Google Scholar] [CrossRef]
- Antón-Galindo, E.; Adel, M.R.; García-Gonzalez, J.; Leggieri, A.; López-Blanch, L.; Irimia, M.; Norton, W.H.; Brennan, C.H.; Fernàndez-Castillo, N.; Cormand, B. Pleiotropic Contribution of Rbfox1 to Psychiatric and Neurodevelopmental Phenotypes in two Zebrafish Models. Transl. Psychiatry, 2024; in press. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an Emerging Model for Studying Complex Brain Disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef]
- MacRae, C.A.; Peterson, R.T. Zebrafish as Tools for Drug Discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef]
- Correia, D.; Domingues, I.; Faria, M.; Oliveira, M. Effects of Fluoxetine on Fish: What Do We Know and Where Should We Focus Our Efforts in the Future? Sci. Total Environ. 2023, 857, 159486. [Google Scholar] [CrossRef]
- Baldwin, D.S.; Waldman, S.; Allgulander, C. Evidence-Based Pharmacological Treatment of Generalized Anxiety Disorder. Int. J. Neuropsychopharm. 2011, 14, 697–710. [Google Scholar] [CrossRef]
- Davies, M.N.; Verdi, S.; Burri, A.; Trzaskowski, M.; Lee, M.; Hettema, J.M.; Jansen, R.; Boomsma, D.I.; Spector, T.D. Generalised Anxiety Disorder—A Twin Study of Genetic Architecture, Genome-Wide Association and Differential Gene Expression. PLoS ONE 2015, 10, e0134865. [Google Scholar] [CrossRef]
- Gelenberg, A.J.; Freeman, M.; Markowitz, J.; Rosenbaum, J.; Thase, M.; Trivedi, M.; Van Rhoads, R. American Psychiatric Association Practice Guidelines for the Treatment of Patients with Major Depressive Disorder. Am. J. Psychiatry 2010, 167, 9–118. [Google Scholar]
- Del Casale, A.; Sorice, S.; Padovano, A.; Simmaco, M.; Ferracuti, S.; Lamis, D.A.; Rapinesi, C.; Sani, G.; Girardi, P.; Kotzalidis, G.D.; et al. Psychopharmacological Treatment of Obsessive-Compulsive Disorder (OCD). Curr. Neuropharmacol. 2019, 17, 710–736. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.M.; Grossman, L.; Nguyen, M.; Maximino, C.; Rosemberg, D.B.; Echevarria, D.J.; Kalueff, A.V. Aquatic Toxicology of Fluoxetine: Understanding the Knowns and the Unknowns. Aquat. Toxicol. 2014, 156, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.S.D.; Koakoski, G.; Ferreira, D.; Oliveira, T.A.; Rosa, J.G.S.D.; Gusso, D.; Giacomini, A.C.V.; Piato, A.L.; Barcellos, L.J.G. Diazepam and Fluoxetine Decrease the Stress Response in Zebrafish. PLoS ONE 2014, 9, e103232. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, A.C.V.V.; Abreu, M.S.; Giacomini, L.V.; Siebel, A.M.; Zimerman, F.F.; Rambo, C.L.; Mocelin, R.; Bonan, C.D.; Piato, A.L.; Barcellos, L.J.G. Fluoxetine and Diazepam Acutely Modulate Stress Induced-Behavior. Behav. Brain Res. 2016, 296, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Herculano, A.M.; Maximino, C. Serotonergic Modulation of Zebrafish Behavior: Towards a Paradox. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Lillesaar, C. The Serotonergic System in Fish. J. Chem. Neuroanat. 2011, 41, 294–308. [Google Scholar] [CrossRef]
- Gabriel, J.P.; Mahmood, R.; Kyriakatos, A.; Söll, I.; Hauptmann, G.; Calabrese, R.L.; El Manira, A. Serotonergic Modulation of Locomotion in Zebrafish—Endogenous Release and Synaptic Mechanisms. J. Neurosci. 2009, 29, 10387–10395. [Google Scholar] [CrossRef]
- Fernàndez-Castillo, N.; Gan, G.; Van Donkelaar, M.M.J.; Vaht, M.; Weber, H.; Retz, W.; Meyer-Lindenberg, A.; Franke, B.; Harro, J.; Reif, A.; et al. RBFOX1, Encoding a Splicing Regulator, Is a Candidate Gene for Aggressive Behavior. Eur. Neuropsychopharmacol. 2020, 30, 44–55. [Google Scholar] [CrossRef]
- Perez-Caballero, L.; Torres-Sanchez, S.; Bravo, L.; Mico, J.A.; Berrocoso, E. Fluoxetine: A Case History of Its Discovery and Preclinical Development. Expert Opin. Drug Discov. 2014, 9, 567–578. [Google Scholar] [CrossRef]
- Gaspar, P.; Lillesaar, C. Probing the Diversity of Serotonin Neurons. Philos. Trans. R. Soc. B 2012, 367, 2382–2394. [Google Scholar] [CrossRef]
