
Automated Synthesis of [18F]Flumazenil Application in GABAA Receptor Neuroimaging Availability for Rat Model of Anxiety

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
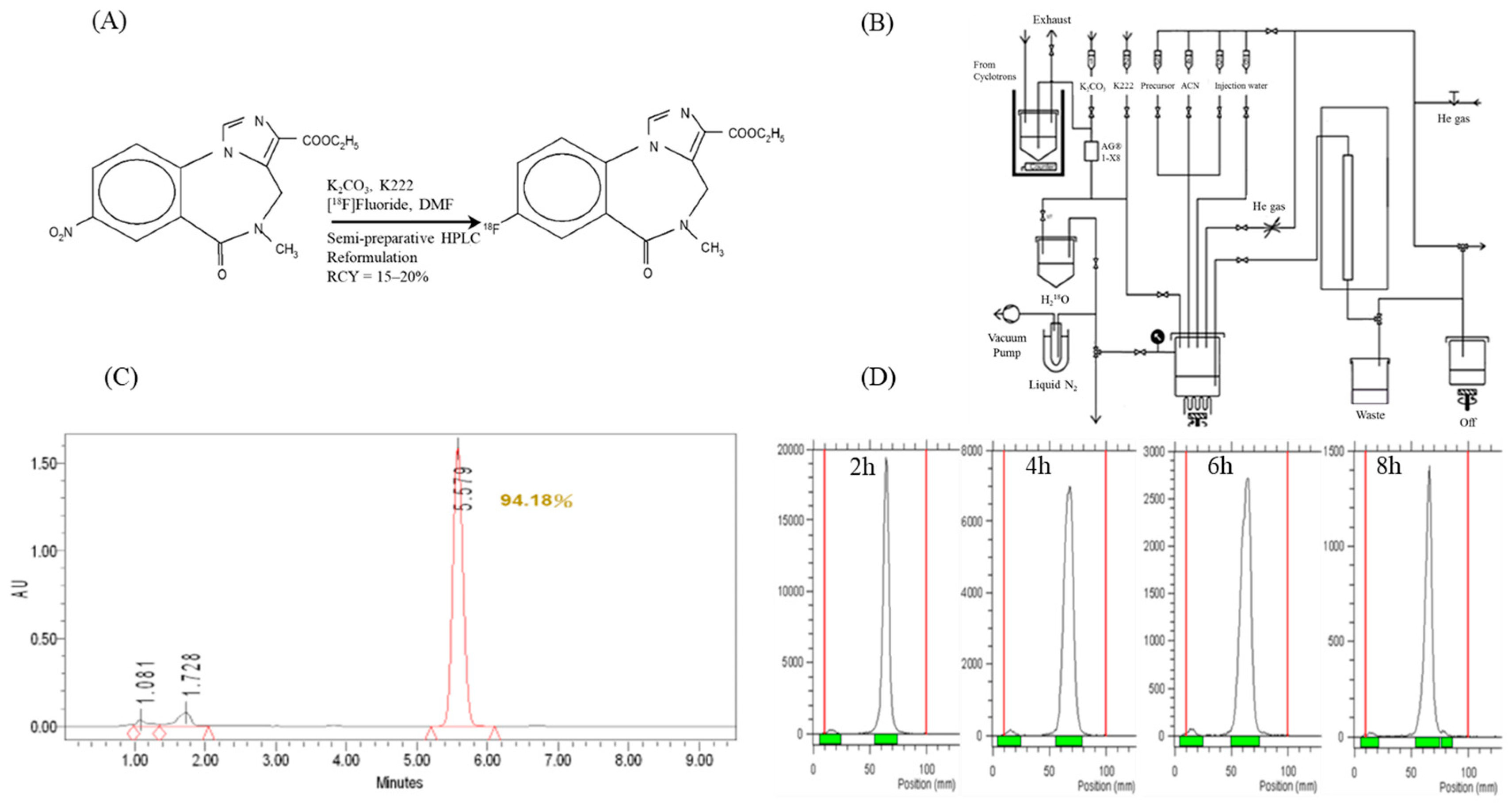
2.1. [18F]Flumazenil Radiosynthesis and Quality Controls
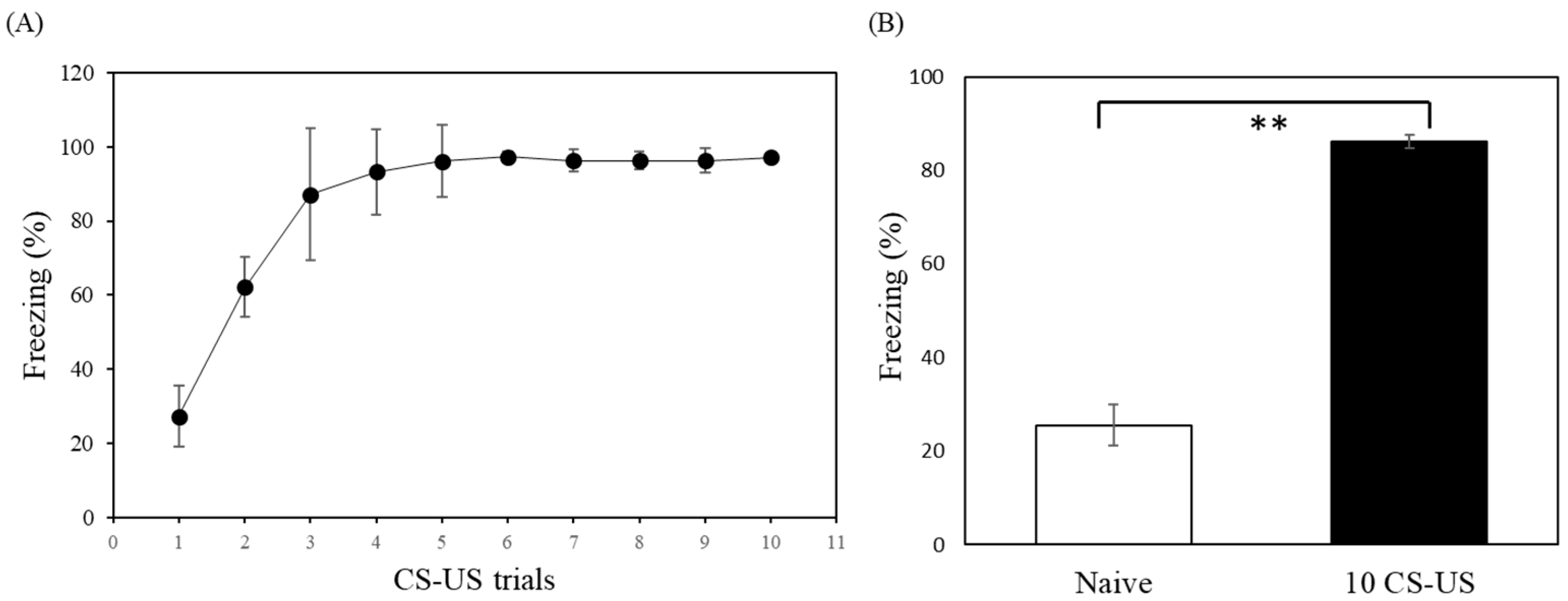
2.2. Behavioral Study
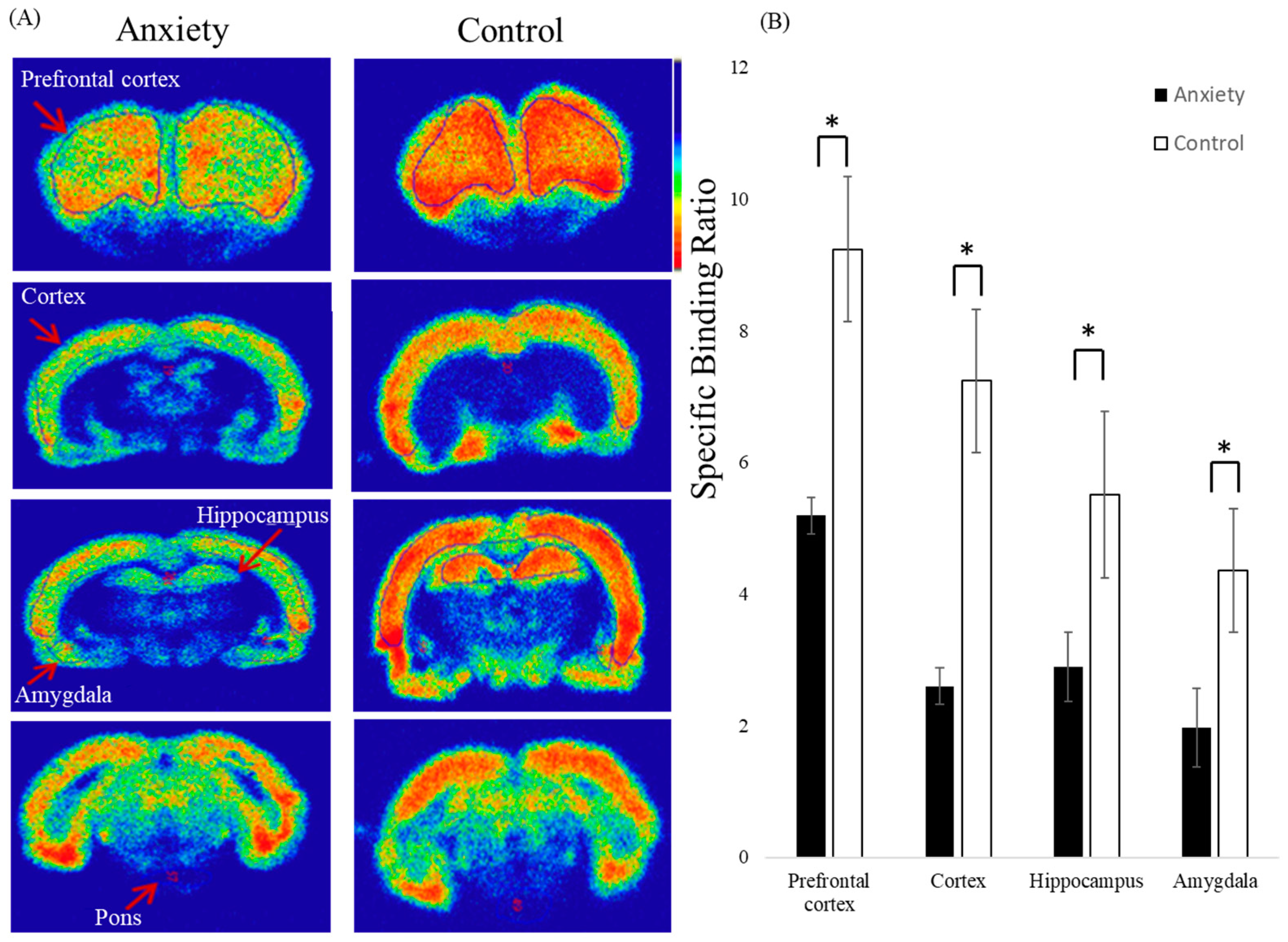
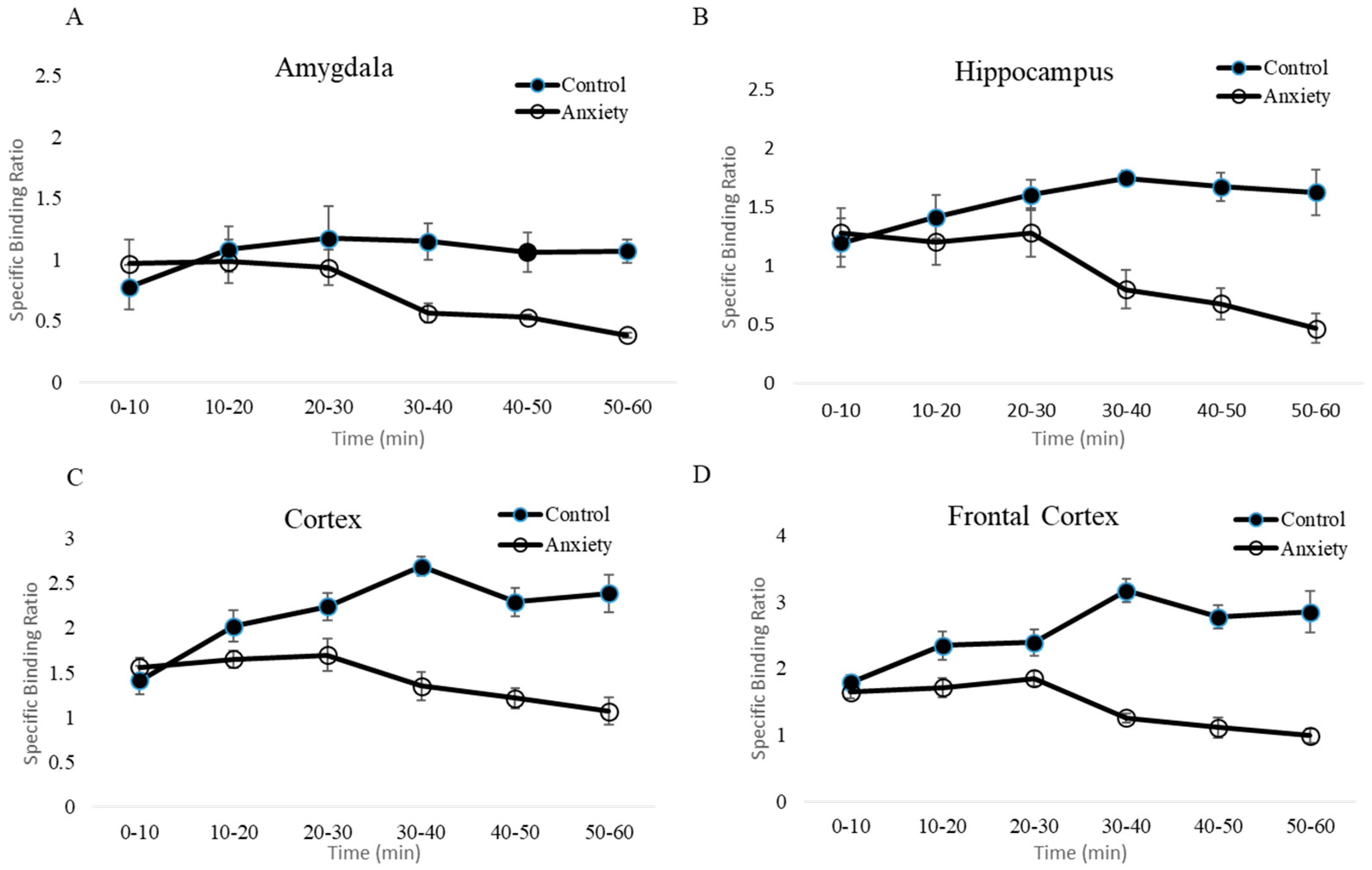
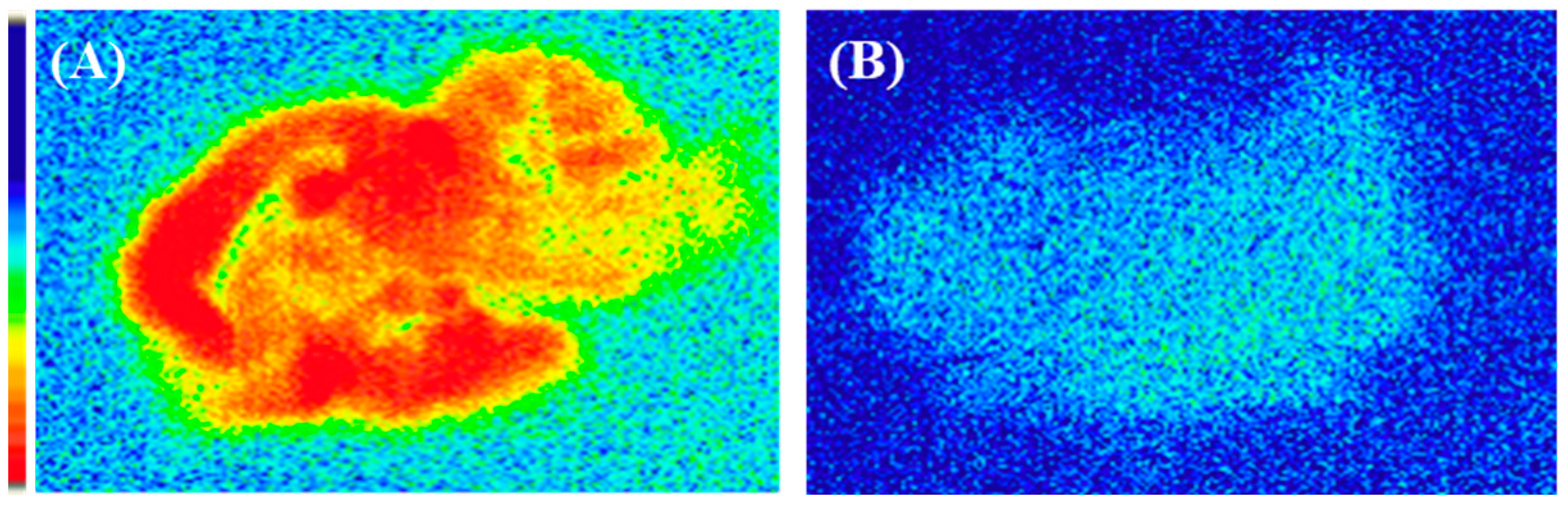
2.3. Ex Vivo Autoradiography
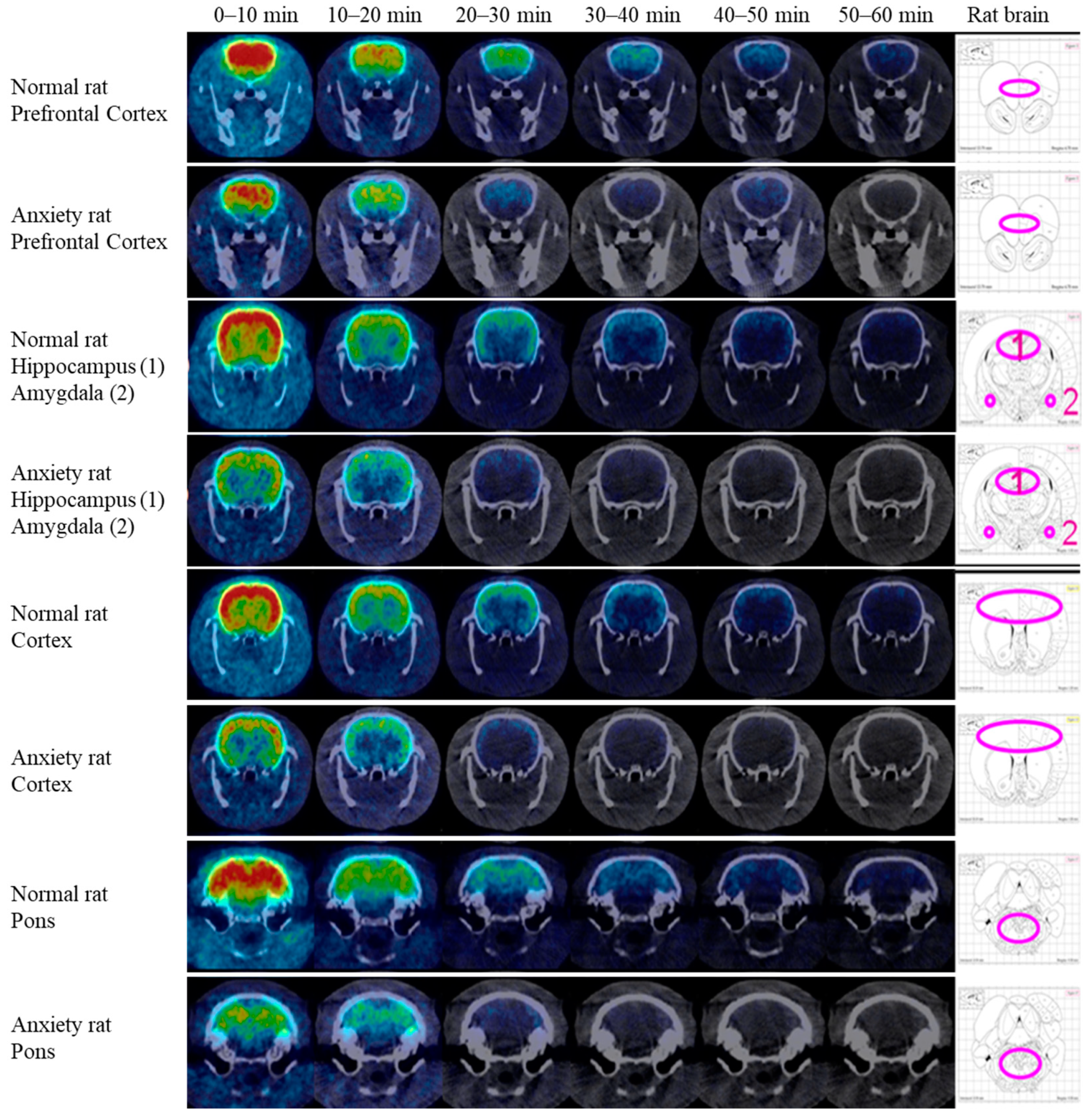
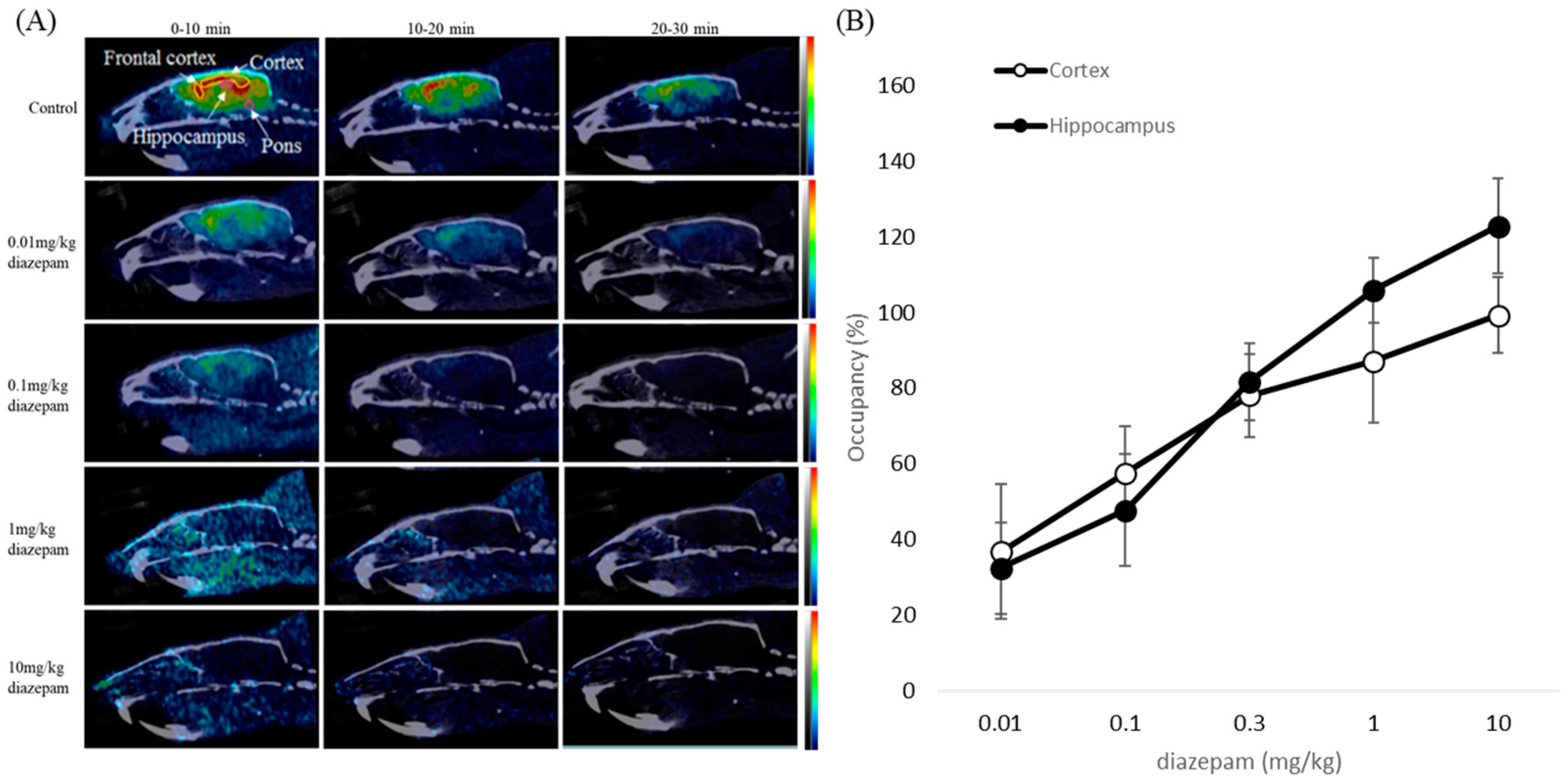
2.4. NanoPET/CT Imaging
2.5. Blocking Study
3. Discussion
4. Materials and Methods
4.1. [18F]Flumazenil Radiosynthesis
4.2. Animals
4.3. Ex Vivo Autoradiography
4.4. NanoPET/CT Image Acquisition and Analysis
4.5. Blocking Study
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nutt, D.J.; Malizia, A.L. New insights into the role of the GABAA-benzodiazepine receptor in psychiatric disorder. Br. J. Psychiatry 2001, 179, 390–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Noviello, C.M.; Teng, J.; Walsh, R.M.; Jr Kim, J.J.; Hibbs, R.E. Structure of a human synaptic GABA A receptor. Nature 2018, 559, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Nutt, D.J. Overview of diagnosis and drug treatments of anxiety disorders. CNS Spectr. 2005, 10, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Valentine, K.E.; Milling, L.S.; Clark, L.J.; Moriarty, C.L. The Efficacy of Hypnosis as a Treatment For Anxiety: A Meta-Analysis. Int. J. Clin. Exp. Hypn. 2019, 67, 336–363. [Google Scholar] [CrossRef]
