
Effect of Agave Fructan Bioconjugates on Metabolic Syndrome Parameters in a Murine Model

Abstract
:1. Introduction
2. Results and Discussion
2.1. Synthesis of Agave Fructan Bioconjugates
2.2. Effects of Acute Supplementation with Agave Fructan Bioconjugates on Metabolic Syndrome Parameters
2.2.1. Induction of Metabolic Syndrome with a Hypercaloric Diet
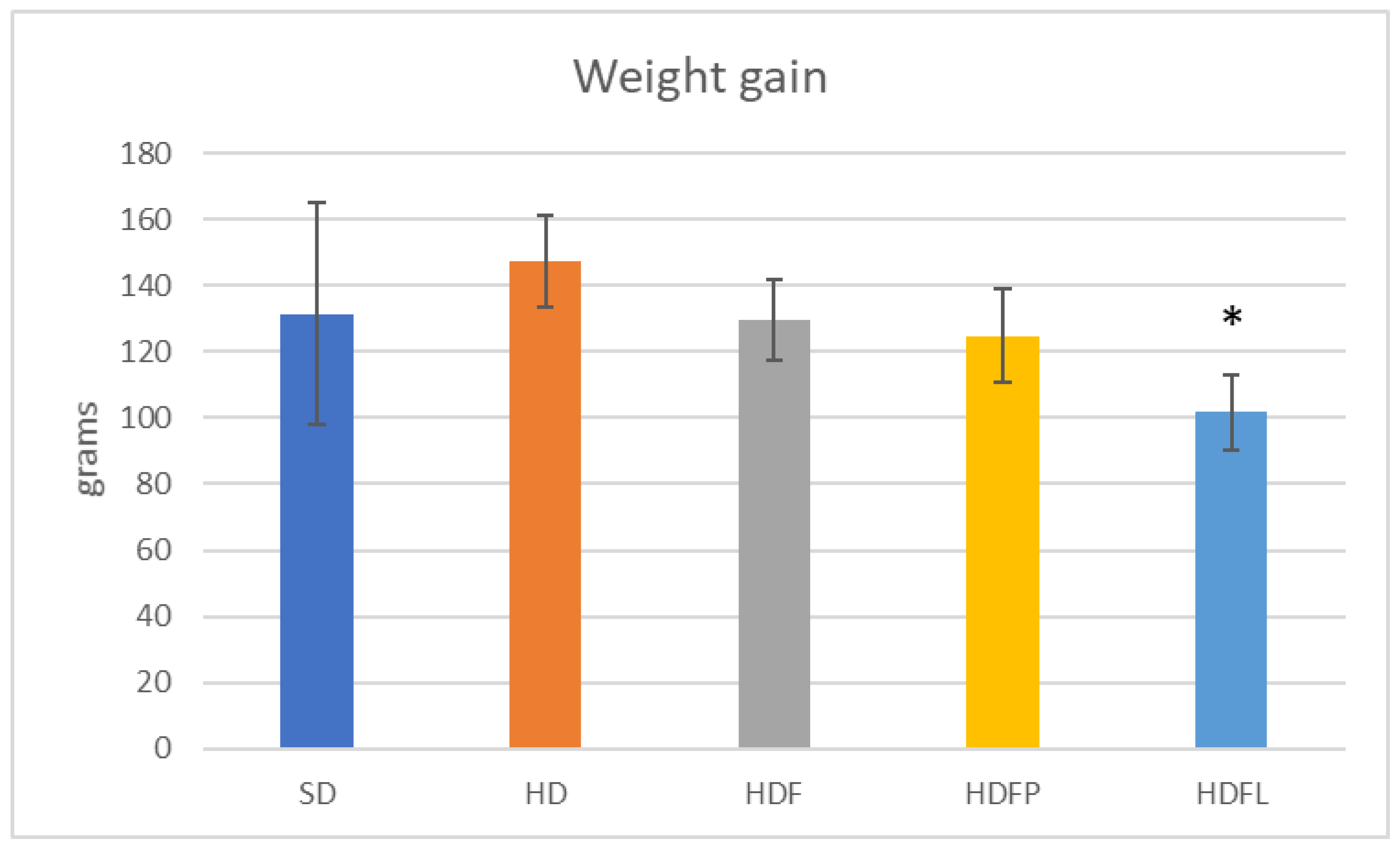
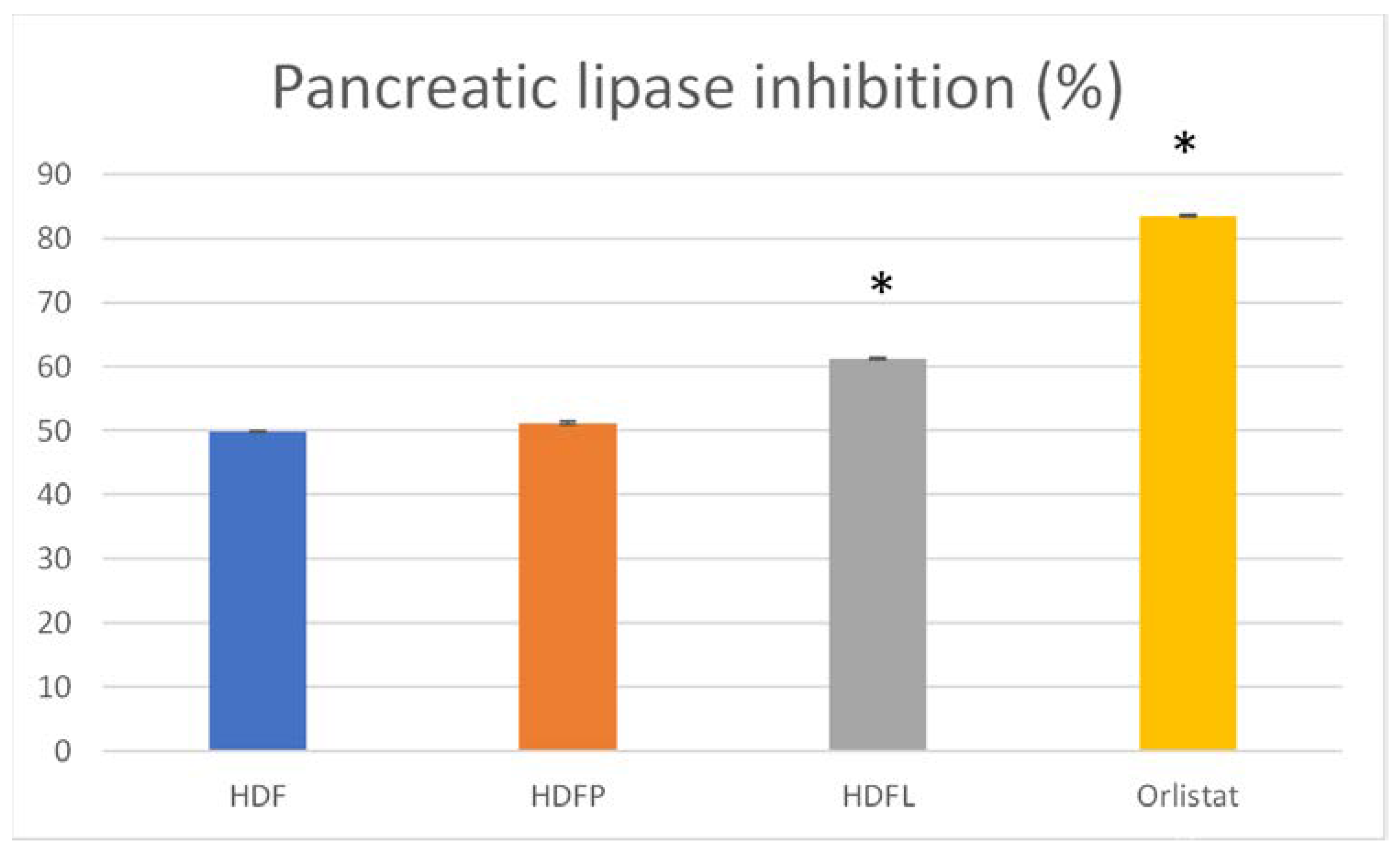
2.2.2. Metabolic Syndrome Prevention by Agave Fructan Bioconjugates
3. Materials and Methods
3.1. Synthesis of Agave Fructan Bioconjugates
3.2. Diet-Induced Metabolic Syndrome Model
3.2.1. Preparation of the Special Diet
3.2.2. Animals
3.2.3. Induction of Metabolic Syndrome and Treatment with Agave Fructans and Bioconjugates
3.2.4. In Vitro Study of Pancreatic Lipase Inhibition
4. Conclusions
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.-C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the Metabolic Syndrome. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Shan, X.; Dun, Y.; Cai, C.; Hao, J.; Li, G.; Cui, K.; Yu, G. Anti-Metabolic Syndrome Effects of Fucoidan from Fucus vesiculosus via Reactive Oxygen Species-Mediated Regulation of JNK, Akt, and AMPK Signaling. Molecules 2019, 24, 3319. [Google Scholar] [CrossRef] [Green Version]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Álvarez, H.; Bravo-Jiménez, A.; Martínez-Arellanes, M.; Gamboa-Osorio, G.O.; Chávez-Gutiérrez, E.; González-Hernández, L.A.; Gallardo-Ignacio, K.; Quintana-Romero, O.J.; Ariza-Castolo, A.; Guerra-Araiza, C.; et al. In Silico-Based Design and In Vivo Evaluation of an Anthranilic Acid Derivative as a Multitarget Drug in a Diet-Induced Metabolic Syndrome Model. Pharmaceuticals 2021, 14, 914. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, P.M.; Tuomilehto, J.; Rydén, L. The metabolic syndrome—What is it and how should it be managed? Eur. J. Prev. Cardiol. 2020, 26, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [Google Scholar] [CrossRef] [PubMed]
