
High Selectivity of 8-Hydroxyquinoline on Leishmania (Leishmania) and Leishmania (Viannia) Species Correlates with a Potent Therapeutic Activity In Vivo

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. In Vitro Studies
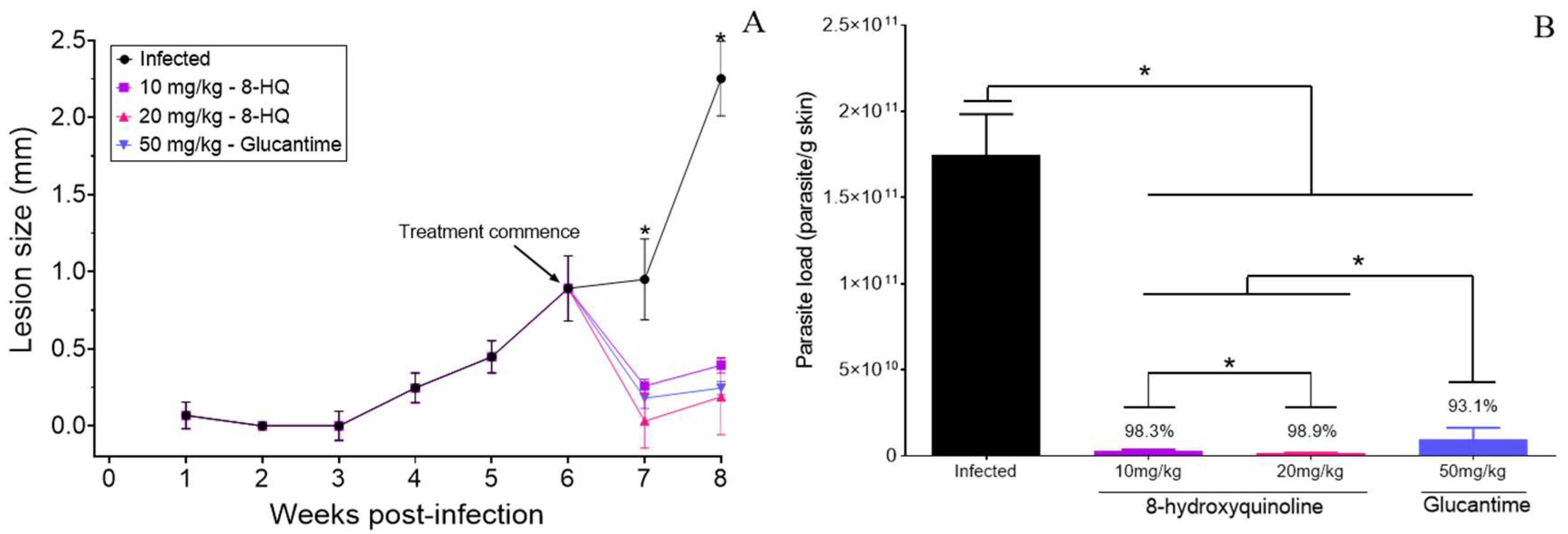
2.2. Efficacy of 8-HQ Administered by the Intralesional Route
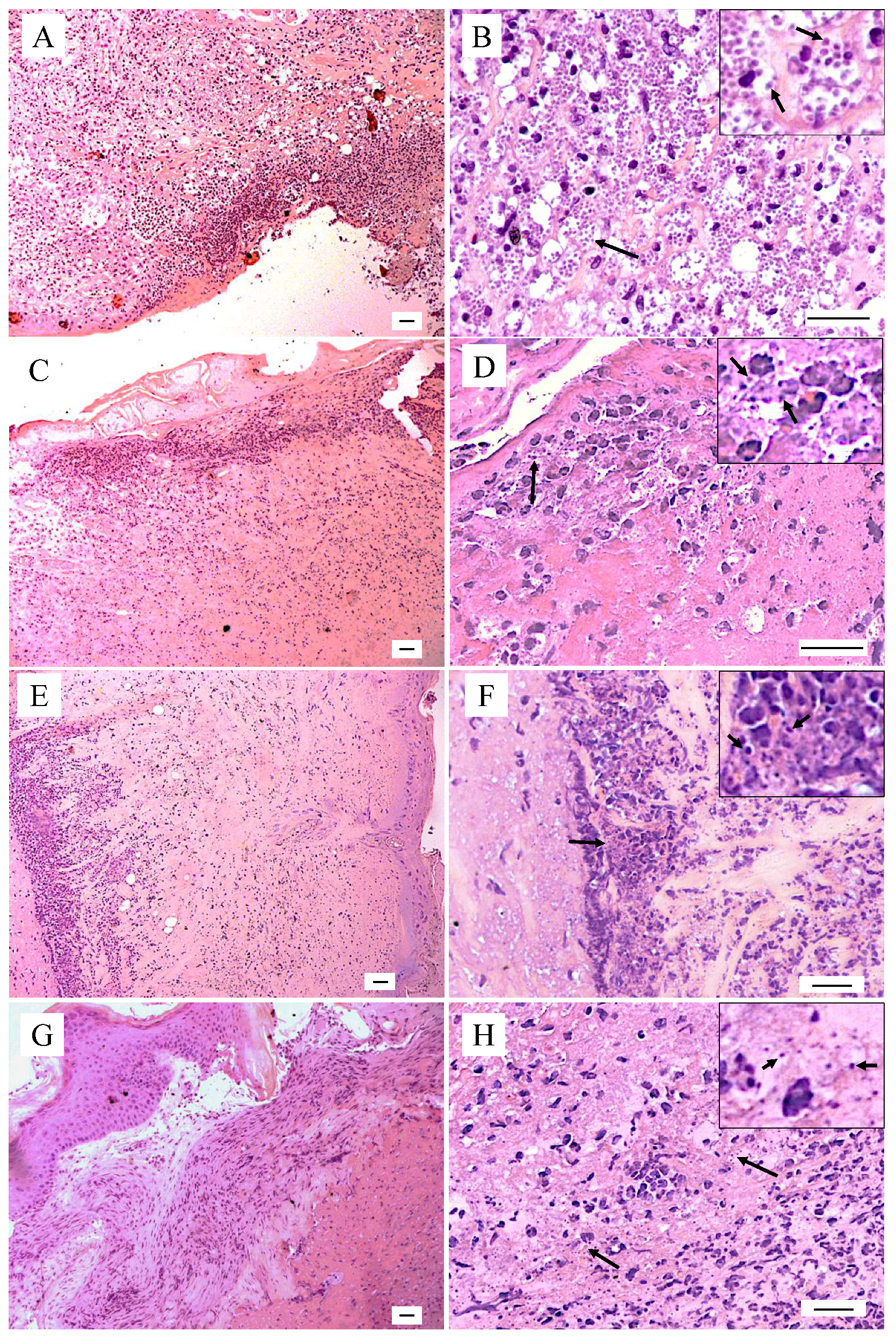
2.3. Histopathological Changes
2.4. Immunological Studies
3. Discussion
4. Materials and Methods
4.1. Culture Media and Drugs
4.2. Animals
4.3. Parasite Strains
4.4. Promastigote Assay
4.5. Bone Marrow-Derived Macrophages (BMMs)
4.5.1. Cytotoxicity Assay
4.5.2. Intracellular Infection and Treatment (In Vitro)
4.6. Determination of Intracellular Nitric Oxide Production
4.7. Infection and Treatment
Clinical Course of Lesion Development and Determination of Parasite Burden in the Skin of Infected and Treated Animals
4.8. Quantification of Cytokines
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gurel, M.S.; Tekin, B.; Uzun, S. Cutaneous Leishmaniasis: A Great Imitator. Clin. Dermatol. 2020, 38, 140–151. [Google Scholar] [CrossRef]
- Saha, B.; Bhattacharjee, S.; Sarkar, A.; Bhor, R.; Pai, K.; Bodhale, N. Conundrums in Leishmaniasis. Cytokine 2021, 145, 155304. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Bhattacharjee, S.; Sarkar, A.; Bhor, R.; Pai, K.; Bodhale, N.; Serafim, T.D.; Coutinho-Abreu, I.V.; Dey, R.; Kissinger, R.; et al. Leishmaniasis: The Act of Transmission. Cytokine 2021, 37, 976–987. [Google Scholar] [CrossRef]
- Kaushal, R.S.; Naik, N.; Prajapati, M.; Rane, S.; Raulji, H.; Ngo, F.A.; Upadhyay, T.K.; Saeed, M. Leishmania Species: A Narrative Review on Surface Proteins with Structural Aspects Involved in Host-pathogen Interaction. Chem. Biol. Drug Des. 2023, 1, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Reithinger, R.; Dujardin, J.C.; Louzir, H.; Pirmez, C.; Alexander, B.; Brooker, S. Cutaneous Leishmaniasis. Lancet Infect. Dis. 2007, 7, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Real, F.; Florentino, P.T.V.; Reis, L.C.; Ramos-Sanchez, E.M.; Veras, P.S.T.; Goto, H.; Mortara, R.A. Cell-to-Cell Transfer of Leishmania Amazonensis Amastigotes Is Mediated by Immunomodulatory LAMP-Rich Parasitophorous Extrusions. Cell. Microbiol. 2014, 16, 1549–1564. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Ghatee, M.A.; Taylor, W.R.; Karamian, M. The Geographical Distribution of Cutaneous Leishmaniasis Causative Agents in Iran and Its Neighboring Countries, A Review. Front. Public Health 2020, 8, 11. [Google Scholar] [CrossRef]