- Babaev, O.; Chatain, C.P.; Krueger-Burg, D. Inhibition in the amygdala anxiety circuitry. Exp. Mol. Med. 2018, 50, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kalueff, A.V.; Nutt, D.J. Role of GABA in anxiety and depression. Depress. Anxiety 2007, 24, 495–517. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Kustrin, E.; Gegechkori, V.; Morton, D.W. Anxiolytic Terpenoids and Aromatherapy for Anxiety and Depression. Adv. Exp. Med. Biol. 2020, 1260, 283–296. [Google Scholar]
- Steimer, T. Animal models of anxiety disorders in rats and mice: Some conceptual issues. Dialogues Clin. Neurosci. 2011, 13, 495–506. [Google Scholar] [CrossRef]
- Delgado, M.R.; Olsson, A.; Phelps, E.A. Extending animal models of fear conditioning to humans. Biol. Psychol. 2006, 73, 39–48. [Google Scholar] [CrossRef]
- Toth, I.; Neumann, I.D. Animal models of social avoidance and social fear. Cell Tissue Res. 2013, 354, 107–118. [Google Scholar] [CrossRef]
- Ganella, D.E.; Kim, J.H. Developmental rodent models of fear and anxiety: From neurobiology to pharmacology. Br. J. Pharmacol. 2014, 171, 4556–4574. [Google Scholar] [CrossRef] [Green Version]
- Lezak, K.R.; Missig, G.; Carlezon, W.A., Jr. Behavioral methods to study anxiety in rodents. Dialogues Clin. Neurosci. 2017, 19, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Anagnostaras, S.G.; Wood, S.C.; Shuman, T.; Cai, D.J.; LeDuc, A.D.; Zurn, K.R.; Zurn, J.B.; Sage, J.R.; Herrera, G.M. Automated assessment of pavlovian conditioned freezing and shock reactivity in mice using the video freeze system. Front. Behav. Neurosci. 2010, 4, 158. [Google Scholar] [CrossRef] [Green Version]
- Maren, S. Neurobiology of Pavlovian fear conditioning. Annu. Rev. Neurosci. 2001, 24, 897–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponder, C.A.; Kliethermes, C.L.; Drew, M.; Muller, J.; Das, K.; Risbrough, V.B.; Crabbe, J.C.; Gilliam, T.C.; Palmer, A.A. Selection for contextual fear conditioning affects anxiety-like behaviors and gene expression. Genes Brain Behav. 2007, 6, 736–749. [Google Scholar] [CrossRef]