- Welty, F.K.; Alfaddagh, A.; Elajami, T.K. Targeting inflammation in metabolic syndrome. Transl. Res. 2016, 167, 257–280. [Google Scholar] [CrossRef]
- Samson, S.L.; Garber, A.J. Metabolic Syndrome. Endocrinol. Metab. Clin. N. Am. 2014, 43, 1–23. [Google Scholar] [CrossRef]
- Waltenberger, B.; Mocan, A.; Šmejkal, K.; Heiss, E.H.; Atanasov, A.G. Natural Products to Counteract the Epidemic of Cardiovascular and Metabolic Disorders. Molecules 2016, 21, 807. [Google Scholar] [CrossRef]
- Wang, X.-F.; Chen, X.; Tang, Y.; Wu, J.-M.; Qin, D.-L.; Yu, L.; Yu, C.-L.; Zhou, X.-G.; Wu, A.-G. The Therapeutic Potential of Plant Polysaccharides in Metabolic Diseases. Pharmaceuticals 2022, 15, 1329. [Google Scholar] [CrossRef]
- Arrizon, J.; Urias-Silvas, J.E.; Sandoval, G.; Mancilla-Margalli, N.A.; Gschaedler, A.C.; Morel, S.; Monsan, P. Production and Bioactivity of Fructan-Type Oligosaccharides. In Food Oligosaccharides: Production, Analysis and Bioactivity; Moreno, F.J., Sanz, M.L., Eds.; Wiley-Blackwell: Madrid, Spain, 2014; pp. 184–199. [Google Scholar]
- Lopez, M.G.; Mancilla-Margalli, N.A.; Mendoza-Diaz, G. Molecular Structures of Fructans from Agave tequilana Weber var. azul. J. Agric. Food Chem. 2003, 51, 7835–7840. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Aguirre, A.L.; Camacho-Ruíz, R.M.; Gutiérrez-Mercado, Y.K.; Padilla-Camberos, E.; González-Ávila, M.; Gálvez-Gastélum, F.J.; Díaz-Martínez, N.E.; Ortuño-Sahagún, D. Fructans from Agave tequilana with a Lower Degree of Polymerization Prevent Weight Gain, Hyperglycemia and Liver Steatosis in High-Fat Diet-Induced Obese Mice. Plant Foods Hum. Nutr. 2016, 71, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla-Camberos, E.; Barragán-Álvarez, C.P.; Diaz-Martinez, N.E.; Rathod, V.; Flores-Fernández, J.M. Effects of Agave fructans (Agave tequilana Weber var. azul) on Body Fat and Serum Lipids in Obesity. Plant Foods Hum. Nutr. 2018, 73, 34–39. [Google Scholar] [CrossRef]
