



Dipeptidyl Peptidase IV Inhibitory Peptides from Chickpea Proteins (Cicer arietinum L.): Pharmacokinetics, Molecular Interactions, and Multi-Bioactivities

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Profile of Peptides Released after Enzymatic Hydrolysis
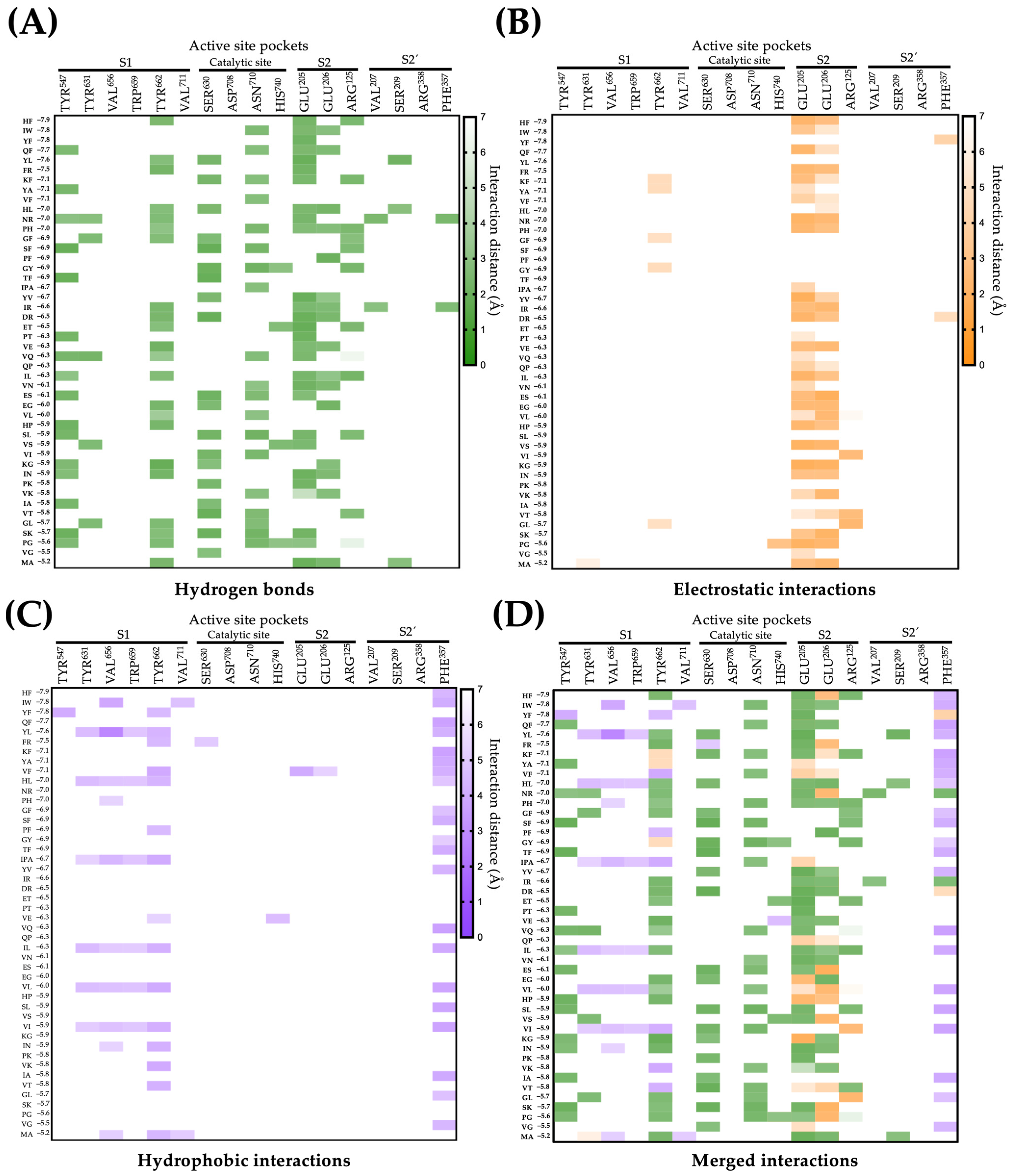
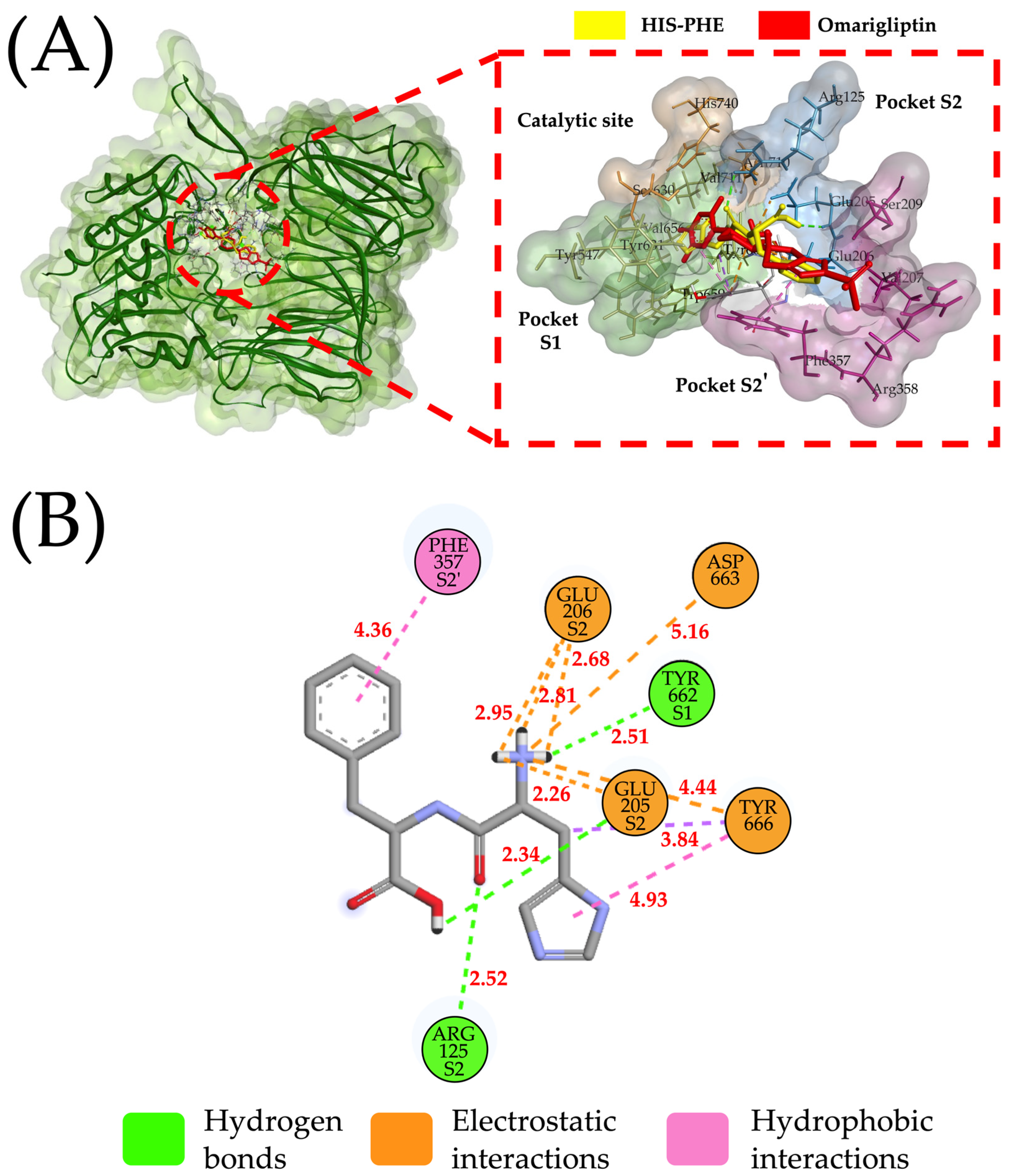
2.2. Molecular Interactions between DPP-IV and Their Inhibitory Peptides
2.3. ADMET Properties
2.4. Multi-Bioactivities and Stability for Gastrointestinal Digestion
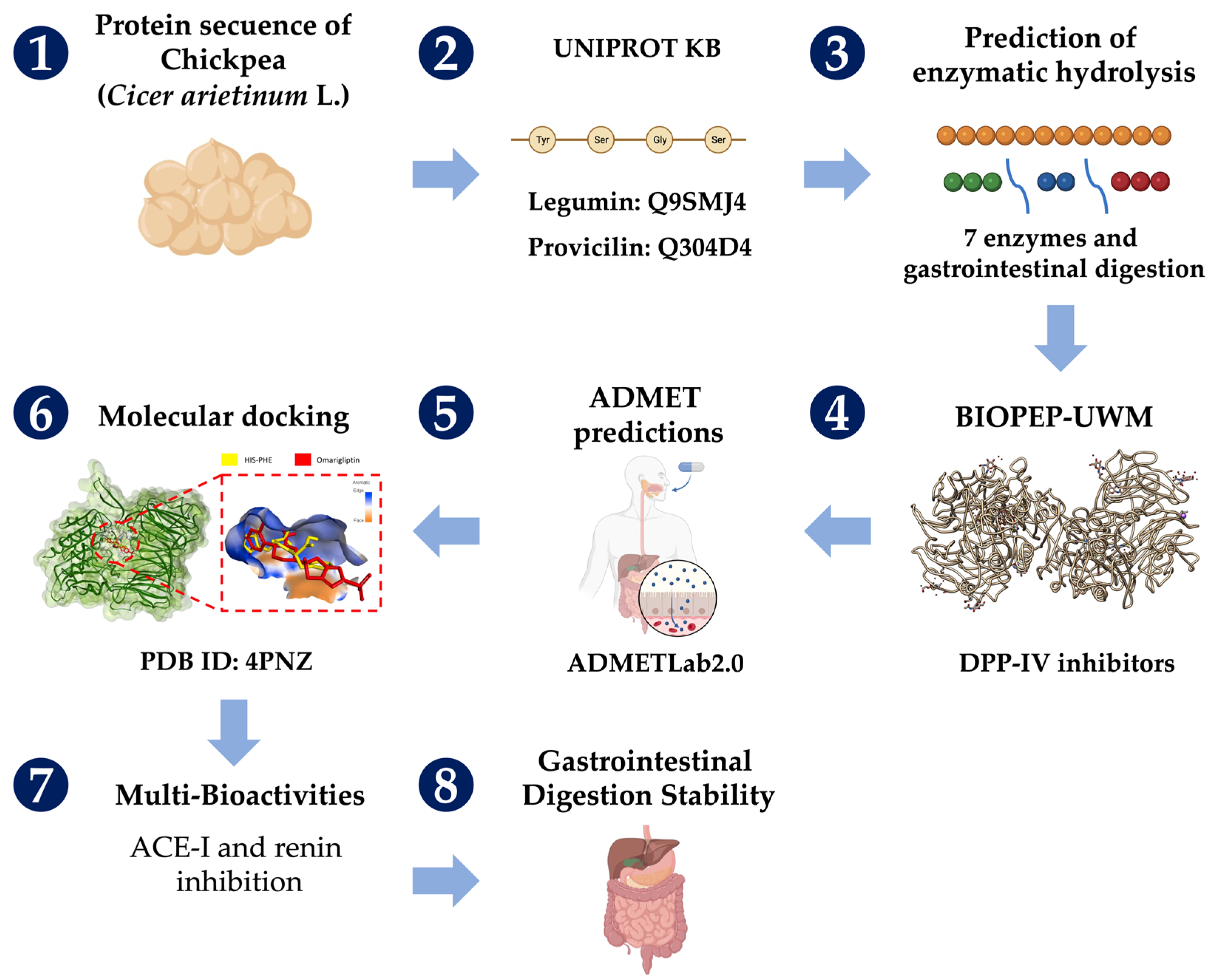
3. Materials and Methods
3.1. Protein Sequence, Enzymatic Hydrolyses, and Analyses of DPP-IV Inhibitory Peptides
3.2. ADMET Predictions
3.3. Molecular Docking
3.4. Multi-Bioactivities and Stability to Gastrointestinal Digestion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 Diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Magliano, D.J.; Boyko, E.J. IDF Diabetes Atlas 10th Edition Scientific Committee. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021; ISBN 9782930229980. [Google Scholar]
- Kasina, S.V.S.K.; Baradhi, K.M. Dipeptidyl Peptidase IV (DPP IV) Inhibitors; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Sterrett, J.J.; Bragg, S.; Weart, C.W. Type 2 Diabetes Medication Review. Am. J. Med. Sci. 2016, 351, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. An in Silico Model to Predict the Potential of Dietary Proteins as Sources of Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides. Food Chem. 2014, 165, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boer, G.A.; Holst, J.J. Incretin Hormones and Type 2 Diabetes—Mechanistic Insights and Therapeutic Approaches. Biology 2020, 9, 473. [Google Scholar] [CrossRef]
- Jia, L.; Wang, L.; Liu, C.; Liang, Y.; Lin, Q. Bioactive Peptides from Foods: Production, Function, and Application. Food Funct. 2021, 12, 7108–7712. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Oh, D.H.; Lee, B.H. Bioactive Peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, M.; Awad, S. Milk Bioactive Peptides: Antioxidant, Antimicrobial and Anti-Diabetic Activities. Adv Biochem. 2019, 7, 22–33. [Google Scholar] [CrossRef]
