
Structural Characterization and Anticancer Activity of a New Anthraquinone from Senna velutina (Fabaceae)

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Results
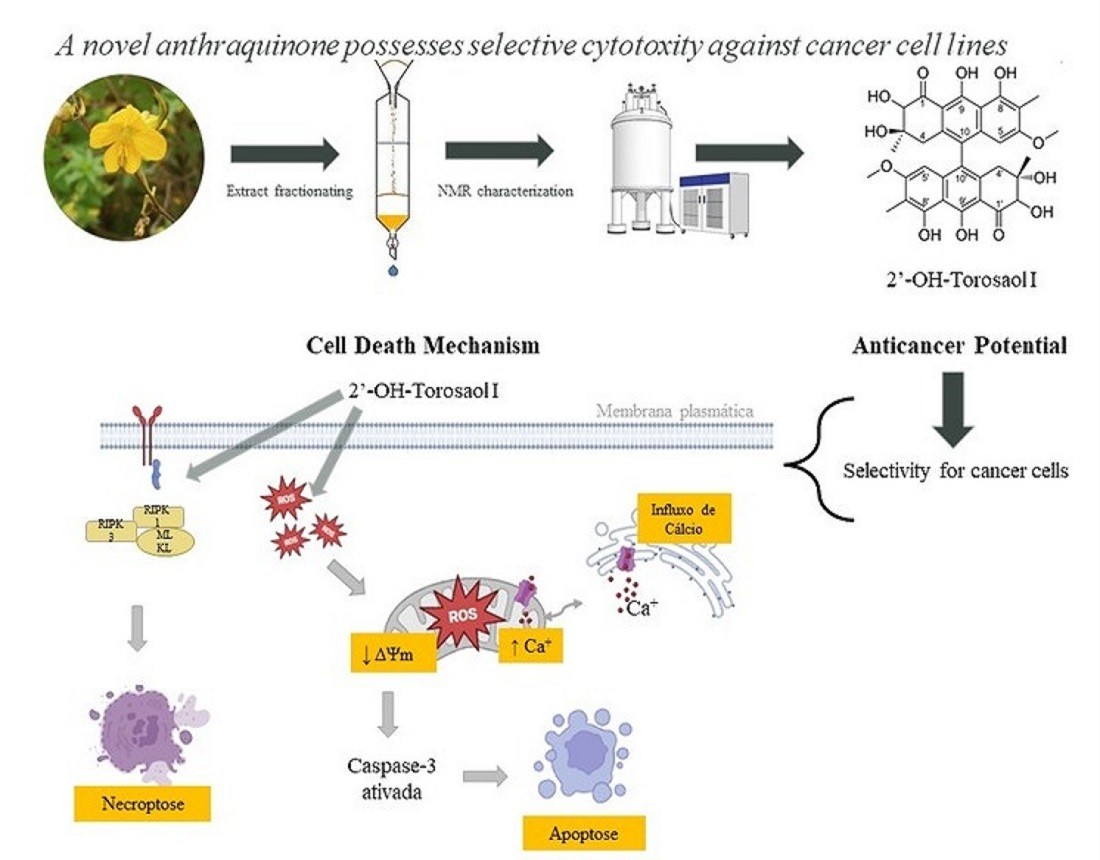
2.1. Structural Determination of 2′-OH-Torosaol I, Which Is Also Referred to as 2,2′-OH- Singueanol I (1)
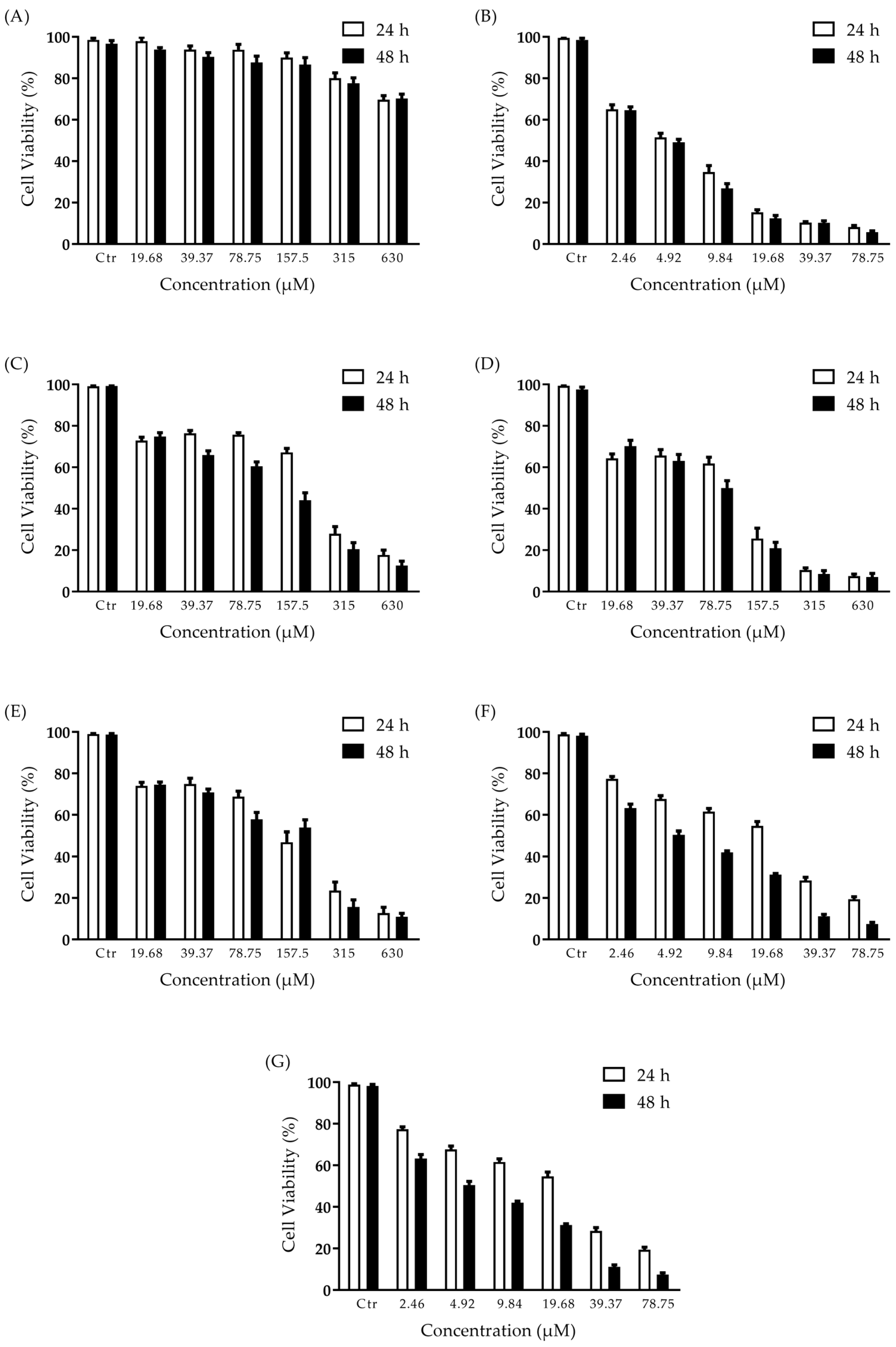
2.2. Effect of 2′-OH-Torosaol (1) on Cell Viability
2.3. Evaluation of 2′-OH-Torosaol (1) in Suppressing Cell Migration of Melanoma Cells
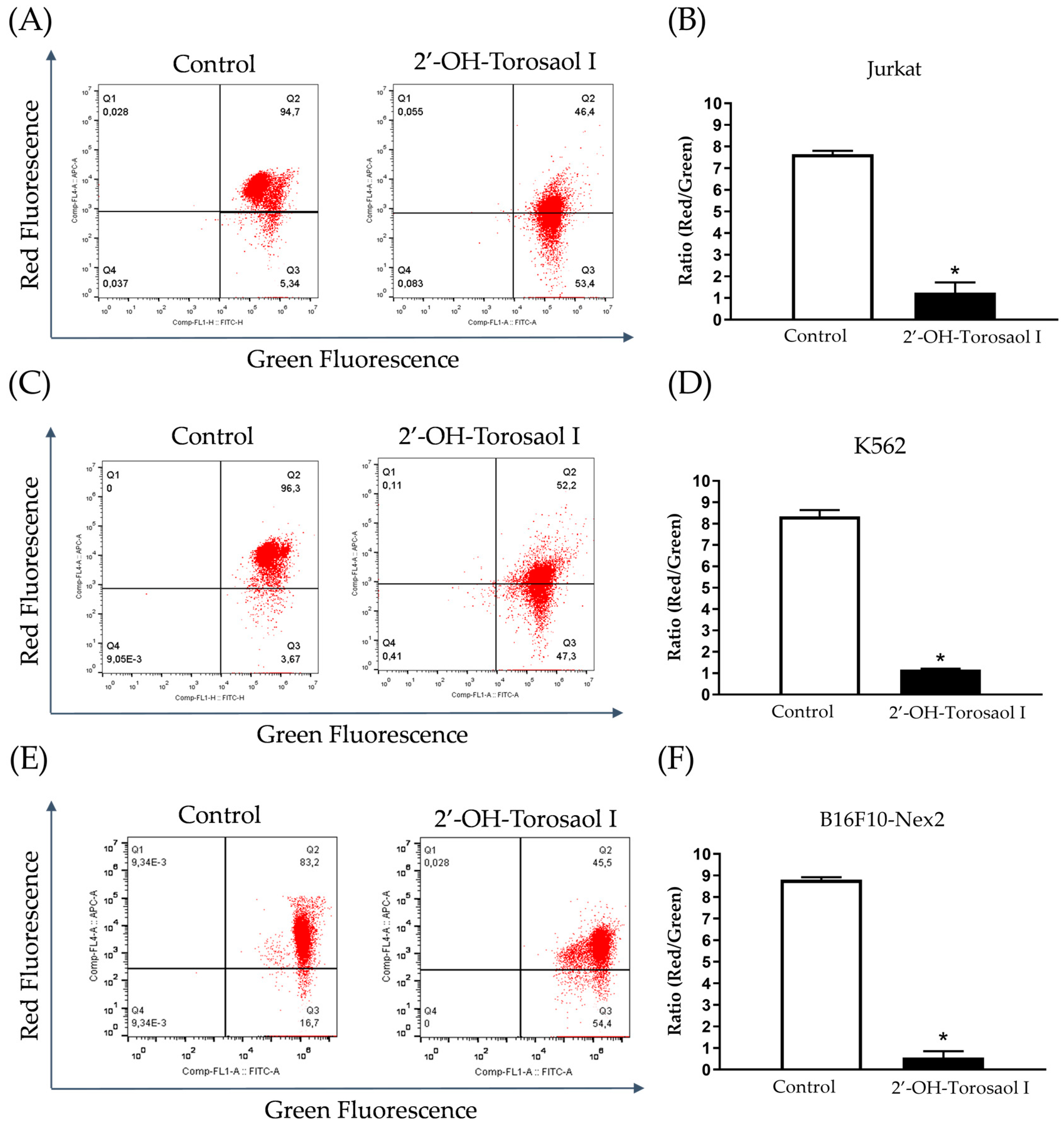
2.4. Analysis of the Cell Death Profile Induced by 2′-OH-Torosaol I
2.5. Assessment of Mitochondrial Membrane Potential (MMP) in Melanoma and Leukemic Cells
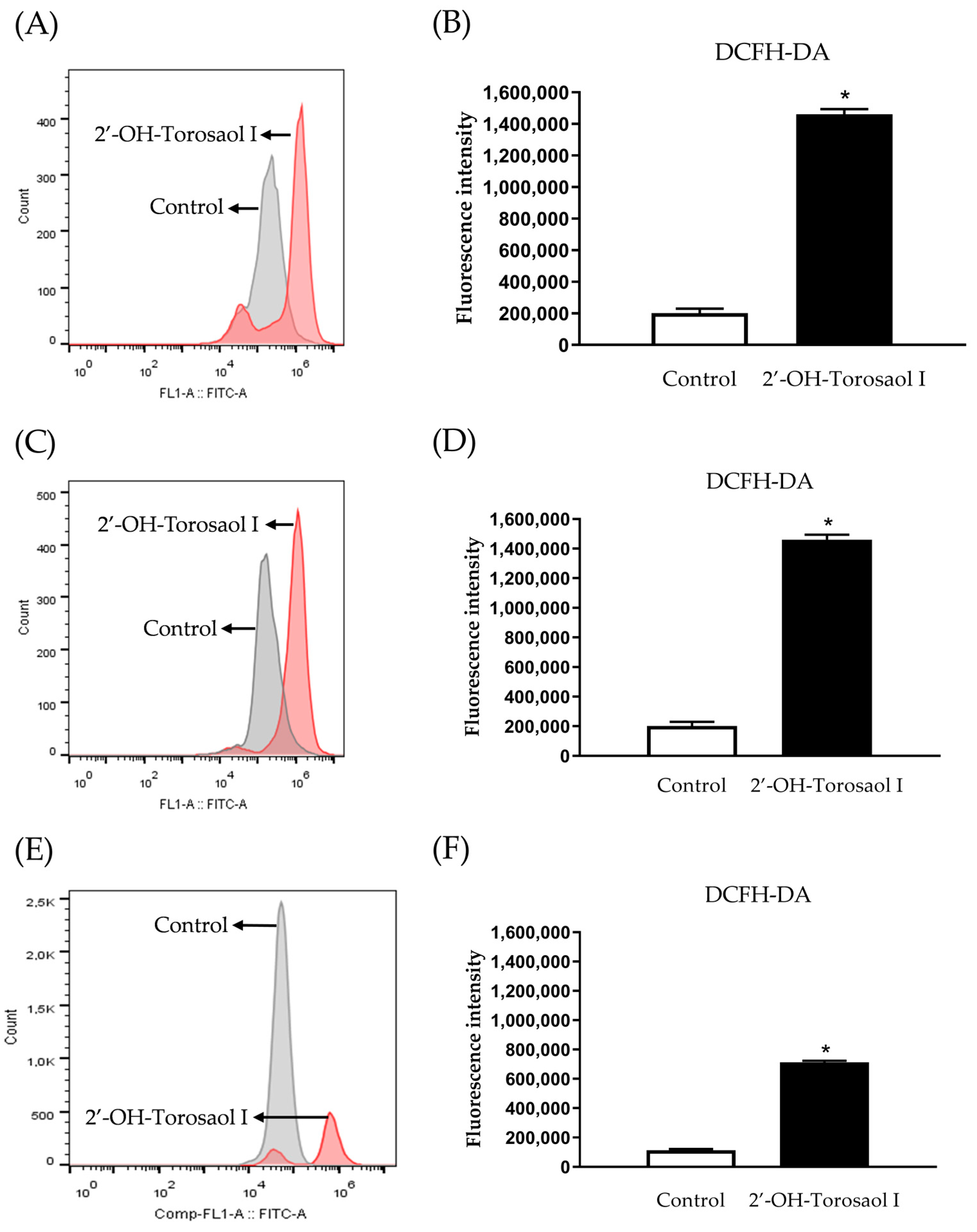
2.6. Determination of Levels of Reactive Oxygen Species (ROS)
2.7. Evaluation of Activated Caspase-3 Protein
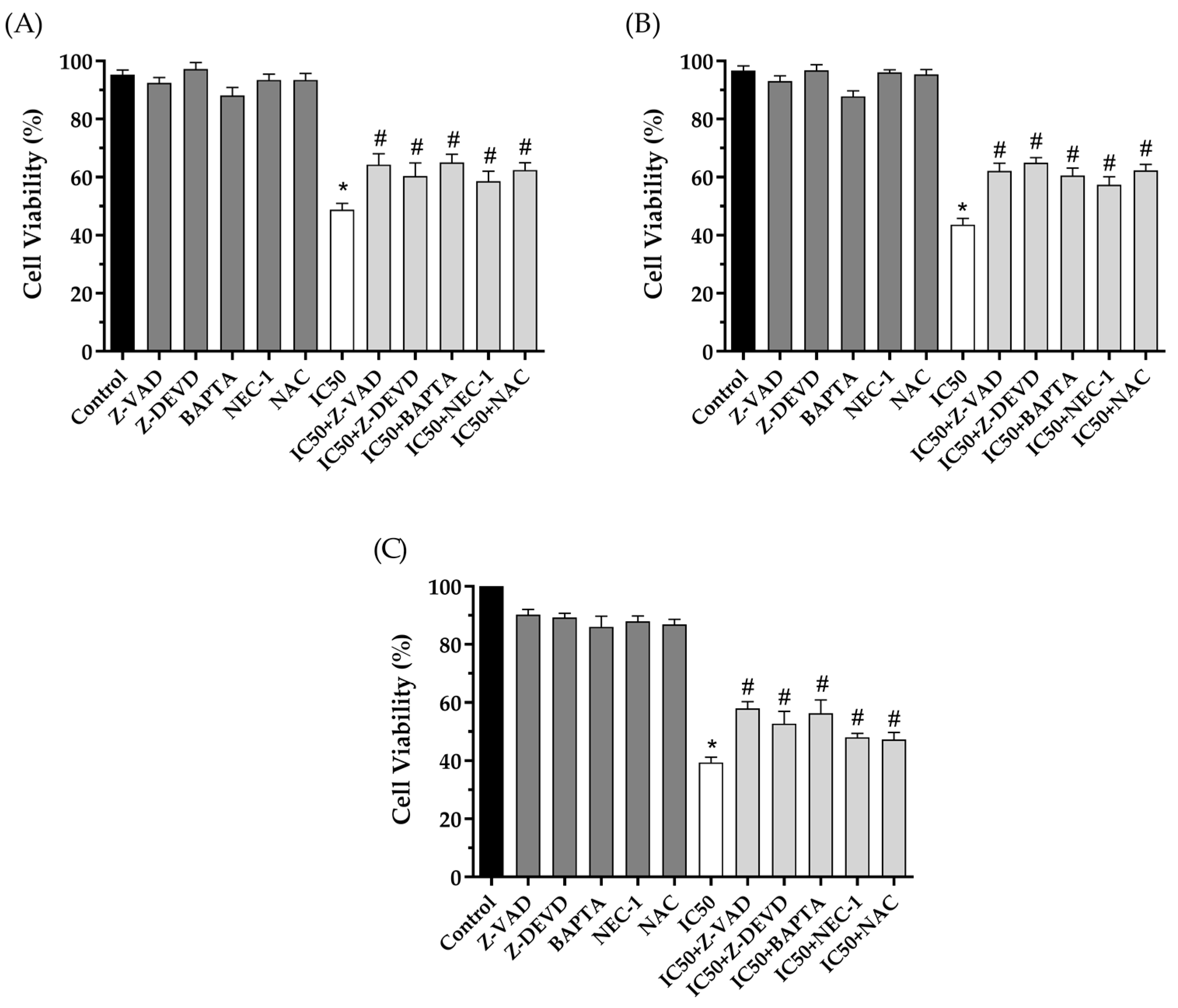
2.8. Cell Death Analysis Using Inhibitors of Cell Death Pathways
3. Discussion
4. Materials and Methods
4.1. Collection and Preparation of Plant Material
4.2. The Roots Extract Fractionating Process
4.3. HPLC-DAD-MS Analysis
4.4. Structural Classification of theCompound by NMR
4.5. Cell Culture
4.6. Cell Viability Assay
4.7. Scratch Assay
4.8. Flow Cytometry Evaluation of Apoptosis
4.9. Measurement of Mitochondrial Membrane Potential (Δψm)
4.10. Measurement of the Generation of Intracellular Reactive Oxygen Species (ROS)
4.11. Measurement of Activated Caspase-3
4.12. Cellular Cytotoxicity Assay with Pharmacological Inhibitors
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhatnagar, N.; Qureshi, A.; Hall, G. Leukaemias: A Review. Paediatr. Child. Health 2017, 27, 489–494. [Google Scholar] [CrossRef]
- Baldivia, D.D.S.; Leite, D.F.; de Castro, D.T.H.; Campos, J.F.; Dos Santos, U.P.; Paredes-Gamero, E.J.; Carollo, C.A.; Silva, D.B.; Souza, K.D.P.; Dos Santos, E.L. Evaluation of In Vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens. Int. J. Mol. Sci. 2018, 19, 2432. [Google Scholar] [CrossRef] [Green Version]
- Chummun, S.; McLean, N.R. The Management of Malignant Skin Cancers. Surgery 2017, 35, 519–524. [Google Scholar] [CrossRef]