- Lee, H.-J.; Kim, J.-S.; Moon, C.; Son, Y. Profiling of gene expression in the brain associated with anxiety-related behaviors in the chronic phase following cranial irradiation. Sci. Rep. 2022, 12, 13162. [Google Scholar] [CrossRef]
- Wong, C.G.; Bottiglieri, T.; Snead, O.C., 3rd. GABA, gamma-hydroxybutyric acid, and neurological disease. Ann. Neurol. 2003, 54 (Suppl. S6), S3–S12. [Google Scholar] [CrossRef]
- Brambilla, P.; Perez, J.; Barale, F.; Schettini, G.; Soares, J.C. GABAergic dysfunction in mood disorders. Mol. Psychiatry 2003, 8, 715, 721–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herde, A.M.; Benke, D.; Ralvenius, W.T.; Mu, L.; Schibli, R.; Zeilhofer, H.U.; Krämer, S.D. GABA A receptor subtypes in the mouse brain: Regional mapping and diazepam receptor occupancy by in vivo [18F]flumazenil PET. Neuroimage 2017, 150, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Chua, H.C.; Chebib, M. GABA A Receptors and the Diversity in their Structure and Pharmacology. Adv. Pharmacol. 2017, 79, 1–34. [Google Scholar]
- Watanabe, M.; Maemura, K.; Kanbara, K.; Tamayama, T.; Hayasaki, H. GABA and GABA receptors in the central nervous system and other organs. Int. Rev. Cytol. 2002, 213, 1–47. [Google Scholar] [PubMed]
- Kraguljac, N.V.; McDonald, W.M.; Widge, A.S.; Rodriguez, C.I.; Tohen, M.; Nemeroff, C.B. Neuroimaging Biomarkers in Schizophrenia. Am. J. Psychiatry. 2021, 178, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Giusti, P.; Guidetti, G.; Costa, E.; Guidotti, A. The preferential antagonism of pentylenetetrazole proconflict responses differentiates a class of anxiolytic benzodiazepines with potential antipanic action. J. Pharmacol. Exp. Ther. 1991, 257, 1062–1068. [Google Scholar] [PubMed]