- Acevedo Martínez, K.A.; Gonzalezde Mejia, E. Comparison of Five Chickpea Varieties, Optimization of Hydrolysates Production and Evaluation of Biomarkers for Type 2 Diabetes. Food Res. Int. 2021, 147, 110572. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; de Mejia, E.G. Optimization, Identification, and Comparison of Peptides from Germinated Chickpea (Cicer rietinum) Protein Hydrolysates Using Either Papain or Ficin and Their Relationship with Markers of Type 2 Diabetes. Food Chem. 2022, 374, 131717. [Google Scholar] [CrossRef]
- Farias, T.C.; de Souza, T.S.P.; Fai, A.E.C.; Koblitz, M.G.B. Critical Review for the Production of Antidiabetic Peptides by a Bibliometric Approach. Nutrients. 2022, 14, 4275. [Google Scholar] [CrossRef]
- Real Hernandez, L.M.; Gonzalez de Mejia, E. Enzymatic Production, Bioactivity, and Bitterness of Chickpea (Cicer arietinum) Peptides. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1913–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morellon-Sterling, R.; El-Siar, H.; Tavano, O.L.; Berenguer-Murcia, Á.; Fernández-Lafuente, R. Ficin: A Protease Extract with Relevance in Biotechnology and Biocatalysis. Int. J. Biol. Macromol. 2020, 162, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Nanda, R.F.; Bahar, R.; Syukri, D.; Thu, N.N.A.; Kasim, A. A Review: Application of Bromelain Enzymes in Animal Food Products. Int. J. Agric. Nat. Sci. AIJANS 2020, 1, 33–44. [Google Scholar] [CrossRef]
- Dziuba, J.; Niklewicz, M.; Iwaniak, A.; Darewicz, M.; Minkiewicz, P. Bioinformatic-Aided Prediction for Release Possibilities of Bioactive Peptides from Plant Proteins. Acta Aliment. 2004, 33, 227–235. [Google Scholar] [CrossRef]
- Ma, C.; Liu, D.; Hao, H.; Wu, X. Identification of the DPP-IV Inhibitory Peptides from Donkey Blood and Regulatory Effect on the Gut Microbiota of Type 2 Diabetic Mice. Foods 2022, 11, 2148. [Google Scholar] [CrossRef]
- Xu, F.; Yao, Y.; Xu, X.; Wang, M.; Pan, M.; Ji, S.; Wu, J.; Jiang, D.; Ju, X.; Wang, L. Identification and Quantification of DPP-IV-Inhibitory Peptides from Hydrolyzed-Rapeseed-Protein-Derived Napin with Analysis of the Interactions between Key Residues and Protein Domains. J. Agric. Food Chem. 2019, 67, 3679–3690. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; Luna-Vital, D.; de Mejia, E.G. Identification and Comparison of Peptides from Chickpea Protein Hydrolysates Using Either Bromelain or Gastrointestinal Enzymes and Their Relationship with Markers of Type 2 Diabetes and Bitterness. Nutrients 2020, 12, 3843. [Google Scholar] [CrossRef]