- Silveira, F.T.; Lainson, R.; De Castro Gomes, C.M.; Laurenti, M.D.; Corbett, C.E.P. Immunopathogenic Competences of Leishmania (V.) braziliensis and L. (L.) amazonensis in American Cutaneous Leishmaniasis. Parasite Immunol. 2009, 31, 423–431. [Google Scholar] [CrossRef]
- Laurenti, M.D.; Sosa-Ochoa, W.; Araujo Flores, G.V.; Sandoval Pacheco, C.M.; Tomokane, T.Y.; Oliveira, L.M.d.S.; Zúniga, C.; Silveira, F.T.; Corbett, C.E.P. Evaluation of Systemic Immunity in Atypical Cutaneous Leishmaniasis Caused by Leishmania (L.) Infantum Chagasi. Parasite Immunol. 2022, 44, 1–2. [Google Scholar] [CrossRef]
- Campos, M.B.; De Castro Gomes, C.M.; De Souza, A.A.A.; Lainson, R.; Corbett, C.E.P.; Silveira, F.T. In Vitro Infectivity of Species of Leishmania (Viannia) Responsible for American Cutaneous Leishmaniasis. Parasitol. Res. 2008, 103, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Silveira, F.T. What Makes Mucosal and Anergic Diffuse Cutaneous Leishmaniases so Clinically and Immunopathogically Different? A Review in Brazil. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in Humans: Drug or Vaccine Therapy? Drug Des. Devel. Ther. 2018, 12, 25–40. [Google Scholar] [CrossRef]
- Taslimi, Y.; Zahedifard, F.; Rafati, S. Leishmaniasis and Various Immunotherapeutic Approaches. Parasitology 2018, 145, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Frézard, F.; Demicheli, C.; Ribeiro, R.R. Pentavalent Antimonials: New Perspectives for Old Drugs. Molecules 2009, 14, 2317–2336. [Google Scholar] [CrossRef]
- Baiocco, P.; Colotti, G.; Franceschini, S.; Ilari, A. Molecular Basis of Antimony Treatment in Leishmaniasis. J. Med. Chem. 2009, 52, 2603–2612. [Google Scholar] [CrossRef]
- Ilari, A.; Genovese, I.; Fiorillo, F.; Battista, T.; De Ionna, I.; Fiorillo, A.; Colotti, G. Toward a Drug Against All Kinetoplastids: From LeishBox to Specific and Potent Trypanothione Reductase Inhibitors. Mol. Pharm. 2018, 15, 3069–3078. [Google Scholar] [CrossRef]
- Battista, T.; Colotti, G.; Ilari, A.; Fiorillo, A. Targeting Trypanothione Reductase, a Key Enzyme in the Redox Trypanosomatid Metabolism, to Develop New Drugs against Leishmaniasis and Trypanosomiases. Molecules 2020, 25, 8. [Google Scholar] [CrossRef]
