
Scaling the Andean Shilajit: A Novel Neuroprotective Agent for Alzheimer’s Disease
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
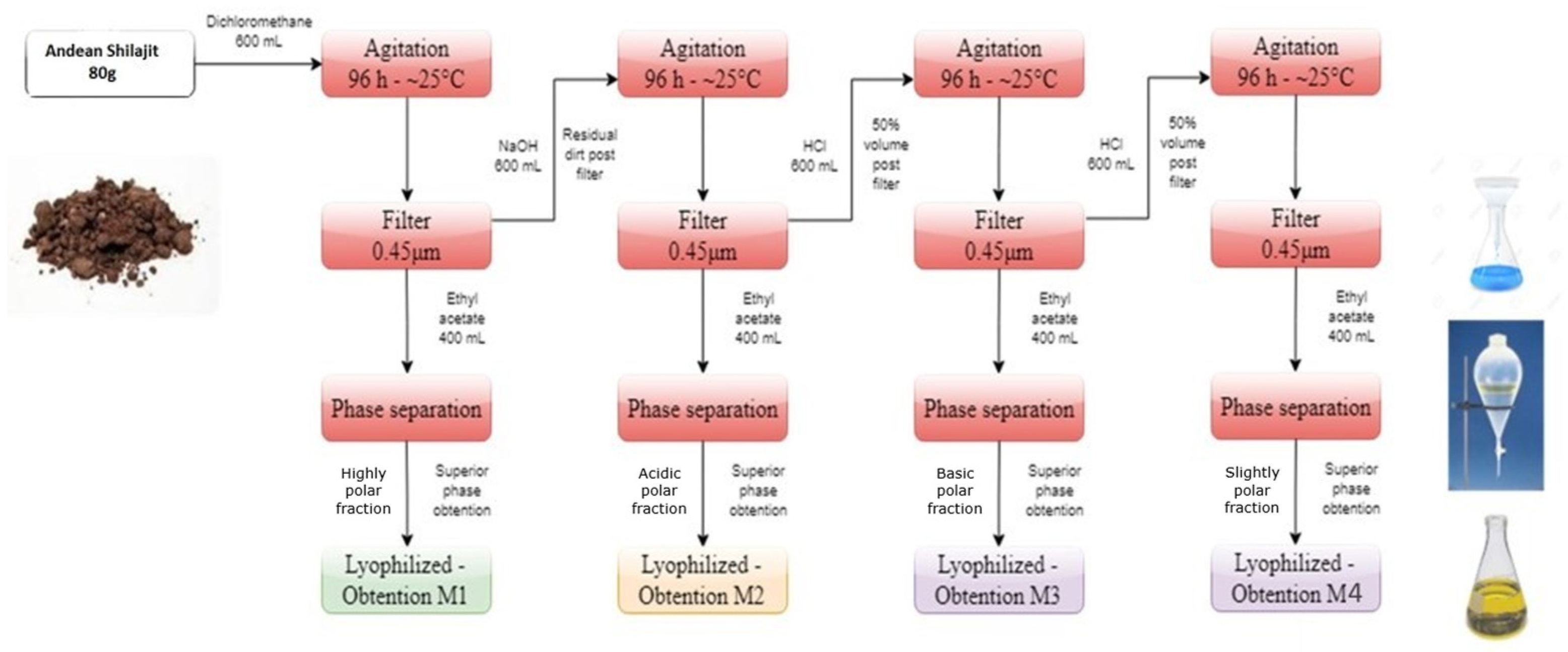
2.1. Chemical Investigation of the Andean Shilajit
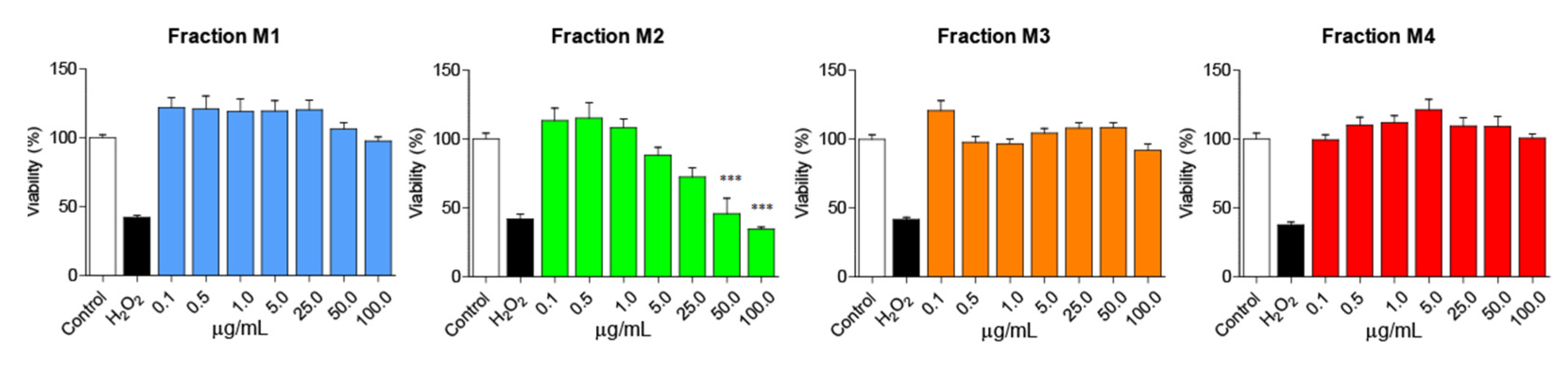
2.2. Andean Shilajit Fractions Are Not Toxic in N2a Cells
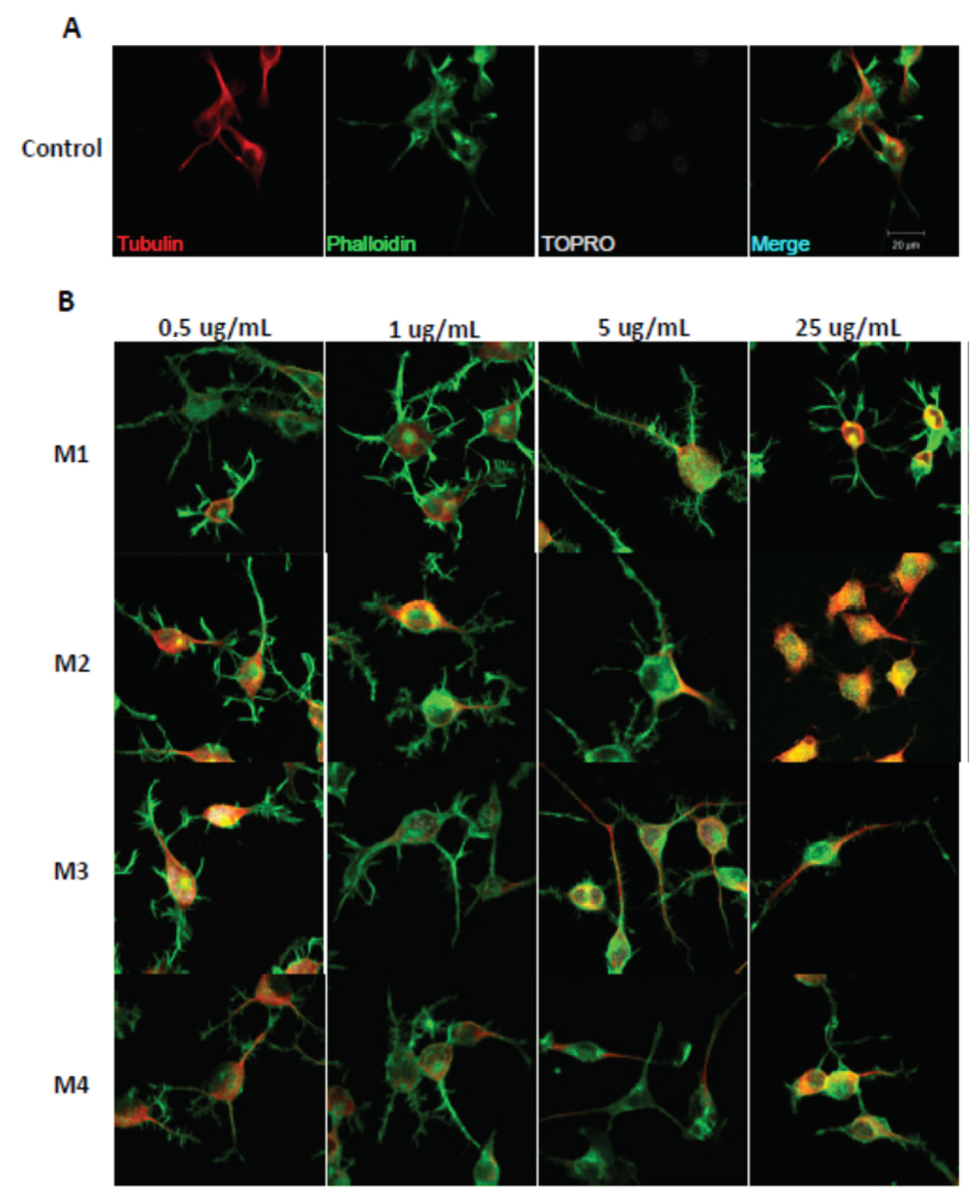
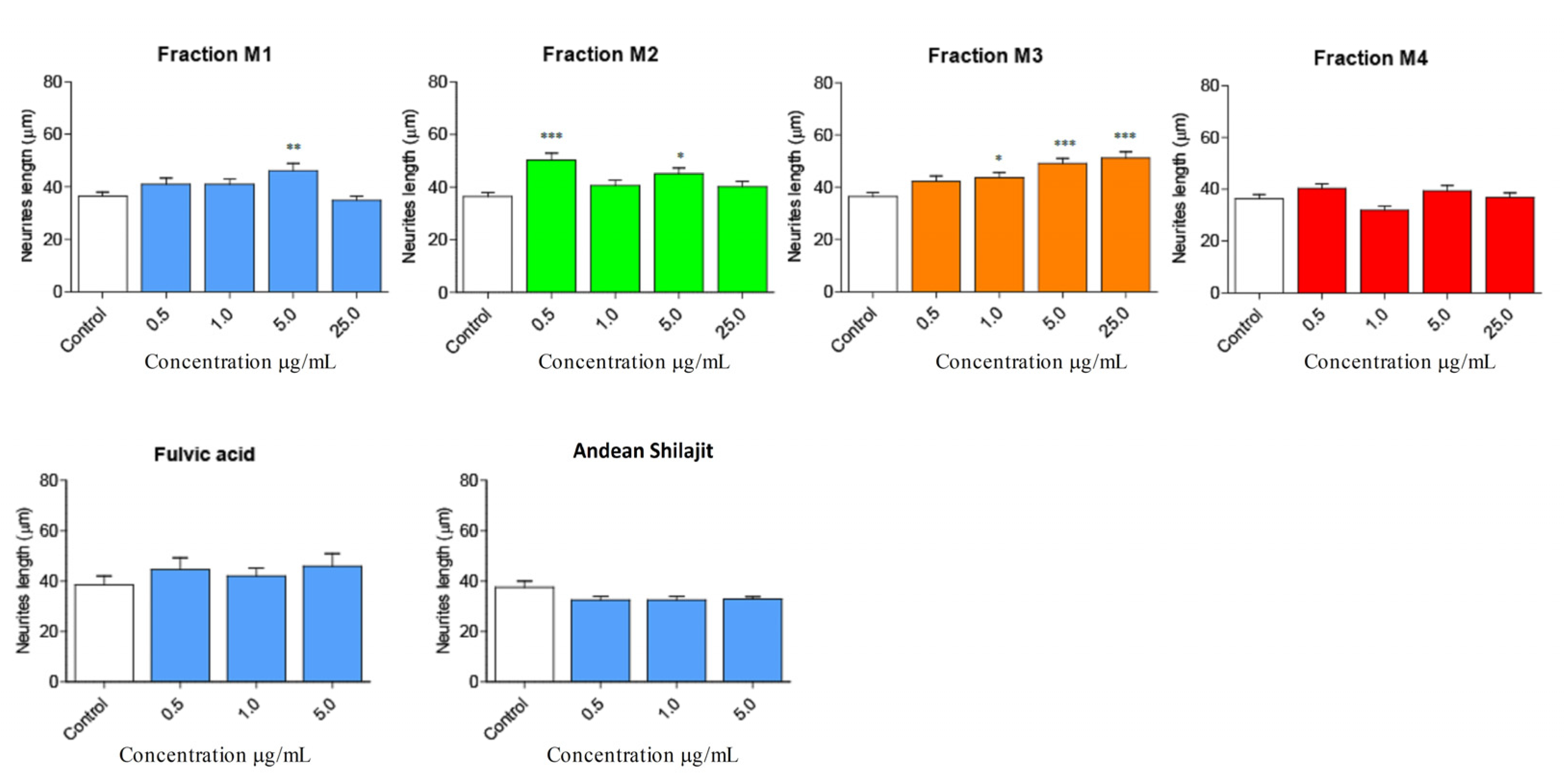
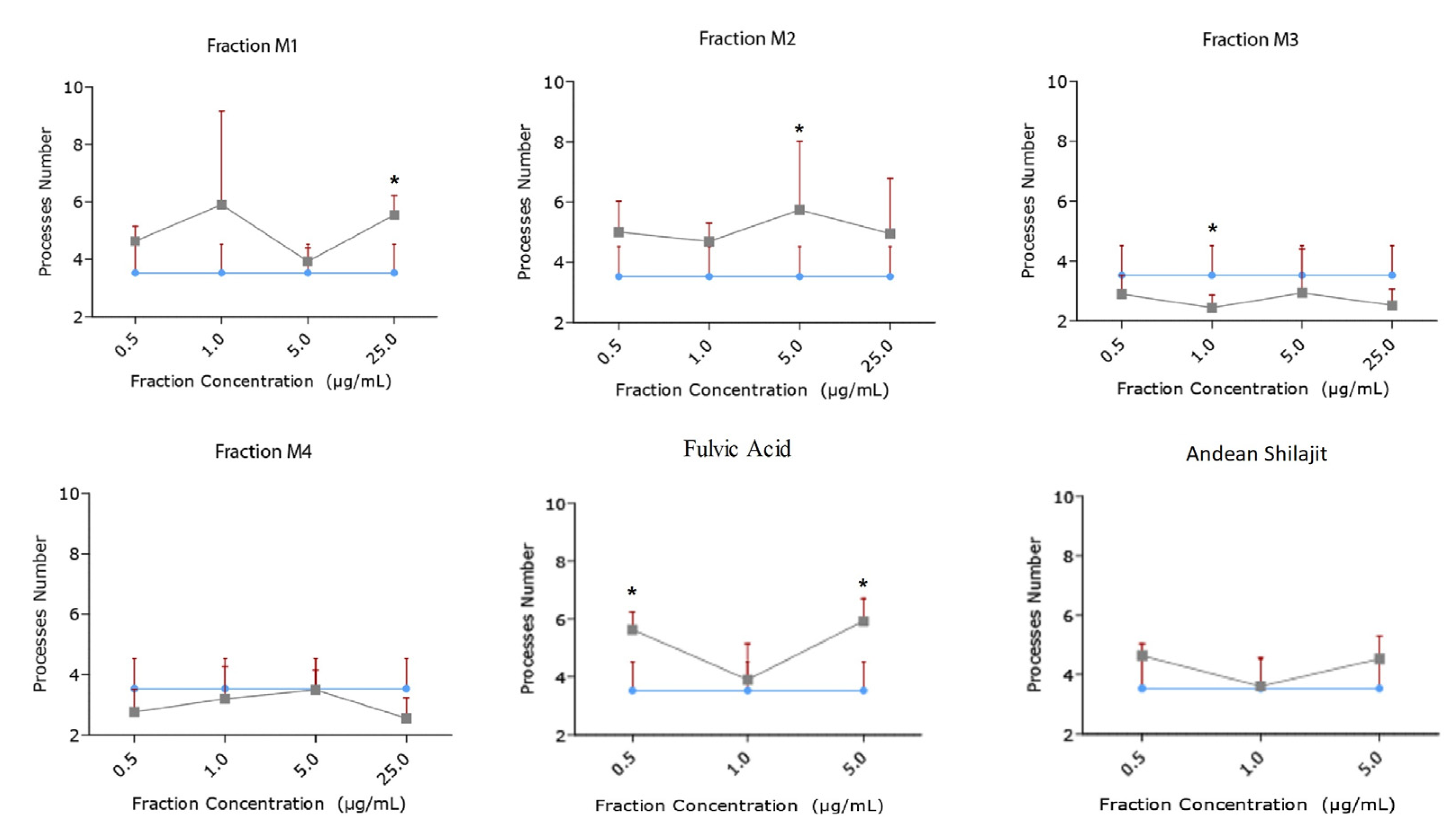
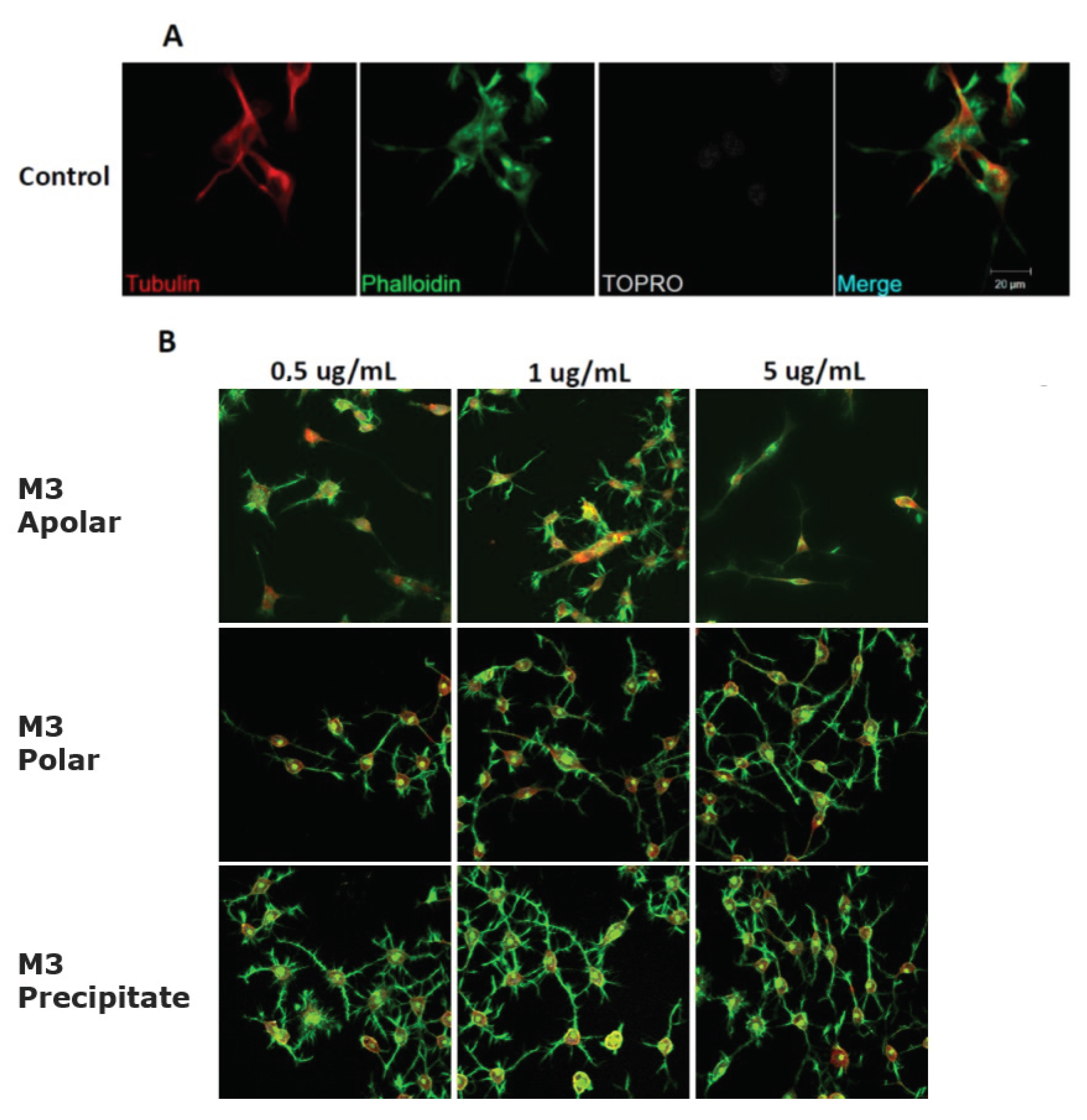
2.3. Andean Shilajit Fractions Increase Neuritogenesis and the Number of Neuronal Processes in N2a Cells
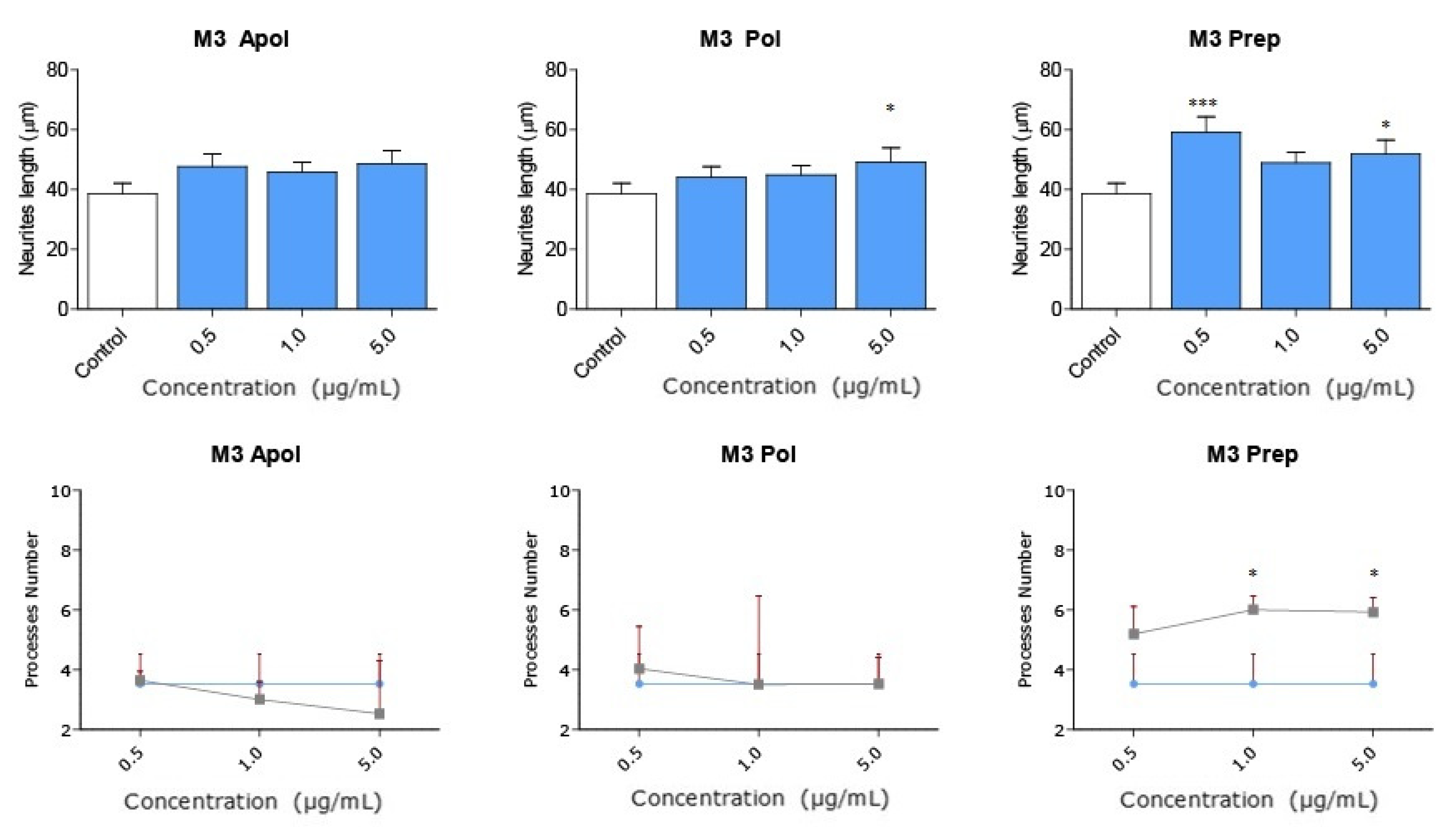
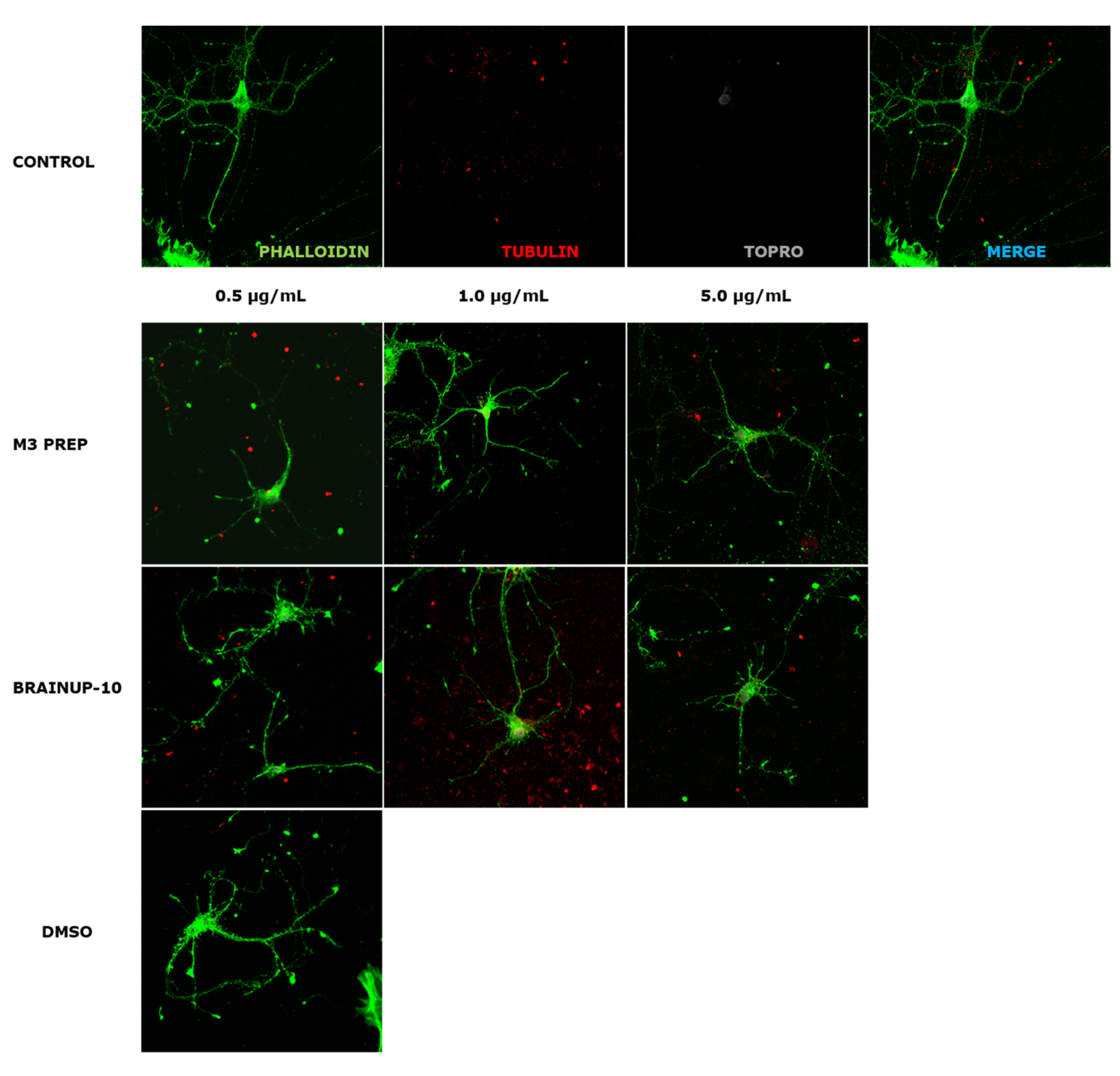
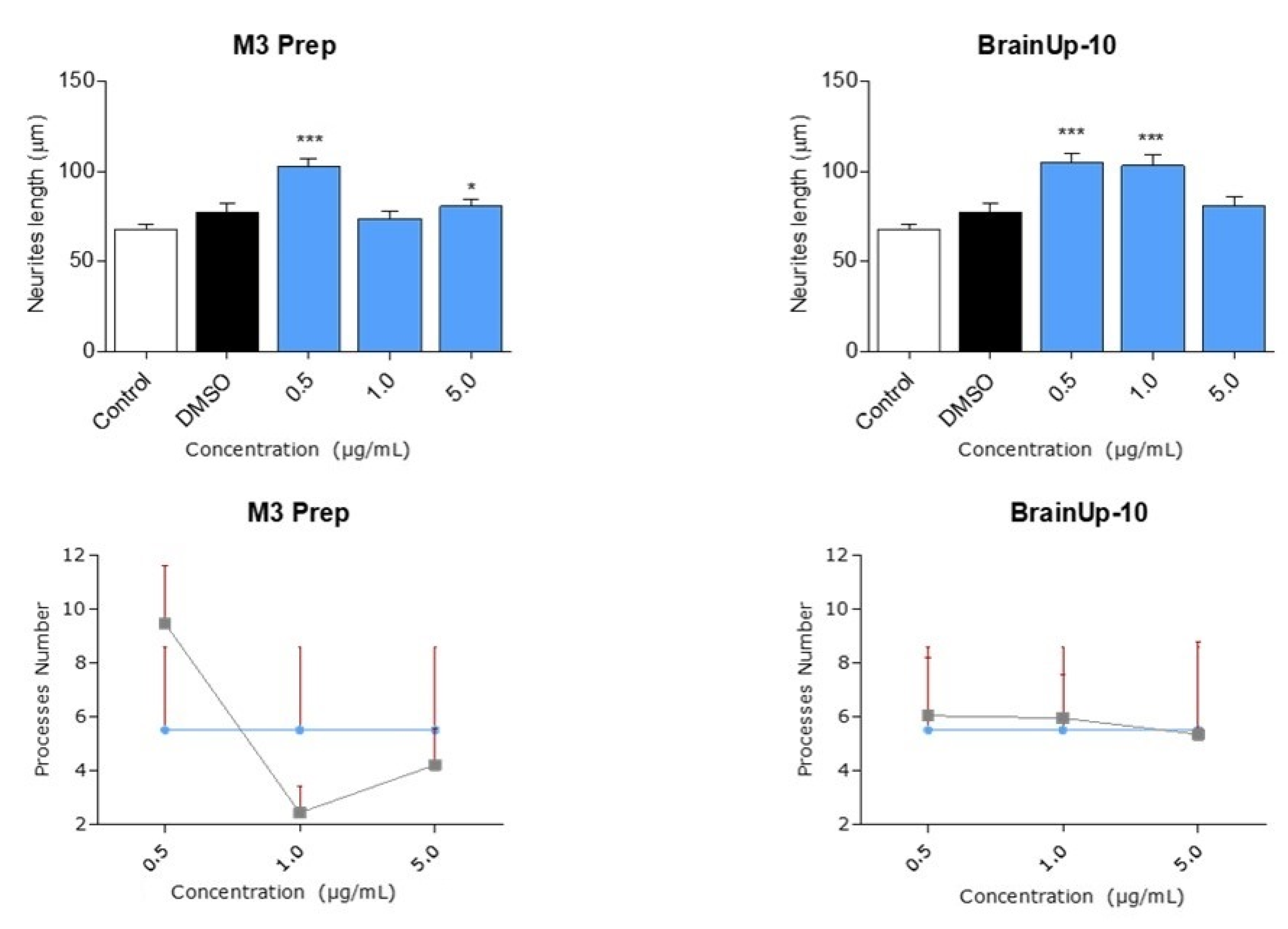
2.4. The Precipitate M3 Subfraction and BrainUp-10 ® Increase Neuritogenesis and the Number of Processes in Rat Hippocampal Neurons (RHN)
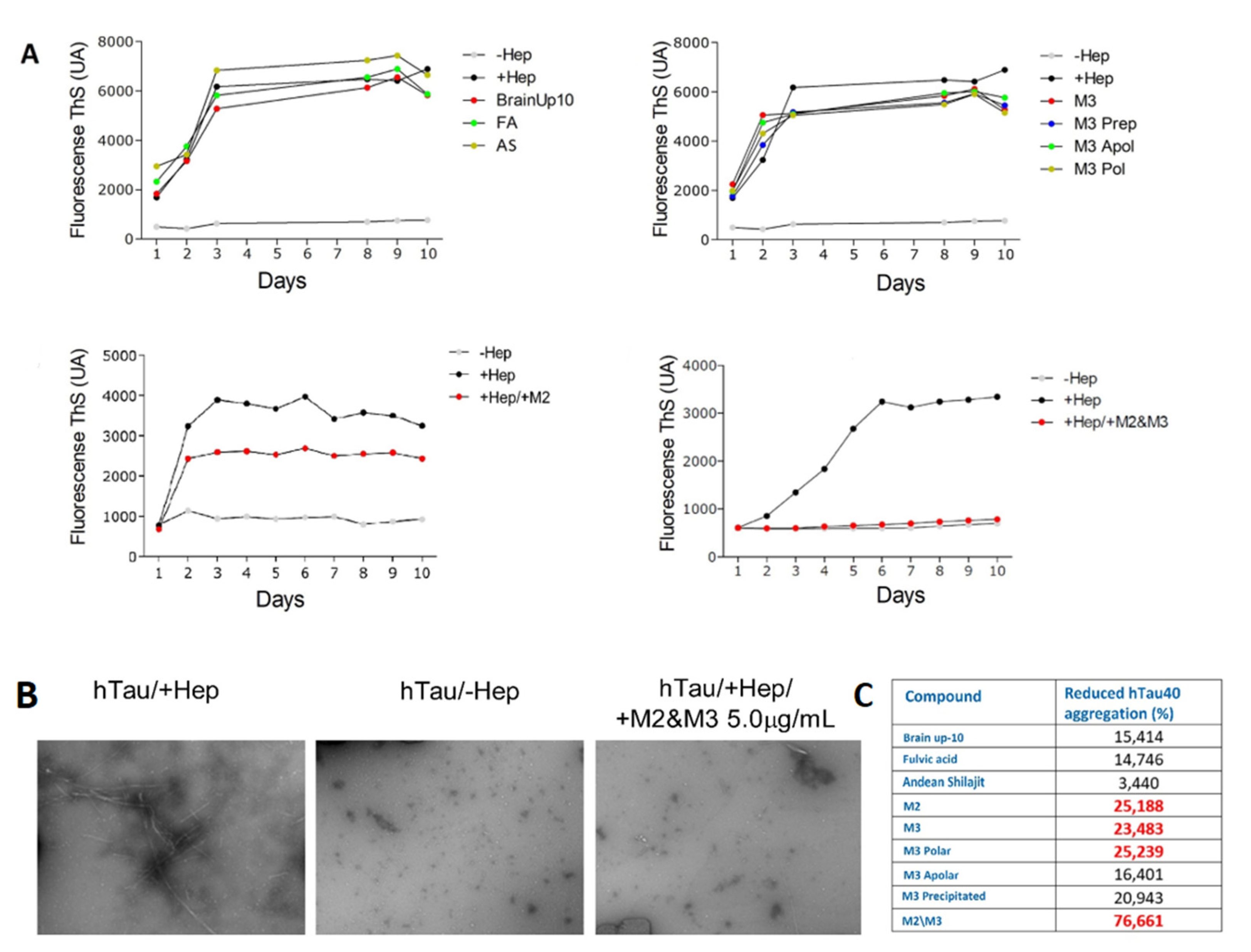
2.5. AnSh Fractions Inhibit Human Tau Aggregation
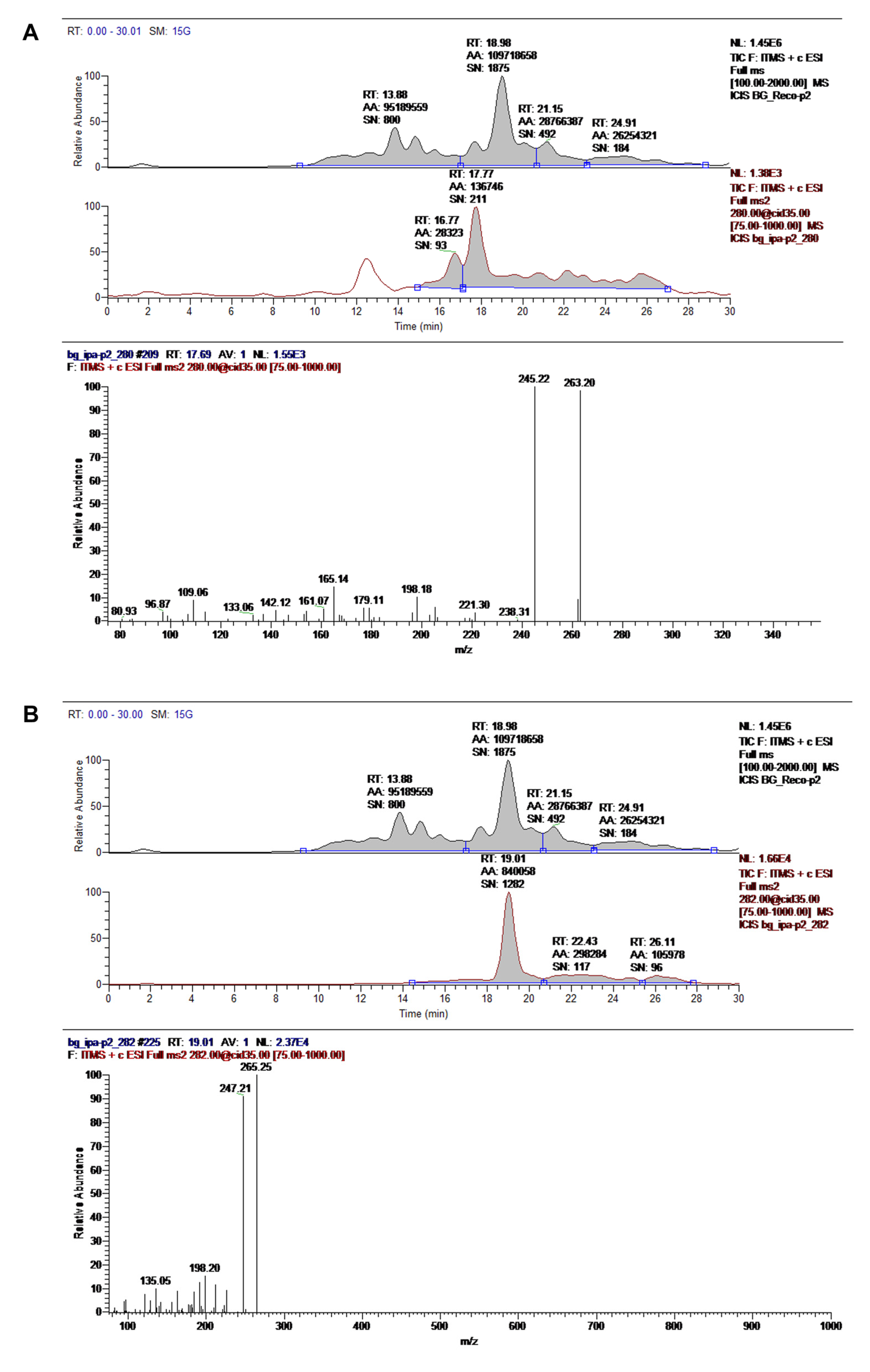
2.6. Characterization of Two Main Candidates by UPLC/MS
3. Discussion
4. Materials and Methods
4.1. Andean Shilajit