- Ahmed, T.; Sun, X.; Udenigwe, C.C. Role of Structural Properties of Bioactive Peptides in Their Stability during Simulated Gastrointestinal Digestion: A Systematic Review. Trends Food Sci. Technol. 2022, 120, 265–273. [Google Scholar] [CrossRef]
- Amigo, L.; Hernández-Ledesma, B. Current Evidence on the Bioavailability of Food Bioactive Peptides. Molecules 2020, 25, 4479. [Google Scholar] [CrossRef]
- Ramírez-Torres, G.I.; Ontiveros, N.; López-Teros, V.; Suarez-Jiménez, G.M.; Cabrera-Chávez, F. Food Matrices for the Delivery of Antihypertensive Peptides in Functional Foods. Biotecnia 2018, 20, 165–169. [Google Scholar] [CrossRef]
- Grasso, N.; Lynch, N.L.; Arendt, E.K.; O’Mahony, J.A. Chickpea Protein Ingredients: A Review of Composition, Functionality, and Applications. Compr. Rev. Food Sci. Food Saf. 2022, 21, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Cao, M.-J.; Yu, Y.Q.; Engen, J.R. Accessing the Reproducibility and Specificity of Pepsin and Other Aspartic Proteases. Biochim. Biophys. Acta BBA Proteins Proteom. 2013, 1834, 1222–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of Novel DPP–IV Inhibitory Peptides from Atlantic Salmon (Salmo salar) Skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Gong, H.; Mao, X. Dipeptidyl Peptidase-IV Inhibitory Activity and Related Molecular Mechanism of Bovine α-Lactalbumin-Derived Peptides. Molecules 2020, 25, 3009. [Google Scholar] [CrossRef]
- Gu, H.; Gao, J.; Shen, Q.; Gao, D.; Wang, Q.; Tangyu, M.; Mao, X. Dipeptidyl Peptidase-IV Inhibitory Activity of Millet Protein Peptides and the Related Mechanisms Revealed by Molecular Docking. LWT 2021, 138, 110587. [Google Scholar] [CrossRef]
- Gupta, A.; Jacobson, G.A.; Burgess, J.R.; Jelinek, H.F.; Nichols, D.S.; Narkowicz, C.K.; Al-Aubaidy, H.A. Citrus Bioflavonoids Dipeptidyl Peptidase-4 Inhibition Compared with Gliptin Antidiabetic Medications. Biochem. Biophys. Res. Commun. 2018, 503, 21–25. [Google Scholar] [CrossRef]
- Sneha, P.; Doss, C.G.P. Gliptins in Managing Diabetes-Reviewing Computational Strategy. Life Sci. 2016, 166, 108–120. [Google Scholar] [CrossRef]