- Basu, J.M.; Mookerjee, A.; Sen, P.; Bhaumik, S.; Sen, P.; Banerjee, S.; Naskar, K.; Choudhuri, S.K.; Saha, B.; Raha, S.; et al. Sodium Antimony Gluconate Induces Generation of Reactive Oxygen Species and Nitric Oxide via Phosphoinositide 3-Kinase and Mitogen-Activated Protein Kinase Activation in Leishmania Donovani-Infected Macrophages. Antimicrob. Agents Chemother. 2006, 50, 1788–1797. [Google Scholar] [CrossRef]
- Garza-Tovar, T.F.; Sacriste-Hernández, M.I.; Juárez-Durán, E.R.; Arenas, R. An Overview of the Treatment of Cutaneous Leishmaniasis. Fac. Rev. 2020, 9, 28. [Google Scholar] [CrossRef]
- Sundar, S.; Chakravarty, J. Antimony Toxicity. Int. J. Environ. Res. Public Health 2010, 7, 4267–4277. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.F.; Schubach, A.O.; Martins, M.M.; Passos, S.L.; Oliveira, R.V.; Marzochi, M.C.; Andrade, C.A. Systematic Review of the Adverse Effects of Cutaneous Leishmaniasis Treatment in the New World. Acta Trop. 2011, 118, 87–96. [Google Scholar] [CrossRef]
- Chakravarty, J.; Sundar, S. Current and Emerging Medications for the Treatment of Leishmaniasis. Expert Opin. Pharmacother. 2019, 20, 1251–1265. [Google Scholar] [CrossRef] [PubMed]
- Van Griensven, J.; Diro, E. Visceral Leishmaniasis. Infect. Dis. Clin. North Am. 2012, 26, 309–322. [Google Scholar] [CrossRef] [PubMed]
- De Melo, E.C.; Fortaleza, C.M.C.B. Challenges in the Therapy of Visceral Leishmaniasis in Brazil: A Public Health Perspective. J. Trop. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Ponte-Sucre, A.; Gamarro, F.; Dujardin, J.C.; Barrett, M.P.; López-Vélez, R.; García-Hernández, R.; Pountain, A.W.; Mwenechanya, R.; Papadopoulou, B. Drug Resistance and Treatment Failure in Leishmaniasis: A 21st Century Challenge. PLoS Negl. Trop. Dis. 2017, 11, 12. [Google Scholar] [CrossRef]
- Hamill, R.J. Amphotericin B Formulations: A Comparative Review of Efficacy and Toxicity. Drugs 2013, 73, 919–934. [Google Scholar] [CrossRef]
- Mosimann, V.; Neumayr, A.; Paris, D.H.; Blum, J. Liposomal Amphotericin B Treatment of Old World Cutaneous and Mucosal Leishmaniasis: A Literature Review. Acta Trop. 2018, 182, 246–250. [Google Scholar] [CrossRef]