- Regalado-Rentería, E.; Aguirre-Rivera, J.R.; Godínez-Hernández, C.I.; García-López, J.C.; Oros-Ovalle, A.C.; Martínez-Gutiérrez, F.; Martinez-Martinez, M.; Ratering, S.; Schnell, S.; Ruíz-Cabrera, M.Á.; et al. Effects of Agave Fructans, Inulin, and Starch on Metabolic Syndrome Aspects in Healthy Wistar Rats. ACS Omega 2020, 5, 10740–10749. [Google Scholar] [CrossRef] [PubMed]
- Urías-Silvas, J.E.; Cani, P.D.; Delmée, E.; Neyrinck, A.; López, M.G.; Delzenne, N.M. Physiological effects of dietary fructans extracted from Agave tequilana Gto. and Dasylirion spp. Br. J. Nutr. 2008, 99, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Bonsu, N.K.A.; Johnson, C.S.; Mcleod, K.M. Can dietary fructans lower serum glucose? J. Diabetes 2011, 3, 58–66. [Google Scholar] [CrossRef]
- Castillo Andrade, A.I.; Rivera Bautista, C.; Godínez Hernández, C.; Ruiz Cabrera, M.A.; Fuentes Ahumada, C.; García Chávez, E.; Grajales Lagunes, A. Physiometabolic effects of Agave salmiana fructans evaluated in Wistar rats. Int. J. Biol. Macromol. 2018, 108, 1300–1309. [Google Scholar] [CrossRef]
- Espinosa-Andrews, H.; Urías-Silvas, J.E.; Morales-Hernández, N. The role of agave fructans in health and food applications: A review. Trends Food Sci. Technol. 2021, 114, 585–598. [Google Scholar] [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [Green Version]
- Casas-Godoy, L.; Arrizon, J.; Arrieta-Baez, D.; Plou, F.J.; Sandoval, G. Synthesis and emulsifying properties of carbohydrate fatty acid esters produced from Agave tequilana fructans by enzymatic acylation. Food Chem. 2016, 204, 437–443. [Google Scholar] [CrossRef]
- Hernández-Pérez, S.; Oliart-Ros, R.M.; Casas-Godoy, L.; Sandoval, G.; Guarner-Lans, V.; Castrejón-Téllez, V.; Quevedo-Corona, L.; Peña-Montes, C.; Ramírez-Higuera, A. Beneficial Effects of Fructooligosaccharides Esterified with Lauric Acid in a Metabolic Syndrome Model Induced by a High-Fat and High-Carbohydrate Diet in Wistar Rats. J. Med. Food 2022, 25, 825–835. [Google Scholar] [CrossRef]