- Juillerat-Jeanneret, L. Dipeptidyl Peptidase IV and Its Inhibitors: Therapeutics for Type 2 Diabetes and What Else? J. Med. Chem. 2014, 57, 2197–2212. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Features of Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides from Dietary Proteins. J. Food Biochem. 2019, 43, e12451. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liu, X.; Li, Y.; You, H.; Yu, Z.; Wang, L.; Liu, X.; Ding, L. Identification and Characterization of Dipeptidyl Peptidase-IV Inhibitory Peptides from Oat Proteins. Foods 2022, 11, 1406. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Wu, Z.; Yi, J.; Fu, L.; Yang, Z.; Hsieh, C.; Yin, M.; Zeng, X.; Wu, C.; Lu, A. ADMETlab 2.0: An Integrated Online Platform for Accurate and Comprehensive Predictions of ADMET Properties. Nucleic Acids Res. 2021, 49, W5–W14. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.; Beaumont, K.; Maurer, T.S.; Di, L. Volume of Distribution in Drug Design: Miniperspective. J. Med. Chem. 2015, 58, 5691–5698. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, S.; Weber, J. Saxagliptin. Drugs 2009, 69, 2103–2114. [Google Scholar] [CrossRef] [PubMed]
- Galloway, I.; McKay, G.; Fisher, M. Omarigliptin. Pract. Diabetes 2017, 34, 70–71. [Google Scholar] [CrossRef]
- Lauster, C.D.; McKaveney, T.P.; Muench, S.V. Vildagliptin: A Novel Oral Therapy for Type 2 Diabetes Mellitus. Am. J. Health Syst. Pharm. 2007, 64, 1265–1273. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Hernández Álvarez, A.J.; Maycock, J.; Murray, B.S.; Boesch, C. Comparison of Alcalase- and Pepsin-Treated Oilseed Protein Hydrolysates—Experimental Validation of Predicted Antioxidant, Antihypertensive and Antidiabetic Properties. Curr. Res. Food Sci. 2021, 4, 141–149. [Google Scholar] [CrossRef]
- Michelke, L.; Deussen, A.; Dieterich, P.; Martin, M. Effects of Bioactive Peptides Encrypted in Whey-, Soy- and Rice Protein on Local and Systemic Angiotensin-Converting Enzyme Activity. J. Funct. Foods 2017, 28, 299–305. [Google Scholar] [CrossRef]
- Naha, S.; Gardner, M.J.; Khangura, D.; Kurukulasuriya, L.R.; Sowers, J.R. Hypertension in Diabetes. Eur. PMC 2015. [Google Scholar]