- van Griensven, J.; Diro, E. Visceral Leishmaniasis: Recent Advances in Diagnostics and Treatment Regimens. Infect. Dis. Clin. N. Am. 2019, 33, 79–99. [Google Scholar] [CrossRef]
- Dorlo, T.P.C.; Balasegaram, M.; Beijnen, J.H.; de vries, P.J. Miltefosine: A Review of Its Pharmacology and Therapeutic Efficacy in the Treatment of Leishmaniasis. J. Antimicrob. Chemother. 2012, 67, 2576–2597. [Google Scholar] [CrossRef]
- Palić, S.; Beijnen, J.H.; Dorlo, T.P.C. An Update on the Clinical Pharmacology of Miltefosine in the Treatment of Leishmaniasis. Int. J. Antimicrob. Agents 2022, 59, 106459. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Singh, B. Emerging Therapeutic Targets for Treatment of Leishmaniasis. Expert Opin. Ther. Targets 2018, 22, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Mishra, J.; Gupta, A.K.; Singh, A.; Shankar, P.; Singh, S. Laboratory Confirmed Miltefosine Resistant Cases of Visceral Leishmaniasis from India. Parasites Vectors 2017, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Goto, H. Review of the Current Treatments for Leishmaniases. Res. Rep. Trop. Med. 2012, 3, 69. [Google Scholar] [CrossRef]
- Pradhan, S.; Schwartz, R.A.; Patil, A.; Grabbe, S.; Goldust, M. Treatment Options for Leishmaniasis. Clin. Exp. Dermatol. 2022, 47, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Passero, L.F.D.; Brunelli, E.d.S.; Sauini, T.; Amorim Pavani, T.F.; Jesus, J.A.; Rodrigues, E. The Potential of Traditional Knowledge to Develop Effective Medicines for the Treatment of Leishmaniasis. Front. Pharmacol. 2021, 12, 690432. [Google Scholar] [CrossRef]
- Gupta, O.; Pradhan, T.; Bhatia, R.; Monga, V. Recent Advancements in Anti-Leishmanial Research: Synthetic Strategies and Structural Activity Relationships. Eur. J. Med. Chem. 2021, 223, 113606. [Google Scholar] [CrossRef]
- Pippi, B.; Joaquim, A.R.; Lopes, W.; Machado, G.R.M.; Bergamo, V.Z.; Giuliani, L.M.; Abegg, M.A.; Cruz, L.; Vainstein, M.H.; Fuentefria, A.M.; et al. 8-Hydroxyquinoline-5-Sulfonamides Are Promising Antifungal Candidates for the Topical Treatment of Dermatomycosis. J. Appl. Microbiol. 2020, 128, 1038–1049. [Google Scholar] [CrossRef]
- Gupta, R.; Luxami, V.; Paul, K. Insights of 8-Hydroxyquinolines: A Novel Target in Medicinal Chemistry. Bioorg. Chem. 2021, 108, 104633. [Google Scholar] [CrossRef]