- Panchal, S.K.; Brown, L. Rodent Models for Metabolic Syndrome Research. J. Biomed. Biotechnol. 2011, 2011, 351982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Correa, E.; González-Pérez, I.; Clavel-Pérez, P.I.; Contreras-Vargas, Y.; Carvajal, K. Biochemical and nutritional overview of diet-induced metabolic syndrome models in rats: What is the best choice? Nutr. Diabetes 2020, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- van Kempen, S.E.H.J.; Boeriu, C.G.; Schols, H.A.; de Waard, P.; van der Linden, E.; Sagis, L.M.C. Novel surface-active oligofructose fatty acid mono-esters by enzymatic esterification. Food Chem. 2013, 138, 1884–1891. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Byrne, C.S.; Chambers, E.S.; Alhabeeb, H.; Chhina, N.; Morrison, D.J.; Preston, T.; Tedford, C.; Fitzpatrick, J.; Irani, C.; Busza, A.; et al. Increased colonic propionate reduces anticipatory reward responses in the human striatum to high-energy foods. Am. J. Clin. Nutr. 2016, 104, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, M.F.; DiNicolantonio, J.J. Lauric acid-rich medium-chain triglycerides can substitute for other oils in cooking applications and may have limited pathogenicity. Open Heart 2016, 3, e000467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garfinkel, M.; Lee, S.; Opara, E.C.; Akwari, O.E. Insulinotropic potency of lauric acid: A metabolic rationale for medium chain fatty acids (MCF) in TPN formulation. J. Surg. Res. 1992, 52, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, B.F.; Gosmann, G.; Zimmer, A.R. Modeling Diet-Induced Metabolic Syndrome in Rodents. Mol. Nutr. Food Res. 2020, 64, 2000249. [Google Scholar] [CrossRef]
- St-Onge, M.P.; Mayrsohn, B.; O’Keeffe, M.; Kissileff, H.R.; Choudhury, A.R.; Laferrère, B. Impact of medium and long chain triglycerides consumption on appetite and food intake in overweight men. Eur. J. Clin. Nutr. 2014, 68, 1134–1140. [Google Scholar] [CrossRef]
- Franco-Robles, E.; López, M.G. Agavins Increase Neurotrophic Factors and Decrease Oxidative Stress in the Brains of High-Fat Diet-Induced Obese Mice. Molecules 2016, 21, 998. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2017, 114, 1752–1761. [Google Scholar] [CrossRef]