- Kim, W.; Park, H.S.; Moon, B.S.; Lee, B.C.; Kim, S.E. PET measurement of “GABA shift” in the rat brain: A preclinical application of bolus plus constant infusion paradigm of [18F]flumazenil. Nucl. Med. Biol. 2017, 45, 30–34. [Google Scholar] [CrossRef]
- Vivash, L.; Gregoire, M.-C.; Bouilleret, V.; Berard, A.; Wimberley, C.; Binns, D.; Roselt, P.; Katsifis, A.; Myers, D.; Hicks, R.J.; et al. In vivo measurement of hippocampal GABAA/cBZR density with [18F]-flumazenil PET for the study of disease progression in an animal model of temporal lobe epilepsy. PLoS ONE 2014, 9, e86722. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, H.; Kim, C.H.; Lee, G.Y.; Baik, H.J.; Han, J.I. Effect of flumazenil on recovery from anesthesia and the bispectral index after sevoflurane/fentanyl general anesthesia in unpremedicated patients. Korean J. Anesthesiol. 2012, 62, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Mosier, D.M.; Mosier, N.F. Flumazenil: A new benzodiazepine antagonist. Plast. Reconstr. Surg. 1993, 91, 194–195. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, J. Flumazenil. Masui 2013, 62, 10–18. [Google Scholar]
- Yabré, M.; Ferey, L.; Somé, I.T.; Gaudin, K. Greening Reversed-Phase Liquid Chromatography Methods Using Alternative Solvents for Pharmaceutical Analysis. Molecules 2018, 23, 1065. [Google Scholar] [CrossRef] [Green Version]
- Dedeurwaerdere, S.; Gregoire, M.-C.; Vivash, L.; Roselt, P.; Binns, D.; Fookes, C.; Greguric, I.; Pham, T.; Loc’H, C.; Katsifis, A.; et al. In-vivo imaging characteristics of two fluorinated flumazenil radiotracers in the rat. Eur. J. Nucl. Med. Mol. Imaging 2009, 36, 958–965. [Google Scholar] [CrossRef]