- Naik, P.P. Cutaneous Malignant Melanoma: A Review of Early Diagnosis and Management. World J. Oncol. 2021, 12, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Maher, T.; Raus, R.A.; Daddiouaissa, D.; Ahmad, F.; Adzhar, N.S.; Latif, E.S.; Abdulhafiz, F.; Mohammed, A. Medicinal Plants with Anti-Leukemic Effects: A Review. Molecules 2021, 26, 2741. [Google Scholar] [CrossRef]
- Curti, B.D.; Faries, M.B. Recent Advances in the Treatment of Melanoma. N. Engl. J. Med. 2021, 384, 73–74. [Google Scholar] [CrossRef]
- Switzer, B.; Puzanov, I.; Skitzki, J.J.; Hamad, L.; Ernstoff, M.S. Managing Metastatic Melanoma in 2022: A Clinical Review. JCO Oncol. Pract. 2022, 18, 335–351. [Google Scholar] [CrossRef]
- Hu, X.; Yang, Z.; Liu, W.; Pan, Z.; Zhang, X.; Li, M.; Liu, X.; Zheng, Q.; Li, D. The Anti-Tumor Effects of p- Coumaric Acid on Melanoma A375 and B16 Cells. Front. Oncol. 2020, 10, 558414. [Google Scholar] [CrossRef]
- Prakash, O.; Kumar, A.; Kumar, P.; Ajeet, A. Anticancer Potential of Plants and Natural Products: A Review. Am. J. Pharmacol. Sci. 2013, 1, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Jeong, M.I.; Kim, H.R.; Park, H.; Moon, W.K.; Kim, B. Plant Extracts as Possible Agents for Sequela of Cancer Therapies and Cachexia. Antioxidants 2020, 9, 836. [Google Scholar] [CrossRef]
- Kato, N.N.; Stavis, V.K.; Boaretto, A.G.; Castro, D.T.H.; Alves, F.M.; de Picoli Souza, K.; dos Santos, E.L.; Silva, D.B.; Carollo, C.A. Application of the Metabolomics Approach to the Discovery of Active Compounds from Brazilian Trees against Resistant Human Melanoma Cells. Phytochem. Anal. 2021, 32, 992–1002. [Google Scholar] [CrossRef]
- Guimaraes Amaral, R.; dos Santos, S.; Andrade, L.N.; Severino, P.; Carvalho, A.A. Natural Products as Treatment against Cancer: A Historical and Current Vision. Clin. Oncol. 2019, 4, 1562. [Google Scholar]
- Campos, J.F.; De Castro, D.T.H.; Damiaõ, M.J.; Vieira Torquato, H.F.; Paredes-Gamero, E.J.; Carollo, C.A.; Estevinho, L.M.; De Picoli Souza, K.; Santos, E.L. Dos The Chemical Profile of Senna Velutina Leaves and Their Antioxidant and Cytotoxic Effects. Oxid. Med. Cell. Longev. 2016, 2016, 8405957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, D.T.H.; Campos, J.F.; Damião, M.J.; Torquato, H.F.V.; Paredes-Gamero, E.J.; Carollo, C.A.; Rodrigues, E.G.; de Picoli Souza, K.; dos Santos, E.L. Ethanolic Extract of Senna Velutina Roots: Chemical Composition, In Vitro and In Vivo Antitumor Effects, and B16F10-Nex2 Melanoma Cell Death Mechanisms. Oxid. Med. Cell. Longev. 