- Silva, E.J.; Bezerra-Souza, A.; Passero, L.F.; Laurenti, M.D.; Ferreira, G.M.; Fujii, D.G.V.; Trossini, G.H.G.; Raminelli, C. Synthesis, Leishmanicidal Activity, Structural Descriptors and Structure-Activity Relationship of Quinoline Derivatives. Future Med. Chem. 2018, 10, 2069–2085. [Google Scholar] [CrossRef]
- Costa Duarte, M.; dos Reis Lage, L.M.; Lage, D.P.; Mesquita, J.T.; Salles, B.C.S.; Lavorato, S.N.; Menezes-Souza, D.; Roatt, B.M.; Alves, R.J.; Tavares, C.A.P.; et al. An Effective in Vitro and in Vivo Antileishmanial Activity and Mechanism of Action of 8-Hydroxyquinoline against Leishmania Species Causing Visceral and Tegumentary Leishmaniasis. Vet. Parasitol. 2016, 217, 81–88. [Google Scholar] [CrossRef]
- Tavares, G.S.V.; Mendonça, D.V.C.; Miyazaki, C.K.; Lage, D.P.; Soyer, T.G.; Carvalho, L.M.; Ottoni, F.M.; Dias, D.S.; Ribeiro, P.A.F.; Antinarelli, L.M.R.; et al. A Pluronic® F127-Based Polymeric Micelle System Containing an Antileishmanial Molecule Is Immunotherapeutic and Effective in the Treatment against Leishmania Amazonensis Infection. Parasitol. Int. 2019, 68, 63–72. [Google Scholar] [CrossRef]
- Tavares, G.d.S.V.; Mendonça, D.V.C.; Lage, D.P.; Granato, J.d.T.; Ottoni, F.M..; Ludolf, F.; Chávez-Fumagalli, M.A.; Duarte, M.C.; Tavares, C.A.P.; Alves, R.J.; et al. Antileishmanial Activity, Cytotoxicity and Mechanism of Action of Clioquinol Against Leishmania Infantum and Leishmania Amazonensis Species. Basic Clin. Pharmacol. Toxicol. 2018, 123, 236–246. [Google Scholar] [CrossRef]
- Allam, G.; Eweas, A.F.; Abuelsaad, A.S.A. In Vivo Schistosomicidal Activity of Three Novels 8-Hydroxyquinoline Derivatives against Adult and Immature Worms of Schistosoma Mansoni. Parasitol. Res. 2013, 112, 3137–3149. [Google Scholar] [CrossRef]
- Kadri, D.; Crater, A.K.; Lee, H.; Solomon, V.R.; Ananvoranich, S. The Potential of Quinoline Derivatives for the Treatment of Toxoplasma Gondii Infection. Exp. Parasitol. 2014, 145, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.C.; Lage, L.M.d.R.; Lage, D.P.; Martins, V.T.; Carvalho, A.M.R.S.; Roatt, B.M.; Menezes-Souza, D.; Tavares, C.A.P.; Alves, R.J.; Barichello, J.M.; et al. Treatment of Murine Visceral Leishmaniasis Using an 8-Hydroxyquinoline-Containing Polymeric Micelle System. Parasitol. Int. 2016, 65, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Tavares, G.S.V.; Mendonça, D.V.C.; Pereira, I.A.G.; Oliveira-da-Silva, J.A.; Ramos, F.F.; Lage, D.P.; Machado, A.S.; Carvalho, L.M.; Reis, T.A.R.; Perin, L.; et al. A Clioquinol-Containing Pluronic® F127 Polymeric Micelle System Is Effective in the Treatment of Visceral Leishmaniasis in a Murine Model. Parasite 2020, 27, 63–72. [Google Scholar] [CrossRef]