- Cheng, R.-K.; Jesuthasan, S.J.; Penney, T.B. Zebrafish Forebrain and Temporal Conditioning. Philos. Trans. R. Soc. B 2014, 369, 20120462. [Google Scholar] [CrossRef]
- Giacomini, A.C.V.V.; Piassetta, A.S.; Genario, R.; Bonan, C.D.; Piato, A.; Barcellos, L.J.G.; De Abreu, M.S. Tryptophan Alleviates Neuroendocrine and Behavioral Responses to Stress in Zebrafish. Behav. Brain Res. 2020, 378, 112264. [Google Scholar] [CrossRef] [PubMed]
- Al Shuraiqi, A.; Al-Habsi, A.; Barry, M.J. Time-, Dose- and Transgenerational Effects of Fluoxetine on the Behavioural Responses of Zebrafish to a Conspecific Alarm Substance. Environ. Pollut. 2021, 270, 116164. [Google Scholar] [CrossRef] [PubMed]
- Winberg, S.; Thörnqvist, P.-O. Role of Brain Serotonin in Modulating Fish Behavior. Curr. Zool. 2016, 62, 317–323. [Google Scholar] [CrossRef]
- Jones, L.J.; Norton, W.H.J. Using Zebrafish to Uncover the Genetic and Neural Basis of Aggression, a Frequent Comorbid Symptom of Psychiatric Disorders. Behav. Brain Res. 2015, 276, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Oliva, C.; Arias, A.; Ruiz, M.; Pujol, A.; Garrabou, G.; Canto-Santos, J.; Urreizti, R.; Castilla-Vallmanya, L.; Rodriguez-Gonzalez, H.; Jou, C.; et al. Fibroblast Phenylalanine Concentration as a Surrogate Biomarker of Cellular Number. J. Chromatogr. B 2023, 1226, 123787. [Google Scholar] [CrossRef] [PubMed]
- Casado, M.; Sierra, C.; Batllori, M.; Artuch, R.; Ormazabal, A. A Targeted Metabolomic Procedure for Amino Acid Analysis in Different Biological Specimens by Ultra-High-Performance Liquid Chromatography–Tandem Mass Spectrometry. Metabolomics 2018, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D.K.; Bhat, A. Alone but Not Always Lonely: Social Cues Alleviate Isolation Induced Behavioural Stress in Wild Zebrafish. Appl. Anim. Behav. Sci. 2022, 251, 105623. [Google Scholar] [CrossRef]
- Romero-Ferrero, F.; Bergomi, M.G.; Hinz, R.C.; Heras, F.J.H.; De Polavieja, G.G. Idtracker.Ai: Tracking All Individuals in Small or Large Collectives of Unmarked Animals. Nat. Methods 2019, 16, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Norton, W.H.J.; Stumpenhorst, K.; Faus-Kessler, T.; Folchert, A.; Rohner, N.; Harris, M.P.; Callebert, J.; Bally-Cuif, L. Modulation of Fgfr1a Signaling in Zebrafish Reveals a Genetic Basis for the Aggression–Boldness Syndrome. J. Neurosci. 2011, 31, 13796–13807. [Google Scholar] [CrossRef]
- Carreño Gutiérrez, H.; Colanesi, S.; Cooper, B.; Reichmann, F.; Young, A.M.J.; Kelsh, R.N.; Norton, W.H.J. Endothelin Neurotransmitter Signalling Controls Zebrafish Social Behaviour. Sci. Rep. 2019, 9, 3040. [Google Scholar] [CrossRef]
- Parker, M.O.; Brock, A.J.; Millington, M.E.; Brennan, C.H. Behavioral Phenotyping of Casper Mutant and 1-Pheny-2-Thiourea Treated Adult Zebrafish. Zebrafish 2013, 10, 466–471. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adel, M.R.; Antón-Galindo, E.; Gago-Garcia, E.; Arias-Dimas, A.; Arenas, C.; Artuch, R.; Cormand, B.; Fernàndez-Castillo, N. Decreased Brain Serotonin in rbfox1 Mutant Zebrafish and Partial Reversion of Behavioural Alterations by the SSRI Fluoxetine. Pharmaceuticals 2024, 17, 254. https://doi.org/10.3390/ph17020254
Adel MR, Antón-Galindo E, Gago-Garcia E, Arias-Dimas A, Arenas C, Artuch R, Cormand B, Fernàndez-Castillo N. Decreased Brain Serotonin in rbfox1 Mutant Zebrafish and Partial Reversion of Behavioural Alterations by the SSRI Fluoxetine. Pharmaceuticals. 2024; 17(2):254. https://doi.org/10.3390/ph17020254
Chicago/Turabian StyleAdel, Maja R., Ester Antón-Galindo, Edurne Gago-Garcia, Angela Arias-Dimas, Concepció Arenas, Rafael Artuch, Bru Cormand, and Noèlia Fernàndez-Castillo. 2024. "Decreased Brain Serotonin in rbfox1 Mutant Zebrafish and Partial Reversion of Behavioural Alterations by the SSRI Fluoxetine" Pharmaceuticals 17, no. 2: 254. https://doi.org/10.3390/ph17020254