- Ghosh, K.; Padmanabhan, P.; Yang, C.-T.; Wang, Z.; Palanivel, M.; Ng, K.; Lu, J.; Carlstedt-Duke, J.; Halldin, C.; Gulyás, B. An In Vivo Study of a Rat Fluid-Percussion-Induced Traumatic Brain Injury Model with [11C]PBR28 and [18F]flumazenil PET Imaging. Int. J. Mol. Sci. 2021, 22, 951. [Google Scholar] [CrossRef] [PubMed]
- Vivash, L.; Gregoire, M.C.; Lau, E.W.; Ware, R.E.; Binns, D.; Roselt, P.; Bouilleret, V.; Myers, D.E.; Cook, M.J.; Hicks, R.J.; et al. 18F-flumazenil: A γ-aminobutyric acid A-specific PET radiotracer for the localization of drug-resistant temporal lobe epilepsy. J. Nucl. Med. 2013, 54, 1270–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Fougère, C.; Grant, S.; Kostikov, A.; Schirrmacher, R.; Gravel, P.; Schipper, H.M.; Reader, A.; Evans, A.; Thiel, A. Where in-vivo imaging meets cytoarchitectonics: The relationship between cortical thickness and neuronal density measured with high-resolution [18F]flumazenil-PET. Neuroimage 2011, 56, 951–960. [Google Scholar] [CrossRef]
- Janak, H.; Tye, K.M. From circuits to behaviour in the amygdala. Nature 2015, 517, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Jung, M.W. Neural circuits and mechanisms involved in Pavlovian fear conditioning: A critical review. Neurosci. Biobehav. Rev. 2006, 30, 188–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelofs, K. Freeze for action: Neurobiological mechanisms in animal and human freezing. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160206. [Google Scholar] [CrossRef] [Green Version]
- Crestani, F.; Lorez, M.; Baer, K.; Essrich, C.; Benke, D.; Laurent, J.P.; Belzung, C.; Fritschy, J.M.; Lüscher, B.; Mohler, H. Decreased GABA A -receptor clustering results in enhanced anxiety and a bias for threat cues. Nat. Neurosci. 1999, 2, 833–839. [Google Scholar] [CrossRef]
- Collins, S.A.; Ninan, I. Development-Dependent Plasticity in Vasoactive Intestinal Polypeptide Neurons in the Infralimbic Cortex. Cereb. Cortex Commun. 2021, 2, tgab007. [Google Scholar] [CrossRef]