- Lage, L.M.d.R.; Barichello, J.M.; Lage, D.P.; Mendonça, D.V.C.; Carvalho, A.M.R.S.; Rodrigues, M.R.; Menezes-Souza, D.; Roatt, B.M.; Alves, R.J.; Tavares, C.A.P.; et al. An 8-Hydroxyquinoline-Containing Polymeric Micelle System Is Effective for the Treatment of Murine Tegumentary Leishmaniasis. Parasitol. Res. 2016, 115, 4083–4095. [Google Scholar] [CrossRef] [PubMed]
- Chanmol, W.; Siriyasatien, P.; Intakhan, N. In Vitro Anti-Leishmania Activity of 8-Hydroxyquinoline and Its Synergistic Effect with Amphotericin B Deoxycholate against Leishmania Martiniquensis. PeerJ 2022, 10, 12813. [Google Scholar] [CrossRef]
- Sousa-Franco, J.; Araújo-Mendes, É.; Silva-Jardim, I.; Jane, L.; Faria, D.R.; Dutra, W.O.; Horta, M.D.F. Infection-Induced Respiratory Burst in BALB/c Macrophages Kills Leishmania Guyanensis Amastigotes through Apoptosis: Possible Involvement in Resistance to Cutaneous Leishmaniasis. Microbes Infect. 2006, 8, 390–400. [Google Scholar] [CrossRef]
- Mukbel, R.M.; Patten, C.; Gibson, K.; Ghosh, M.; Petersen, C.; Jones, D.E. Macrophage Killing of Leishmania Amazonensis Amastigotes Requires Both Nitric Oxide and Superoxide. Am. J. Trop. Med. Hyg. 2007, 76, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, E.S.; Antinarelli, L.M.R.; Silva, N.P.; Souza, I.O.; Meinel, R.S.; Rocha, M.N.; Soares, R.P.P.; da Silva, A.D. Quinoline Derivatives: Synthesis, Leishmanicidal Activity and Involvement of Mitochondrial Oxidative Stress as Mechanism of Action. Chem. Biol. Interact. 2016, 260, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, D.; Granato, J.T.; Macedo, G.C.; Dib, P.R.B.; Piplani, S.; Fung, J.; Da Silva, A.D.; Coimbra, E.S.; Petrovsky, N.; Salunke, D.B. Toll-like Receptor-7/8 Agonist Kill Leishmania Amazonensis by Acting as pro-Oxidant and pro-Inflammatory Agent. J. Pharm. Pharmacol. 2021, 73, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Torres Suarez, E.; Granados-Falla, D.S.; Robledo, S.M.; Murillo, J.; Upegui, Y.; Delgado, G. Antileishmanial Activity of Synthetic Analogs of the Naturally Occurring Quinolone Alkaloid N-Methyl-8-Methoxyflindersin. PLoS ONE 2020, 15, 0243392. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Serrato, M.A.; Salinas-Carmona, M.C.; Limón-Flores, A.Y. Immune Response to Leishmania Mexicana: The Host-Parasite Relationship. Pathog. Dis. 2020, 78, 60. [Google Scholar] [CrossRef]