4.2. Liquid Chromatography–Electrospray Ionization Mass Spectrometry (LC-ESI–MS)
4.3. Mouse Neuroblastoma Cells
4.4. Primary Cultures of Rat Hippocampal Neurons (RHN)
4.5. Viability Assay
4.6. Immunofluorescence
4.7. Tau Protein Purification
4.8. Tau Aggregation Assays
4.9. ThS Fluorescence Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bettens, K.; Sleegers, K.; Van Broeckhoven, C. Current status on Alzheimer disease molecular genetics: From past, to present, to future. Hum. Mol. Genet. 2010, 19, R4–R11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, M.J.; Wimo, A.; Guerchet, M.M.; Ali, G.C.; Wu, Y.T.; Prina, M. World Alzheimer Report 2015 The Global Impact of Dementia an Analysis of Prevalence, Incidence, Cost and Trends; Alzheimer’s Disease International (ADI): London, UK, 2015. [Google Scholar]
- Alzheimer’s Disease International; Guerchet, M.; Prince, M.; Prina, M. Numbers of People with Dementia Worldwide; Alzheimer’s Disease International: London, UK, 2020. [Google Scholar]
- Andrade, V.; Cortes, N.; Guzman-Martinez, L.; Maccioni, R.B. An Overview of the Links between Behavioral Disorders and Alzheimer’s Disease. JSM Alzheimer Dis. Relat. Dement. 2017, 4, 1031. [Google Scholar]
- Maccioni, R.B. Introductory remarks. Molecular, biological and clinical aspects of Alzheimer’s disease. Arch. Med. Res. 2012, 43, 593–594. [Google Scholar] [CrossRef] [PubMed]
- Maccioni, R.B.; Munoz, J.P.; Barbeito, L. The molecular bases of Alzheimer’s disease and other neurodegenerative disorders. Arch. Med. Res. 2001, 32, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Farias, G.; Cornejo, A.; Jimenez, J.; Guzman, L.; Maccioni, R.B. Mechanisms of tau self-aggregation and neurotoxicity. Curr. Alzheimer Res. 2011, 8, 608–614. [Google Scholar] [CrossRef]
- Guzman-Martinez, L.; Farias, G.A.; Maccioni, R.B. Tau Oligomers as Potential Targets for Alzheimer’s Diagnosis and Novel Drugs. Front. Neurol. 2013, 4, 167. [Google Scholar] [CrossRef] [Green Version]
- Lambert, J.C.; Schraen-Maschke, S.; Richard, F.; Fievet, N.; Rouaud, O.; Berr, C.; Dartigues, J.F.; Tzourio, C.; Alperovitch, A.; Buee, L.; et al. Association of plasma amyloid beta with risk of dementia: The prospective Three-City Study. Neurology 2009, 73, 847–853. [Google Scholar] [CrossRef]