2019, 2019, 5719483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, E.M.; Müller, C.E. Anthraquinones As Pharmacological Tools and Drugs. Med. Res. Rev. 2016, 36, 705–748. [Google Scholar] [CrossRef]
- Chiang, J.H.; Yang, J.S.; Ma, C.Y.; Yang, M.D.; Huang, H.Y.; Hsia, T.C.; Kuo, H.M.; Wu, P.P.; Lee, T.H.; Chung, J.G. Danthron, an Anthraquinone Derivative, Induces DNA Damage and Caspase Cascades-Mediated Apoptosis in SNU-1 Human Gastric Cancer Cells through Mitochondrial Permeability Transition Pores and Bax-Triggered Pathways. Chem. Res. Toxicol. 2011, 24, 20–29. [Google Scholar] [CrossRef]
- Suboj, P.; Babykutty, S.; Srinivas, P.; Gopala, S. Aloe Emodin Induces G2/M Cell Cycle Arrest and Apoptosis via Activation of Caspase-6 in Human Colon Cancer Cells. Pharmacology 2012, 89, 91–98. [Google Scholar] [CrossRef]
- Dong, X.; Ni, B.; Fu, J.; Yin, X.; You, L.; Leng, X.; Liang, X.; Ni, J. Emodin Induces Apoptosis in Human Hepatocellular Carcinoma HepaRG Cells via the Mitochondrial Caspase-dependent Pathway. Oncol. Rep. 2018, 40, 1985–1993. [Google Scholar] [CrossRef]
- Hsu, J.L.; Liu, S.P.; Lee, C.C.; Hsu, L.C.; Ho, Y.F.; Huang, H.S.; Guh, J.H. A Unique Amidoanthraquinone Derivative Displays Antiproliferative Activity against Human Hormone-Refractory Metastatic Prostate Cancers through Activation of LKB1-AMPK-MTOR Signaling Pathway. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 979–990. [Google Scholar] [CrossRef]
- Tian, W.; Wang, C.; Li, D.; Hou, H. Novel Anthraquinone Compounds as Anticancer Agents and Their Potential Mechanism. Future Med. Chem. 2020, 12, 627–644. [Google Scholar] [CrossRef]
- Malik, M.S.; Alsantali, R.I.; Jassas, R.S.; Alsimaree, A.A.; Syed, R.; Alsharif, M.A.; Kalpana, K.; Morad, M.; Althagafi, I.I.; Ahmed, S.A. Journey of Anthraquinones as Anticancer Agents—A Systematic Review of Recent Literature. RSC Adv. 2021, 11, 35806–35827. [Google Scholar] [CrossRef] [PubMed]
- Endo, M.; Naoki, H. Antimicrobial and Antispasmodic Tetrahydroanthracenes from Cassia singueana. Tetrahedron 1980, 36, 2449–2452. [Google Scholar] [CrossRef]