- Luyten, L.; Casteels, C.; Vansteenwegen, D.; van Kuyck, K.; Koole, M.; Van Laere, K.; Nuttin, B. Micro-positron emission tomography imaging of rat brain metabolism during expression of contextual conditioning. J. Neurosci. 2012, 32, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Mandap, K.S.; Ido, T.; Kiyono, Y.; Kobayashi, M.; Lohith, T.G.; Mori, T.; Kasamatsu, S.; Kudo, T.; Okazawa, H.; Fujibayashi, Y. Development of microwave-based automated nucleophilic [18F]fluorination system and its application to the production of [18F]flumazenil. Nucl. Med. Biol. 2009, 36, 403–409. [Google Scholar] [CrossRef]
- Rodnick, M.E.; Hockley, B.G.; Sherman, P.; Quesada, C.; Battle, M.R.; Jackson, A.; Linder, K.E.; Macholl, S.; Trigg, W.J.; Kilbourn, M.R.; et al. Novel fluorine-18 PET radiotracers based on flumazenil for GABA A imaging in the brain. Nucl. Med. Biol. 2013, 40, 901–905. [Google Scholar] [CrossRef] [Green Version]
- Moon, B.S.; Park, J.H.; Lee, H.J.; Lee, B.C.; Kim, S.E. Routine production of [18F]flumazenil from iodonium tosylate using a sample pretreatment method: A 2.5-year production report. Mol. Imaging Biol. 2014, 16, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Ryzhikov, N.N.; Seneca, N.; Krasikova, R.N.; Gomzina, N.A.; Shchukin, E.; Fedorova, O.S.; Vassiliev, D.A.; Gulyás, B.; Hall, H.; Savic, I.; et al. Preparation of highly specific radioactivity [18F]flumazenil and its evaluation in cynomolgus monkey by positron emission tomography. Nucl. Med. Biol. 2005, 32, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Chugani, H.T. Imaging Brain Metabolism in the Newborn. J. Child Neurol. 2018, 33, 851–860. [Google Scholar] [CrossRef]