- Chang, Y.-W.; Alli, I.; Molina, A.T.; Konishi, Y.; Boye, J.I. Isolation and Characterization of Chickpea (Cicer arietinum L.) Seed Protein Fractions. Food Bioprocess Technol. 2012, 5, 618–625. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [Green Version]
- Arámburo-Gálvez, J.G.; Arvizu-Flores, A.A.; Cárdenas-Torres, F.I.; Cabrera-Chávez, F.; Ramírez-Torres, G.I.; Flores-Mendoza, L.K.; Gastelum-Acosta, P.E.; Figueroa-Salcido, O.G.; Ontiveros, N. Prediction of ACE-I Inhibitory Peptides Derived from Chickpea (Cicer arietinum L.): In Silico Assessments Using Simulated Enzymatic Hydrolysis, Molecular Docking and ADMET Evaluation. Foods 2022, 11, 1576. [Google Scholar] [CrossRef]
- Bell, E.W.; Zhang, Y. DockRMSD: An Open-Source Tool for Atom Mapping and RMSD Calculation of Symmetric Molecules through Graph Isomorphism. J. Cheminform. 2019, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2. 0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protein | A | B | Enzyme | AE | BE |
|---|---|---|---|---|---|
| Provicilin | 0.6093 | 0.0003526629028523 | Pepsin | 0.022 | 4.7635643240156 × 10−5 |
| Chymotrypsin | 0.0287 | 3.2429337219228 × 10−5 | |||
| Trypsin | 0.0088 | 6.8625336714504 × 10−7 | |||
| Gastrointestinal | 0.0596 | 6.7698735295912 × 10−5 | |||
| Papain | 0.0795 | 8.271490429923 × 10−5 | |||
| Ficin | 0.0728 | 6.4223543298536 × 10−5 | |||
| Stem bromelain | 0.0817 | 6.0467750947608 × 10−5 | |||
| Subtilisin (Alcalase) | 0.0662 | 0.000165417 | |||
| Legumin | 0.5847 | 0.0002483186581999 | Pepsin | 0.0141 | 5.3720267147866 × 10−6 |
| Chymotrypsin | 0.0423 | 1.302957714179 × 10−5 | |||
| Trypsin | 0.0040 | 0 | |||
| Gastrointestinal | 0.0585 | 1.8401603856577 × 10−5 | |||
| Papain | 0.0927 | 2.5172135076017 × 10−5 | |||
| Ficin | 0.0484 | 4.2932403610376 × 10−5 | |||
| Stem bromelain | 0.0867 | 0 | |||
| Subtilisin (Alcalase) | 0.0383 | 9.1420948488684 × 10−6 |
| Peptide | BIOPEP ID | Binding Energy (Kcal/Mol) | Protein | Location | Released by | Pubchem ID |
|---|---|---|---|---|---|---|
| HF | 8791 | −7.9 | Provicilin | 426–427 | Stem bromelain; Subtilisin | 152198 |
| Legumin | 473–474 | Stem bromelain; Subtilisin; Pepsin; Papain | ||||
| IW | 8807 | −7.8 | Legumin | 457–458 | Subtilisin | 7019084 |
| YF | 8935 | −7.8 | Provicilin | 95–96 | Stem bromelain | 7009600 |