- Beattie, K.; Elsworth, C.; Gill, M.; Milanovic, N.M.; Prima-Putra, D.; Raudies, E. Austrocolorins A1 and B1: Atropisomeric 10,10′-Linked Dihydroanthracenones from an Australian Dermocybe Sp. Phytochemistry 2004, 65, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Oliveira, P.; Otero, P.; Pereira, A.G.; Chamorro, F.; Carpena, M.; Echave, J.; Fraga-Corral, M.; Simal-Gandara, J.; Prieto, M.A. Status and Challenges of Plant-Anticancer Compounds in Cancer Treatment. Pharmaceuticals 2021, 14, 157. [Google Scholar] [CrossRef] [PubMed]
- Bolzani, V.D.S.; Valli, M.; Pivatto, M.; Viegas, C. Natural Products from Brazilian Biodiversity as a Source of New Models for Medicinal Chemistry. Pure Appl. Chem. 2012, 84, 1837–1846. [Google Scholar] [CrossRef]
- Fouillaud, M.; Venkatachalam, M.; Girard-Valenciennes, E.; Caro, Y.; Dufossé, L. Anthraquinones and Derivatives from Marine-Derived Fungi: Structural Diversity and Selected Biological Activities. Mar. Drugs 2016, 14, 64. [Google Scholar] [CrossRef] [Green Version]
- Kadam, S.; Kanase, V. Laxative Activity of Ethanolic Extract of Capparis moonii W. Fruit. Res. J. Pharm. Technol. 2021, 14, 3528–3532. [Google Scholar] [CrossRef]
- Wuthi-udomlert, M.; Kupittayanant, P.; Gritsanapan, W. In vitro evaluation of antifungal activity of anthraquinone derivatives of Senna alata. J. Health Res. 2010, 24, 117–122. [Google Scholar]
- Fosso, M.Y.; Chan, K.Y.; Gregory, R.; Chang, C.W.T. Library Synthesis and Antibacterial Investigation of Cationic Anthraquinone Analogs. ACS Comb. Sci. 2012, 14, 231–235. [Google Scholar] [CrossRef]
- Barnard, D.L.; Fairbairn, D.W.; O’Neill, K.L.; Gage, T.L.; Sidwell, R.W. Anti-Human Cytomegalovirus Activity and Toxicity of Sulfonated Anthraquinones and Anthraquinone Derivatives. Antiviral Res. 1995, 28, 317–329. [Google Scholar] [CrossRef]
- Kshirsagar, A.D.; Panchal, P.V.; Harle, U.N.; Nanda, R.K.; Shaikh, H.M. Anti-Inflammatory and Antiarthritic Activity of Anthraquinone Derivatives in Rodents. Int. J. Inflamm. 2014, 2014, 690596. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Lu, G.; Shen, H.M.; Chung, M.C.M.; Choon, N.O. Anti-Cancer Properties of Anthraquinones from Rhubarb. Med. Res. Rev. 2007, 27, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, J.P.; Subedi, Y.P.; Chen, L.; Chang, C.W.T. A Mode of Action Study of Cationic Anthraquinone Analogs: A New Class of Highly Potent Anticancer Agents. Medchemcomm 2015, 6, 2012–2022. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef] [Green Version]