- Maccioni, R.B.; Rojo, L.E.; Fernández, J.A.; Kuljis, R.O. The role of neuroimmunomodulation in Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2009, 1153, 240–246. [Google Scholar] [CrossRef]
- Fernández, J.A.; Rojo, L.; Kuljis, R.O.; Maccioni, R.B. The damage signals hypothesis of Alzheimer’s disease pathogenesis. J. Alzheimer’s Dis. 2008, 14, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-Gallardo, C.; Farías, G.A.; Fuentes, P.; Crespo, F.; Maccioni, R.B. Can nutraceuticals prevent Alzheimer’s disease? Potential therapeutic role of a formulation containing shilajit and complex B vitamins. Arch. Med. Res. 2012, 43, 699–704. [Google Scholar] [CrossRef]
- Fenech, M. Vitamins Associated with Brain Aging, Mild Cognitive Impairment, and Alzheimer Disease: Biomarkers, Epidemiological and Experimental Evidence, Plausible Mechanisms, and Knowledge Gaps. Adv. Nutr. 2017, 8, 958–970. [Google Scholar] [CrossRef] [Green Version]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Kim, M.O.; Ali, T. Editorial: Natural Products-Based Drugs: Potential Therapeutics Against Alzheimer’s Disease and Other Neurological Disorders. Front. Pharmacol. 