- Massaweh, G.; Schirrmacher, E.; la Fougere, C.; Kovacevic, M.; Wängler, C.; Jolly, D.; Gravel, P.; Reader, A.J.; Thiel, A.; Schirrmacher, R. Improved work-up procedure for the production of [18F]flumazenil and first results of its use with a high-resolution research tomograph in human stroke. Nucl. Med. Biol. 2009, 36, 721–727. [Google Scholar] [CrossRef]
- Heiss, W.D. Radionuclide imaging in ischemic stroke. J. Nucl. Med. 2014, 55, 1831–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odano, I.; Halldin, C.; Karlsson, P.; Varrone, A.; Airaksinen, A.; Krasikova, R.N.; Farde, L. [18F]flumazenil binding to central benzodiazepine receptor studies by PET—Quantitative analysis and comparisons with [11C]flumazenil. Neuroimage 2009, 45, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Alagille, D.; Tamagnan, G.; Price, R.R.; Baldwin, R.M. Microwave-induced nucleophilic [18F]fluorination on aromatic rings: Synthesis and effect of halogen on [18F]fluoride substitution of meta-halo (F, Cl, Br, I)-benzonitrile derivatives. Appl. Radiat. Isot. 2008, 66, 1396–1402. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, K.; Miura, Y.; Matsumura, K.; Kanemasa, Y.; Nakamichi, K.; Saijo, M.; Toyohara, J.; Ishii, K. PET Imaging of 18F-FDG, 11C-methionine, 11C-flumazenil, and 11C-4DST in Progressive Multifocal Leukoencephalopathy. Intern. Med. 2017, 56, 1219–1223. [Google Scholar] [CrossRef] [Green Version]
- Kuchar, M.; Mamat, C. Methods to Increase the Metabolic Stability of 18F-Radiotracers. Molecules 2015, 20, 16186–16220. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.E.; Squire, L.R. Classical conditioning and brain systems: The role of awareness. Science 1998, 280, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Leech, R.; Sharp, D.J. The role of the posterior cingulate cortex in cognition and disease. Brain 2014, 137 Pt 1, 12–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuette, S.R.; Hobson, S. Conditioned contextual fear memory to assess natural forgetting and cognitive enhancement in rats. J. Biol. Methods 2018, 5, e99. [Google Scholar] [CrossRef]