- Branco, A.; Pinto, A.C.; Schripsema, J.; Braz-Filho, R. Anthraquinones from the Bark of Senna macranthera. An. Acad. Bras. Cienc. 2011, 83, 1159–1163. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.H.; Pandey, R.P.; Lee, C.M.; Sim, J.S.; Jeong, J.T.; Choi, B.S.; Jung, M.; Ginzburg, D.; Zhao, K.; Won, S.Y.; et al. Genome-Enabled Discovery of Anthraquinone Biosynthesis in Senna tora. Nat. Commun. 2020, 11, 5875. [Google Scholar] [CrossRef]
- Kitanaka, S.; Takido, M. Two New Bitetrahydroanthracenes from Roots of Cassia occidentalis L. Chem. Pharm. Bull. 1989, 37, 511–512. [Google Scholar] [CrossRef] [Green Version]
- Nixon, A.B.; Schalper, K.A.; Jacobs, I.; Potluri, S.; Wang, I.M.; Fleener, C. Peripheral Immune-Based Biomarkers in Cancer Immunotherapy: Can We Realize Their Predictive Potential? J. Immunother. Cancer 2019, 7, 325. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.; Sun, C.; Li, F.; Wu, Y.; Zhang, G.; Gu, Q.; Zhu, T.; Li, D.; Che, Q. Dimeric Tetrahydroanthracene Regioisomers and Their Monomeric Precursor Produced by Streptomyces fumigatiscleroticus HDN10255. J. Nat. Prod. 2020, 83, 2797–2802. [Google Scholar] [CrossRef]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular Principles of Metastasis: A Hallmark of Cancer Revisited. Signal. Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Decker, C.C.; Zechner, L.; Krstin, S.; Wink, M. In Vitro Wound Healing of Tumor Cells: Inhibition of Cell Migration by Selected Cytotoxic Alkaloids. BMC Pharmacol. Toxicol. 2019, 20, 4. [Google Scholar] [CrossRef] [Green Version]
- Roussos, E.T.; Condeelis, J.S.; Patsialou, A. Chemotaxis in Cancer. Nat. Rev. Cancer 2011, 11, 573–587. [Google Scholar] [CrossRef]
- Brauchle, E.; Thude, S.; Brucker, S.Y.; Schenke-Layland, K. Cell Death Stages in Single Apoptotic and Necrotic Cells Monitored by Raman Microspectroscopy. Sci. Rep. 2014, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.T. Secondary Necrosis: The Natural Outcome of the Complete Apoptotic Program. FEBS Lett. 2010, 584, 4491–4499. [Google Scholar] [CrossRef] [Green Version]