2019, 10, 1417. [Google Scholar] [CrossRef] [Green Version]
- Calfio, C.; Gonzalez, A.; Singh, S.; Rojo, L.; Maccioni, R. The Emerging Role of Nutraceuticals and Phytochemicals in the Prevention and Treatment of Alzheimer’s Disease. J. Alzheimer Dis. 2020, 77, 1–19. [Google Scholar] [CrossRef]
- Islam, F.; Khadija, J.F.; Harun-Or-Rashid, M.; Rahaman, M.S.; Nafady, M.H.; Islam, M.R.; Akter, A.; Emran, T.B.; Wilairatana, P.; Mubarak, M.S. Bioactive Compounds and Their Derivatives: An Insight into Prospective Phytotherapeutic Approach against Alzheimer’s Disease. Oxid. Med. Cell. Longev. 2022, 2022, 5100904. [Google Scholar] [CrossRef]
- Nelson, A.; Karageorgis, G. Natural product-informed exploration of chemical space to enable bioactive molecular discovery. RSC Med. Chem. 2021, 12, 353–362. [Google Scholar] [CrossRef]
- Bolognesi, M.L. Polypharmacology in a single drug: Multitarget drugs. Curr. Med. Chem. 2013, 20, 1639–1645. [Google Scholar] [CrossRef]