| Legumin | 99–100 | Stem bromelain; Papain | ||||
| QF | 8870 | −7.7 | Provicilin | 153–154 | Gastrointestinal; Papain; Ficin | 57288566 |
| Legumin | 61–62 | Gastrointestinal; Papain; Ficin | ||||
| YL | 8940 | −7.6 | Legumin | 182–183 | Pepsin; Papain; Stem bromelain | 87071 |
| FR | 8780 | −7.5 | Legumin | 135–136 | Trypsin | 150903 |
| KF | 8809 | −7.1 | Legumin | 124–125 | Stem bromelain; Subtilisin | 151410 |
| VF | 8917 | −7.1 | Provicilin | 58–59 122–123 | Papain; Subtilisin | 6993120 |
| Legumin | 103–104 148–149 403–404 | Chymotrypsin; Gastrointestinal; Ficin; Subtilisin | ||||
| YA | 8932 | −7.1 | Legumin | 384–385 394–395 | Stem bromelain | 7020632 |
| HL | 8557 | −7 | Provicilin | 72–73 | Pepsin; Papain; Stem bromelain; Subtilisin | 189008 |
| NR | 8849 | −7 | Provicilin | 162–163 | Papain; Ficin; Stem bromelain | 14299174 |
| Legumin | 133–134 225–226 | Stem bromelain | ||||
| PH | 8856 | −7 | Provicilin | 295–296 | Chymotrypsin; Gastrointestinal; Ficin | 9856353 |
| GF | 8782 | −6.9 | Provicilin | 63–64 375–376 377–378 | Chymotrypsin; Pepsin; Gastrointestinal; Subtilisin | 92953 |
| Legumin | 212–213 312–313 | Chymotrypsin; Gastrointestinal; Subtilisin | ||||
| GY | 8788 | −6.9 | Legumin | 98–99 | Chymotrypsin; Gastrointestinal | 92829 |
| PF | 8854 | −6.9 | Provicilin | 362–363 | Chymotrypsin; Gastrointestinal; Ficin | 6351946 |
| Legumin | 471–472 | Stem bromelain | ||||
| SF | 8891 | −6.9 | Provicilin | 176–177 | Chymotrypsin; Pepsin; Gastrointestinal; Papain | 7009597 |
| Legumin | 10–11 347–348 | Chymotrypsin; Pepsin; Gastrointestinal; Papain | ||||
| TF | 8900 | −6.9 | Provicilin | 110–111 | Gastrointestinal; Ficin | 7010580 |
| IPA | 8304 | −6.7 | Provicilin | 357–359 | Stem bromelain | 10040393 |
| YV | 8946 | −6.7 | Legumin | 433–434 | Stem bromelain | 7009560 |
| DR | 8769 | −6.6 | Provicilin | 203–204 416–417 | Trypsin; Gastrointestinal; Ficin; Stem bromelain | 16122509 |
| Legumin | 440–441 429–430 | Gastrointestinal; Ficin; Stem bromelain | ||||
| IR | 8806 | −6.6 | Provicilin | 442–443 | Trypsin; Gastrointestinal; Ficin | 7021814 |
| ET | 8774 | −6.5 | Legumin | 109–110 | Stem bromelain | 6998031 |
| PT | 8863 | −6.5 | Legumin | 144–145 | Stem bromelain | 53860028 |