- Passero, L.F.D.; Carvalho, A.K.; Bordon, M.L.; Bonfim-Melo, A.; Carvalho, K.; Kallás, E.G.; Santos, B.B.; Toyama, M.H.; Paes-Leme, A.; Corbett, C.E.; et al. Proteins of Leishmania (Viannia) Shawi Confer Protection Associated with Th1 Immune Response and Memory Generation. Parasites Vectors 2012, 5, 64. [Google Scholar] [CrossRef]
- Ribeiro Antinarelli, L.M.; Glanzmann, N.; Mendonça, D.V.C.; Lage, D.P.; Oliveira-da-Silva, J.A.; Tavares, G.S.V.; Carvalho, A.M.R.S.; Freitas, C.S.; Martins, V.T.; Duarte, M.C.; et al. Parasitological and Immunological Evaluation of a Quinoline Derivative Salt Incorporated into a Polymeric Micelle Formulation against Leishmania Infantum Infection. Parasitol. Res. 2022, 121, 2129–2140. [Google Scholar] [CrossRef]
- Yamamoto, E.S.; de Jesus, J.A.; Bezerra-Souza, A.; Brito, J.R.; Lago, J.H.G.; Laurenti, M.D.; Passero, L.F.D. Tolnaftate Inhibits Ergosterol Production and Impacts Cell Viability of Leishmania sp. Bioorg. Chem. 2020, 102, 104056. [Google Scholar] [CrossRef]
- Jesus, J.A.; Laurenti, M.D.; Lopes-Silva, M.; Ghilardi Lago, J.H.; Domingues Passero, L.F. Leishmanicidal Activity and Ultrastructural Changes of Maslinic Acid Isolated from Hyptidendron Canum. Evid.-Based Complement. Altern. Med. 2021, 2021. [Google Scholar] [CrossRef]
- Jesus, J.A.; Da Silva, T.N.F.; Yamamoto, E.S.; Lago, J.H.G.; Laurenti, M.D.; Passero, L.F.D. Ursolic Acid Potentializes Conventional Therapy in Experimental Leishmaniasis. Pathogens 2020, 9, 855. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC50 (μg/mL) | CC50 (μg/mL) | ||||||
|---|---|---|---|---|---|---|---|
| (Selective Index—SI) | |||||||
| Promastigote Forms | Amastigote Forms | Macrophages | |||||
| 24 h | 72 h | 24 h | 72 h | 24 h | 72 h | ||
| L. (L.) amazonensis | 8-HQ | 2.9 ± 0.3 | 1.1 ± 0.1 | 1.9 ± 0.1 | 0.9 ± 0.09 | ||
| SI (12.5) | SI (30.5) | SI (19.1) | SI (37.3) | ||||
| Milt | 8.5 ± 0.4 | 16.1 ± 0.1 | 6.5 ± 0.06 | 3.5 ± 0.2 | |||
| SI (5.0) | SI (2.0) | SI (6.6) | SI (9.3) | ||||