- Maccioni, R.B.; Calfío, C.; González, A.; Lüttges, V. Novel Nutraceutical Compounds in Alzheimer Prevention. Biomolecules 2022, 12, 249. [Google Scholar] [CrossRef]
- Kola, A.; Dudek, D.; Valensin, D. Metal Complexation Mechanisms of Polyphenols Associated to Alzheimer’s Disease. Curr. Med. Chem. 2021, 28, 7278–7294. [Google Scholar] [CrossRef]
- Lakey-Beitia, J.; Burillo, A.M.; La Penna, G.; Hegde, M.L.; Rao, K.S. Polyphenols as Potential Metal Chelation Compounds Against Alzheimer’s Disease. J. Alzheimer Dis. 2021, 82, S335–S357. [Google Scholar] [CrossRef]
- Carrasco-Gallardo, C.; Guzmán, L.; Maccioni, R.B. Shilajit: A natural phytocomplex with potential procognitive activity. Int. J. Alzheimer Dis. 2012, 2012, 674142. [Google Scholar] [CrossRef] [Green Version]
- Maccioni, B.R.; Sepúlveda, L.Q.; Saavedra, I.S.; Salas, R.S. Nutraceutical Composition That Comprises Extract of Shilajit, Folic Acid, Vitamin b12 and Vitamin b6 and the Use Thereof for Preventing and/or Treating Neurodegenerative Diseases and/or the Cognitive Deterioration Associated with Cerebral Ageing Patents WO2011041920A2, 8 October 2010.