- Poon, I.K.H.; Hulett, M.D.; Parish, C.R. Molecular Mechanisms of Late Apoptotic/Necrotic Cell Clearance. Cell. Death Differ. 2010, 17, 381–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, V.A.; Longacre, A.; Hsiao, K.; Fan, H.; Meng, F.; Mitchell, J.E.; Rauch, J.; Ucker, D.S.; Levine, J.S. Apoptotic Cells, at All Stages of the Death Process, Trigger Characteristic Signaling Events That Are Divergent from and Dominant over Those Triggered by Necrotic Cells: Implications for the Delayed Clearance Model of Autoimmunity. J. Biol. Chem. 2006, 281, 4663–4670. [Google Scholar] [CrossRef] [Green Version]
- Birge, R.B.; Boeltz, S.; Kumar, S.; Carlson, J.; Wanderley, J.; Calianese, D.; Barcinski, M.; Brekken, R.A.; Huang, X.; Hutchins, J.T.; et al. Phosphatidylserine Is a Global Immunosuppressive Signal in Efferocytosis, Infectious Disease, and Cancer. Cell. Death Differ. 2016, 23, 962–978. [Google Scholar] [CrossRef] [Green Version]
- Shlomovitz, I.; Speir, M.; Gerlic, M. Flipping the Dogma—Phosphatidylserine in Non-Apoptotic Cell Death. Cell. Commun. Signal. 2019, 17, 139. [Google Scholar] [CrossRef] [Green Version]
- Naderi, M.; Cherati, M.R.; Mohammadian, A.; Bidhendy, M.B.; Ghiasvand, S.; Marzouni, H.Z.; Aryan, H.; Jan-gholi, E.; Javidi, M.A. Hypericin Induces Apoptosis in AGS Cell Line with No Significant Effect on Normal Cells. Iran J Pharm Res 2020, 19, 349. [Google Scholar] [CrossRef]
- Le, J.; Ji, H.; Pi, P.; Chen, K.; Gu, X.; Ma, Y.; Fu, Y.; Sun, Y.; Zhou, X.; Wu, H. The Effects and Mechanisms of Sennoside A on Inducing Cytotoxicity, Apoptosis, and Inhibiting Metastasis in Human Chondrosarcoma Cells. Evid. Based Complement. Alternat Med. 2022, 2022, 8063497. [Google Scholar] [CrossRef]
- Jiang, H.; Tang, W.; Song, Y.; Jin, W.; Du, Q. Induction of Apoptosis by Metabolites of Rhei Radix et Rhizoma (Da Huang): A Review of the Potential Mechanism in Hepatocellular carcinoma. Front. Pharmacol. 2022, 13, 806175. [Google Scholar] [CrossRef]