| IL | 8802 | −6.3 | Provicilin | 139–140 168–169 304–305 449–450 | Gastrointestinal; Papain; Ficin; Stem bromelain; Subtilisin | 7019083 |
| Legumin | 382–383 | Ficin; Stem bromelain; Subtilisin | ||||
| QP | 8532 | −6.3 | Provicilin | 431–432 | Papain | 11736661 |
| Legumin | 25–26 | |||||
| VE | 8916 | −6.3 | Provicilin | 56–57 | Subtilisin | 7009623 |
| VN | 8924 | −6.3 | Legumin | 132–133 224–225 | Chymotrypsin; Gastrointestinal | 7020201 |
| VQ | 8925 | −6.3 | Provicilin | 340–341 | Subtilisin | 7016045 |
| EG | 8770 | −6.1 | Legumin | 48–49 121–122 137–138 | Papain; Ficin; Stem bromelain | 6427052 |
| ES | 8773 | −6.1 | Provicilin | 244–245 | Ficin; Subtilisin; Stem bromelain | 6995653 |
| Legumin | 116–117 | Ficin; Stem bromelain | ||||
| HP | 8520 | −6 | Legumin | 477–478 | Papain | 152322 |
| VL | 8922 | −6 | Provicilin | 92–93 101–102 104–105 124–125 186–187 | Chymotrypsin; Pepsin; Gastrointestinal; Papain; Ficin; Subtilisin | 6993117 |
| IN | 8804 | −5.9 | Provicilin | 378–379 | Papain | 7016080 |
| Legumin | 444–445 | Chymotrypsin; Gastrointestinal; Subtilisin | ||||
| KG | 8810 | −5.9 | Provicilin | 311–312 | Papain; Stem bromelain | 7022320 |
| Legumin | 248–249 387–388 | Papain; Stem bromelain | ||||
| PK | 8858 | −5.9 | Provicilin | 195–196 | Trypsin; Gastrointestinal; Ficin | 9209431 |
| SL | 8560 | −5.9 | Provicilin | 6–7 61–62 128–129 291–292 | Chymotrypsin; Pepsin; Gastrointestinal; Papain | 7015694 |
| Legumin | 8–9 78–79 364–365 426–427 | Chymotrypsin; Pepsin; Gastrointestinal; Papain | ||||
| VI | 8920 | −5.9 | Provicilin | 217–218 | Subtilisin | 7010531 |
| VS | 8926 | −5.9 | Provicilin | 37–38 67–68 241–242 | Ficin; Subtilisin | 6992640 |
| Legumin | 162–163 | Subtilisin | ||||
| IA | 8525 | −5.8 | Provicilin | 69–70 166–167 | Stem bromelain | 7009577 |
| Legumin | 160–161 422–423 | |||||
| VK | 8921 | −5.8 | Legumin | 245–246 247–248 | Trypsin; Gastrointestinal; Ficin; Subtilisin | 168058 |
| VT | 8927 | −5.8 | Provicilin | 120–121 | Subtilisin | 9815826 |
| Legumin | 345–346 | Papain | ||||
| GL | 8561 | −5.7 | Provicilin | 74–75 | Chymotrypsin; Pepsin; Gastrointestinal; Subtilisin | 1548899 |
| SK | 8894 | −5.7 | Provicilin | 230–231 | Gastrointestinal | 16122513 |
| PG | 8855 | −5.6 | Provicilin | 409–410 | Papain; Ficin; Stem bromelain | 6426709 |