- Guzman-Martinez, L.; Farías, G.A.; Tapia, J.P.; Sánchez, M.P.; Fuentes, P.; Gloger, S.; Maccioni, R.B. Interventional Study to Evaluate the Clinical Effects and Safety of the Nutraceutical Compound BrainUp-10® in a Cohort of Patients with Alzheimer’s Disease: A Multicenter, Randomized, Double-Blind, and Placebo-Controlled Trial. J. Alzheimer Dis. 2021, 81, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Lledo, P.-M.; Alonso, M.; Grubb, M.S. Adult neurogenesis and functional plasticity in neuronal circuits. Nat. Rev. Neurosci. 2006, 7, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.J.; Zhou, Y.; Rogerson, T.; Shobe, J.; Balaji, J. Molecular and cellular approaches to memory allocation in neural circuits. Science 2009, 326, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, A.; Fujii, Y.; Kasai, N.; Okajima, T.; Nakashima, H. Regulation of neuritogenesis in hippocampal neurons using stiffness of extracellular microenvironment. PLoS ONE 2018, 13, e0191928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gertz, C.C.; Leach, M.K.; Birrell, L.K.; Martin, D.C.; Feldman, E.L.; Corey, J.M. Accelerated neuritogenesis and maturation of primary spinal motor neurons in response to nanofibers. Dev. Neurobiol. 2010, 70, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Gil-Mohapel, J.; Brocardo, P.S.; Choquette, W.; Gothard, R.; Simpson, J.M.; Christie, B.R. Hippocampal neurogenesis levels predict WATERMAZE search strategies in the aging brain. PLoS ONE 2013, 8, e75125. [Google Scholar] [CrossRef] [Green Version]
- Horgusluoglu, E.; Nudelman, K.; Nho, K.; Saykin, A.J. Adult neurogenesis and neurodegenerative diseases: A systems biology perspective. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. Off. Publ. Int. Soc. Psychiatr. Genet. 2017, 174, 93–112. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.P.; Khanna, R.; Karmarkar, R.; Anwer, M.K.; Khar, R.K. Shilajit: A review. Phytother. Res. 2007, 21, 401–405. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Khlebnikov, A.I.; Ah, S.Y.; Woo, S.B.; Jeong, C.S.; Klubachuk, O.N.; Kwon, B.S. Characterization and biological activities of humic substances from mumie. J. Agric. Food Chem. 2003, 51, 5245–5254. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Chen, H.Z. AChE inhibition-based multi-target-directed ligands, a novel pharmacological approach for the symptomatic and disease-modifying therapy of Alzheimer’s disease. Curr. Neuropharmacol. 2016, 14, 364–375. [Google Scholar] [CrossRef] [Green Version]
- Citron, M. Alzheimer’s disease: Strategies for disease modification. Nat. Rev. Drug Discov. 2010, 9, 387–398. [Google Scholar] [CrossRef]
- Leach, J.B.; Brown, X.Q.; Jacot, J.G.; DiMilla, P.A.; Wong, J.Y. Neurite outgrowth and branching of PC12 cells on very soft substrates sharply decreases below a threshold of substrate rigidity. J. Neural Eng. 2007, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Cornejo, A.; Jimenez, J.M.; Caballero, L.; Melo, F.; Maccioni, R.B. Fulvic acid inhibits aggregation and promotes disassembly of tau fibrils associated with Alzheimer’s disease. J. Alzheimer’s Dis. 2011, 27, 143–153. [Google Scholar] [CrossRef]
- Winkler, J.; Ghosh, S. Therapeutic Potential of Fulvic Acid in Chronic Inflammatory Diseases and Diabetes. J. Diabetes Res. 2018, 2018, 5391014. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, N.; Coballase-Urrutia, E.; Gertrudis, B.; Chaverri, J.; Barragan, G. Antioxidant activity of fulvic acid: A living matter-derived bioactive compound. J. Food Agric. Environ. 2011, 9, 123–127. [Google Scholar]
- Chong, F.P.; Ng, K.Y.; Koh, R.Y.; Chye, S.M. Tau Proteins and Tauopathies in Alzheimer’s Disease. Cell. Mol. Neurobiol. 2018, 38, 965–980. [Google Scholar] [CrossRef]
- Congdon, E.E.; Sigurdsson, E.M. Tau-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 399–415. [Google Scholar] [CrossRef]
- Boutajangout, A.; Wisniewski, T. Tau-based therapeutic approaches for Alzheimer’s disease—A mini-review. Gerontology 2014, 60, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Lindwall, G.; Cole, R.D. Phosphorylation affects the ability of tau protein to promote microtubule assembly. J. Biol. Chem. 1984, 259, 5301–5305. [Google Scholar] [CrossRef]
- Iqbal, K.; Liu, F.; Gong, C.-X. Tau and neurodegenerative disease: The story so far. Nat. Rev. Neurol. 2016, 12, 15–27. [Google Scholar] [CrossRef]
- De-Paula, V.J.; Radanovic, M.; Diniz, B.S.; Forlenza, O.V. Alzheimer’s disease. Sub-Cell. Biochem. 2012, 65, 329–352. [Google Scholar]
- Andrade, V.; Guzmán-Martínez, L.; Vergara-Pulgar, K.; Maccioni, R.B. Neuroimmune Dynamics in Alzheime’s Disease Progression. In Mechanisms of Neuroinflammation; IntectOpen: London, UK, 2017. [Google Scholar]
- Rojo, L.E.; Fernández, J.A.; Maccioni, A.A.; Jimenez, J.M.; Maccioni, R.B. Neuroinflammation: Implications for the pathogenesis and molecular diagnosis of Alzheimer’s disease. Arch. Med. Res. 2008, 39, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Santa-María, I.; Pérez, M.; Hernández, F.; Avila, J.; Moreno, F.J. Characteristics of the binding of thioflavin S to tau paired helical filaments. J. Alzheimer’s Dis. 2006, 9, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Friedhoff, P.; Schneider, A.; Mandelkow, E.-M.; Mandelkow, E. Rapid Assembly of Alzheimer-like Paired Helical Filaments from Microtubule-Associated Protein Tau Monitored by Fluorescence in Solution†. Biochemistry 1998, 37, 10223–10230. [Google Scholar] [CrossRef] [PubMed]
- Fowler, C.J. Oleamide: A member of the endocannabinoid family? Br. J. Pharmacol. 2004, 141, 195–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leggett, J.D.; Aspley, S.; Beckett, S.; D’antona, A.; Kendall, D. Oleamide is a selective endogenous agonist of rat and human CB1 cannabinoid receptors. Br. J. Pharmacol. 2004, 141, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Cravatt, B.F.; Prospero-Garcia, O.; Siuzdak, G.; Gilula, N.B.; Henriksen, S.J.; Boger, D.L.; Lerner, R.A. Chemical characterization of a family of brain lipids that induce sleep. Science 1995, 268, 1506–1509. [Google Scholar] [CrossRef] [Green Version]