- Delierneux, C.; Kouba, S.; Shanmughapriya, S.; Potier-Cartereau, M.; Trebak, M.; Hempel, N. Mitochondrial Calcium Regulation of Redox Signaling in Cancer. Cells 2020, 9, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K.; Uchida, T.; Yoshie, T.; Mizote, Y.; Ishikawa, F.; Katsuyama, M.; Shibanuma, M. A Mitochondrial ROS Pathway Controls Matrix Metalloproteinase 9 Levels and Invasive Properties in RAS-Activated Cancer Cells. FEBS J. 2019, 286, 459–478. [Google Scholar] [CrossRef] [Green Version]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of Cytochrome c Release from Mitochondria. Cell. Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Llambi, F.; Green, D.R. Apoptosis and Oncogenesis: Give and Take in the BCL-2 Family. Curr. Opin. Genet. Dev. 2011, 21, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zheng, J.; Nussinov, R.; Ma, B. Release of Cytochrome C from Bax Pores at the Mitochondrial Membrane. Sci. Rep. 2017, 7, 2635. [Google Scholar] [CrossRef] [Green Version]
- Boice, A.; Bouchier-Hayes, L. Targeting Apoptotic Caspases in Cancer. Biochim. Biophys. Acta Mol. Cell. Res. 2020, 1867, 118688. [Google Scholar] [CrossRef] [PubMed]
- Julien, O.; Wells, J.A. Caspases and Their Substrates. Cell. Death Differ. 2017, 24, 1380–1389. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of Apoptosis Signalling Pathways by Reactive Oxygen Species. Biochim. Biophys. Acta (BBA)—Mol. Cell. Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of Cell Death: The Calcium-Apoptosis Link. Nat. Rev. Mol. Cell. Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Sukumaran, P.; Da Conceicao, V.N.; Sun, Y.; Ahamad, N.; Saraiva, L.R.; Selvaraj, S.; Singh, B.B. Calcium Signaling Regulates Autophagy and Apoptosis. Cells 2021, 10, 2125. [Google Scholar] [CrossRef]
- Choi, M.E.; Price, D.R.; Ryter, S.W.; Choi, A.M.K. Necroptosis: A Crucial Pathogenic Mediator of Human Disease. JCI Insight 2019, 4, e128834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reczek, C.R.; Chandel, N.S. The Two Faces of Reactive Oxygen Species in Cancer. Annu. Rev. Cancer Biol. 2017, 1, 79–98. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in Cancer Therapy: The Bright Side of the Moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive Oxygen Species and Cancer Paradox: To Promote or to Suppress? Free. Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In Vitro Cell Migration and Invasion Assays. J. Vis. Exp. 2014, 88, e51046. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H/C | 13C (δ) | 1H (mult., J in Hz, int.) | HMBC |