- Siegmund, A.; Wotjak, C.T. A mouse model of posttraumatic stress disorder that distinguishes between conditioned and sensitised fear. J. Psychiatr. Res. 2007, 41, 848–860. [Google Scholar] [CrossRef]
- Curzon, P.; Rustay, N.R.; Browman, K.E. Frontiers in Neuroscience Cued and Contextual Fear Conditioning for Rodents. In Methods of Behavior Analysis in Neuroscience; Buccafusco, J.J., Ed.; Taylor & Francis Group, LLC.: Boca Raton, FL, USA, 2009. [Google Scholar]
- Lin, H.C.; Mao, S.C.; Gean, P.W. Block of gamma-aminobutyric acid-A receptor insertion in the amygdala impairs extinction of conditioned fear. Biol. Psychiatry 2009, 66, 665–673. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farn, S.-S.; Cheng, K.-H.; Huang, Y.-R.; Lee, S.-Y.; Chen, J.-T.; Chang, K.-W. Automated Synthesis of [18F]Flumazenil Application in GABAA Receptor Neuroimaging Availability for Rat Model of Anxiety. Pharmaceuticals 2023, 16, 417. https://doi.org/10.3390/ph16030417
Farn S-S, Cheng K-H, Huang Y-R, Lee S-Y, Chen J-T, Chang K-W. Automated Synthesis of [18F]Flumazenil Application in GABAA Receptor Neuroimaging Availability for Rat Model of Anxiety. Pharmaceuticals. 2023; 16(3):417. https://doi.org/10.3390/ph16030417
Chicago/Turabian StyleFarn, Shiou-Shiow, Kai-Hung Cheng, Yuan-Ruei Huang, Shih-Ying Lee, Jenn-Tzong Chen, and Kang-Wei Chang. 2023. "Automated Synthesis of [18F]Flumazenil Application in GABAA Receptor Neuroimaging Availability for Rat Model of Anxiety" Pharmaceuticals 16, no. 3: 417. https://doi.org/10.3390/ph16030417