| Legumin | 105–106 463–464 | Papain; Ficin; Stem bromelain | ||||
| VG | 8918 | −5.5 | Provicilin | 317–318 | Papain; Ficin | 6993110 |
| MA | 3173 | −5.2 | Legumin | 1–2 | Stem bromelain | 7009581 |
| Peptide/Drug | Lipinski Rules | HIA | F 20% | F 30% | VD (L/kg) | T 1/2 (h) | ROAT |
|---|---|---|---|---|---|---|---|
| Omarigliptin | Accepted |  |  |  |  | 0.151 |  |
| Saxagliptin | Accepted |  |  |  |  | 0.309 |  |
| Vildagliptin | Accepted |  |  |  |  | 0.37 |  |
| HF | Accepted |  |  |  |  | 0.92 |  |
| IW | Accepted |  |  |  |  | 0.91 |  |
| YF | Accepted |  |  |  |  | 0.921 |  |
| QF | Accepted |  |  |  |  | 0.61 |  |
| YL | Accepted |  |  |  |  | 0.909 |  |
| FR | Accepted |  |  |  |  | 0.826 |  |
| KF | Accepted |  |  |  |  | 0.835 |  |
| VF | Accepted |  |  |  |  | 0.872 |  |
| YA | Accepted |  |  |  |  | 0.894 |  |
| HL | Accepted |  |  |  |  | 0.919 |  |
| NR | Accepted |  |  |  |  | 0.422 |  |
| PH | Accepted |  |  |  |  | 0.887 |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mora-Melgem, J.A.; Arámburo-Gálvez, J.G.; Cárdenas-Torres, F.I.; Gonzalez-Santamaria, J.; Ramírez-Torres, G.I.; Arvizu-Flores, A.A.; Figueroa-Salcido, O.G.; Ontiveros, N. Dipeptidyl Peptidase IV Inhibitory Peptides from Chickpea Proteins (Cicer arietinum L.): Pharmacokinetics, Molecular Interactions, and Multi-Bioactivities. Pharmaceuticals 2023, 16, 1109. https://doi.org/10.3390/ph16081109
Mora-Melgem JA, Arámburo-Gálvez JG, Cárdenas-Torres FI, Gonzalez-Santamaria J, Ramírez-Torres GI, Arvizu-Flores AA, Figueroa-Salcido OG, Ontiveros N. Dipeptidyl Peptidase IV Inhibitory Peptides from Chickpea Proteins (Cicer arietinum L.): Pharmacokinetics, Molecular Interactions, and Multi-Bioactivities. Pharmaceuticals. 2023; 16(8):1109. https://doi.org/10.3390/ph16081109
Chicago/Turabian StyleMora-Melgem, José Antonio, Jesús Gilberto Arámburo-Gálvez, Feliznando Isidro Cárdenas-Torres, Jhonatan Gonzalez-Santamaria, Giovanni Isaí Ramírez-Torres, Aldo Alejandro Arvizu-Flores, Oscar Gerardo Figueroa-Salcido, and Noé Ontiveros. 2023. "Dipeptidyl Peptidase IV Inhibitory Peptides from Chickpea Proteins (Cicer arietinum L.): Pharmacokinetics, Molecular Interactions, and Multi-Bioactivities" Pharmaceuticals 16, no. 8: 1109. https://doi.org/10.3390/ph16081109