- Akanmu, M.A.; Adeosun, S.O.; Ilesanmi, O.R. Neuropharmacological effects of oleamide in male and female mice. Behav. Brain Res. 2007, 182, 88–94. [Google Scholar] [CrossRef]
- Murillo-Rodríguez, E.; Giordano, M.; Cabeza, R.; Henriksen, S.J.; Díaz, M.M.; Navarro, L.; Prospéro-García, O. Oleamide modulates memory in rats. Neurosci. Lett. 2001, 313, 61–64. [Google Scholar] [CrossRef]
- Nam, H.Y.; Na, E.J.; Lee, E.; Kwon, Y.; Kim, H.J. Antiepileptic and Neuroprotective Effects of Oleamide in Rat Striatum on Kainate-Induced Behavioral Seizure and Excitotoxic Damage via Calpain Inhibition. Front. Pharmacol. 2017, 8, 817. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.F.; Li, C.L.; Song, H.R.; Zhang, H.F.; Yang, J.Y.; Wang, Y.L. Selective effect of oleamide, an endogenous sleepinducing lipid amide, on pentylenetetrazole-induced seizures in mice. J. Pharm. Pharmacol. 2003, 55, 1159–1162. [Google Scholar] [CrossRef]
- Ano, Y.; Ozawa, M.; Kutsukake, T.; Sugiyama, S.; Uchida, K.; Yoshida, A.; Nakayama, H. Preventive effects of a fermented dairy product against Alzheimer’s disease and identification of a novel oleamide with enhanced microglial phagocytosis and anti-inflammatory activity. PLoS ONE 2015, 10, e0118512. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Hamajima, N.; Ihara, Y. Neurofibrillary tangle-associated collapsin response mediator protein-2 (CRMP-2) is highly phosphorylated on Thr-509, Ser-518, and Ser-522. Biochemistry 2000, 39, 4267–4275. [Google Scholar] [CrossRef]
- Yoshida, H.; Watanabe, A.; Ihara, Y. Collapsin response mediator protein-2 is associated with neurofibrillary tangles in Alzheimer’s disease. J. Biol. Chem. 1998, 273, 9761–9768. [Google Scholar] [CrossRef] [Green Version]
- Gudasheva, T.A.; Ostrovskaya, R.U.; Seredenin, S.B. Novel Technologies for Dipeptide Drugs Design and their Implantation. Curr. Pharm. Des. 2018, 24, 3020–3027. [Google Scholar] [CrossRef]
- Kaneko, J.; Enya, A.; Enomoto, K.; Ding, Q.; Hisatsune, T. Anserine (beta-alanyl-3-methyl-L-histidine) improves neurovascular-unit dysfunction and spatial memory in aged AβPPswe/PSEN1dE9 Alzheimer’s-model mice. Sci. Rep. 2017, 7, 12571. [Google Scholar] [CrossRef]
- Ostrovskaya, R.; Tsaplina, A.; Gudasheva, T. The using perspective of dipeptide drug Noopept for cognitive deficit. Psikhiatriya 2009, 38, 30–37. [Google Scholar]
- Gazme, B.; Boachie, R.T.; Tsopmo, A.; Udenigwe, C.C. Occurrence, properties and biological significance of pyroglutamyl peptides derived from different food sources. Food Sci. Hum. Wellness 2019, 8, 268–274. [Google Scholar] [CrossRef]
- Li, Y.; Duffy, K.B.; Ottinger, M.A.; Ray, B.; Bailey, J.A.; Holloway, H.W.; Tweedie, D.; Perry, T.; Mattson, M.P.; Kapogiannis, D.; et al. GLP-1 receptor stimulation reduces amyloid-beta peptide accumulation and cytotoxicity in cellular and animal models of Alzheimer’s disease. J. Alzheimer’s Dis. 2010, 19, 1205–1219. [Google Scholar] [CrossRef] [Green Version]
- Bae, C.S.; Song, J. The Role of Glucagon-Like Peptide 1 (GLP1) in Type 3 Diabetes: GLP-1 Controls Insulin Resistance, Neuroinflammation and Neurogenesis in the Brain. Int. J. Mol. Sci. 2017, 18, 2493. [Google Scholar] [CrossRef] [Green Version]
- Avila, M.E.; Sepúlveda, F.J.; Burgos, C.F.; Moraga-Cid, G.; Parodi, J.; Moon, R.T.; Aguayo, L.G.; Opazo, C.; De Ferrari, G.V. Canonical Wnt3a modulates intracellular calcium and enhances excitatory neurotransmission in hippocampal neurons. J. Biol. Chem. 2010, 285, 18939–18947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Barghorn, S.; Biernat, J.; Mandelkow, E. Purification of Recombinant Tau Protein and Preparation of Alzheimer-Paired Helical Filaments In Vitro. In Methods in Molecular Biology; Human Press: Totowa, NJ, USA, 2005; pp. 35–51. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound/Fraction | Neurite Length Variation (%) | Neurite Number Variation (%) | ||||
|---|---|---|---|---|---|---|
| 0.5 µg/mL | 1.0 µg/mL | 5.0 µg/mL | 0.5 µg/mL | 1.0 µg/mL | 5.0 µg/mL | |
| M1 | 12.986 | 13.097 | 27.252 ** | 34.493 | 71.125 * | 13.849 |
| M2 | 38.536 * | 11.781 | 24.360 | 42.047 | 33.258 | 62.826 * |
| M3 | 16.240 | 20.424 | 35.233 * | −17.777 | −30.791 | −16.643 |
| Polar M3 | 21.470 | 23.782 | 35.728 * | 14.545 | −0.657 | 0.022 |
| Apolar M3 | 31.241 | 26.479 | 34.187 | 3.204 | −14.834 | −28.331 |
| Precipitate M3 | 63.198 ** | 34.930 * | 42.609 * | 47.434 | 70.224 * | 68.069 * |
| M4 | −19.942 | −7.420 | 1.159 | 11.487 | −13.040 | −8.933 |
| Andean Shilajit | −9.909 | −9.744 | −8.671 | 31.557 | 2.178 | 28.375 |
| Fulvic Acid | 22.956 | 16.295 | 26.865 | 59.909 * | 10.684 | 68.069 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, V.; Wong-Guerra, M.; Cortés, N.; Pastor, G.; González, A.; Calfío, C.; Guzmán-Martínez, L.; Navarrete, L.P.; Ramos-Escobar, N.; Morales, I.; et al. Scaling the Andean Shilajit: A Novel Neuroprotective Agent for Alzheimer’s Disease. Pharmaceuticals 2023, 16, 960. https://doi.org/10.3390/ph16070960
Andrade V, Wong-Guerra M, Cortés N, Pastor G, González A, Calfío C, Guzmán-Martínez L, Navarrete LP, Ramos-Escobar N, Morales I, et al. Scaling the Andean Shilajit: A Novel Neuroprotective Agent for Alzheimer’s Disease. Pharmaceuticals. 2023; 16(7):960. https://doi.org/10.3390/ph16070960
Chicago/Turabian StyleAndrade, Víctor, Maylin Wong-Guerra, Nicole Cortés, Gabriela Pastor, Andrea González, Camila Calfío, Leonardo Guzmán-Martínez, Leonardo P. Navarrete, Nicolas Ramos-Escobar, Inelia Morales, and et al. 2023. "Scaling the Andean Shilajit: A Novel Neuroprotective Agent for Alzheimer’s Disease" Pharmaceuticals 16, no. 7: 960. https://doi.org/10.3390/ph16070960