|---|---|---|---|
| 1,1′ | 205.2 | - | - |
| 2,2′ | 78.1 | 4.35 (s, 2H) | C-1/1′, C-3/3′, C-3/3′-Me |
| 3,3′ | 73.1 | - | |
| 4,4′ | 39.8 | 2.32 (d, 16.8, 2H) 2.71(d, 16.8, 2H) | C-4/4′, C-4a/4a’, C-10/10′ |
| 4a,4a’ | 135.2 | - | - |
| 5,5′ | 95.8 | 5.96 (s, 2H) | C-7/7′-Me, C-10/10′, C-7/7′, C-10/10′ |
| 6,6′ | 161.2 | - | - |
| 7,7′ | 109.7 | - | - |
| 8,8′ | 155.1 | - | - |
| 8a,8a’ | 137.2 | - | - |
| 9,9′ | 162.6 | - | - |
| 9a,9a’ | 107.1 | - | - |
| 10,10′ | 122.9 | - | - |
| 10a,10a’ | 108.2 | - | - |
| 3,3′-Me | 25.9 | 1.09 (s, 6H) | C-2/2′, C-3/3′, C-4/4′, C-4a/4a’ |
| 7,7′-Me | 8.1 | 2.01 (s, 6H) | C-6/6′, C-8/8′, C-7/7′, C-8a/8a’ |
| 6,6′-OMe | 55.1 | 3.38 (s, 6H) | C6/6′ |
| 3,3′-OH | - | 1.17 (s br, 2H) | - |
| 8,8′-OH | - | 10.05 (s br, 2H) | - |
| 9,9′-OH | - | 15.95 (s br, 2H) | - |
| IC50 (µM) | ||
|---|---|---|
| 2′-OH-Torosaol I | ||
| Cell Lines | 24 h | 48 h |
| PBMC | > 630 | > 630 |
| B16F10-Nex2 | 5.8 ± 0.7 | 4.7 ± 0.9 |
| SK-MEL-19 | 217.8 ± 12.8 | 139.21 ± 19.9 |
| SK-MEL-28 | 108.4 ± 13.2 | 73.55 ± 11.0 |
| SK-MEL-103 | 164.6 ± 14.6 | 172.5 ± 27.2 |
| JURKAT | 22.8 ± 4.5 | 4.89 ± 0.1 |
| K562 | 21.7 ± 4.2 | 5.21 ± 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, D.T.H.; Leite, D.F.; da Silva Baldivia, D.; dos Santos, H.F.; Balogun, S.O.; da Silva, D.B.; Carollo, C.A.; de Picoli Souza, K.; dos Santos, E.L. Structural Characterization and Anticancer Activity of a New Anthraquinone from Senna velutina (Fabaceae). Pharmaceuticals 2023, 16, 951. https://doi.org/10.3390/ph16070951
Castro DTH, Leite DF, da Silva Baldivia D, dos Santos HF, Balogun SO, da Silva DB, Carollo CA, de Picoli Souza K, dos Santos EL. Structural Characterization and Anticancer Activity of a New Anthraquinone from Senna velutina (Fabaceae). Pharmaceuticals. 2023; 16(7):951. https://doi.org/10.3390/ph16070951
Chicago/Turabian StyleCastro, David Tsuyoshi Hiramatsu, Daniel Ferreira Leite, Debora da Silva Baldivia, Helder Freitas dos Santos, Sikiru Olaitan Balogun, Denise Brentan da Silva, Carlos Alexandre Carollo, Kely de Picoli Souza, and Edson Lucas dos Santos. 2023. "Structural Characterization and Anticancer Activity of a New Anthraquinone from Senna velutina (Fabaceae)" Pharmaceuticals 16, no. 7: 951. https://doi.org/10.3390/ph16070951