| L. (L.) infantum chagasi | 8-HQ | 2.1 ± 0.2 | 0.34 ± 0.1 | 2.0 ± 0.8 | 0.05 ± 0.001 | ||
| SI (17.3) | SI (98.8) | SI (18.1) | SI (672) | ||||
| Milt | 19.3 ± 1.6 | 7.5 ± 1.5 | 4.1 ± 0.2 | 0.05 ± 0.01 | |||
| SI (2.2) | SI (4.4) | SI (10.4) | SI (656) | ||||
| L. (V.) guyanensis | 8-HQ | 0.3 ± 0.08 | 0.1 ± 0.03 | 0.8 ± 0.1 | 0.03 ± 0.002 | ||
| SI (121) | SI (336) | SI (45.3) | SI (1120) | ||||
| Milt | 5.0 ± 1.3 | 1.8 ± 0.63 | 4.9 ± 1.4 | 0.8 ± 0.2 | |||
| SI (8.6) | SI (18.2) | SI (8.8) | SI (41) | ||||
| L. (V.) lainsoni | 8-HQ | 0.6 ± 0.1 | 0.06 ± 0.01 | 0.1 ± 0.09 | 0.5 ± 0.01 | ||
| SI (60.5) | SI (560) | SI (363) | SI (67.2) | ||||
| Milt | 5.3 ± 0.1 | 2.4 ± 0.2 | 0.5 ± 0.03 | 1.8 ± 1.04 | |||
| SI (8.1) | SI (13.7) | SI (85.8) | SI (18.2) | ||||
| L. (V.) naiffi | 8-HQ | 0.8 ± 0.1 | 0.5 ± 0.08 | 0.45 ± 0.02 | 0.03 ± 0.0002 | ||
| SI (45.4) | SI (67.2) | SI (80.6) | SI (1120) | ||||
| Milt | 23.5 ± 5.7 | 13.7 ± 0.5 | 2.0 ± 0.3 | 0.1 ± 0.08 | |||
| SI (1.8) | SI (2.4) | SI (21.4) | SI (328) | ||||
| L. (V.) shawi | 8-HQ | 0.2 ± 0.03 | 0.31 ± 0.08 | 0.1 ± 0.01 | 0.2 ± 0.001 | ||
| SI (181) | SI (108.4) | SI (363) | SI (168) | ||||
| Milt | 1.7 ± 0.09 | 2.4 ± 0.69 | 3.9 ± 0.4 | 1.6 ± 0.5 | |||
| SI (25.2) | SI (13.7) | SI (11) | SI (20.5) | ||||
| Host cell | 8-HQ | 36.3 ± 2.7 | 33.6 ± 2.2 | ||||
| Milt | 42.9 ± 1.3 | 32.8 ± 12.0 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, S.K.S.d.; Jesus, J.A.; Raminelli, C.; Laurenti, M.D.; Passero, L.F.D. High Selectivity of 8-Hydroxyquinoline on Leishmania (Leishmania) and Leishmania (Viannia) Species Correlates with a Potent Therapeutic Activity In Vivo. Pharmaceuticals 2023, 16, 707. https://doi.org/10.3390/ph16050707
Lima SKSd, Jesus JA, Raminelli C, Laurenti MD, Passero LFD. High Selectivity of 8-Hydroxyquinoline on Leishmania (Leishmania) and Leishmania (Viannia) Species Correlates with a Potent Therapeutic Activity In Vivo. Pharmaceuticals. 2023; 16(5):707. https://doi.org/10.3390/ph16050707
Chicago/Turabian StyleLima, Sarah Kymberly Santos de, Jéssica Adriana Jesus, Cristiano Raminelli, Márcia Dalastra Laurenti, and Luiz Felipe Domingues Passero. 2023. "High Selectivity of 8-Hydroxyquinoline on Leishmania (Leishmania) and Leishmania (Viannia) Species Correlates with a Potent Therapeutic Activity In Vivo" Pharmaceuticals 16, no. 5: 707. https://doi.org/10.3390/ph16050707