- Hsueh, W.A.; Wyne, K. Renin-Angiotensin-Aldosterone System in Diabetes and Hypertension. J. Clin. Hypertens. 2011, 13, 224–237. [Google Scholar] [CrossRef]
- Cronin, P.; Joyce, S.A.; O’Toole, P.W.; O’Connor, E.M. Dietary Fibre Modulates the Gut Microbiota. Nutrients 2021, 13, 1655. [Google Scholar] [CrossRef]
- Nie, Y.; Luo, F. Dietary Fiber: An Opportunity for a Global Control of Hyperlipidemia. Oxidative Med. Cell. Longev. 2021, 2021, 5542342. [Google Scholar] [CrossRef]
- Rault-Nania, M.-H.; Demougeot, C.; Gueux, E.; Berthelot, A.; Dzimira, S.; Rayssiguier, Y.; Rock, E.; Mazur, A. Inulin supplementation prevents high fructose diet-induced hypertension in rats. Clin. Nutr. 2008, 27, 276–282. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, H.; Xiao, X.; Hu, L.; Xin, F.; Yu, X. Inulin-type fructan improves diabetic phenotype and gut microbiota profiles in rats. PeerJ 2018, 6, e4446. [Google Scholar] [CrossRef]
- Li, H.-Y.; Zhou, D.-D.; Gan, R.-Y.; Huang, S.-Y.; Zhao, C.-N.; Shang, A.; Xu, X.-Y.; Li, H.-B. Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review. Nutrients 2021, 13, 3211. [Google Scholar] [CrossRef]
- Sandoval, G.; Arrizon, J.; Gonalez-Avila, M.; Padilla-Camberos, E.; Matinez-Velazquez, M.; Villanueva-Rodriguez, S.; Casas-Godoy, L. Bioconjugate molecules with biological and techno-functional activity, method for the production thereof and use thereof. Patent Number MX 358789, 18 December 2013. [Google Scholar]
- Kim, J.; Lee, Y.S.; Kim, C.-S.; Kim, J.S. Betulinic Acid has an Inhibitory Effect on Pancreatic Lipase and Induces Adipocyte Lipolysis. Phytother. Res. 2012, 26, 1103–1106. [Google Scholar] [CrossRef]
- Filippatos, T.D.; Derdemezis, C.S.; Gazi, I.F.; Nakou, E.S.; Mikhailidis, D.P.; Elisaf, M.S. Orlistat-Associated Adverse Effects and Drug Interactions. Drug Saf. 2008, 31, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Campos-Valdez, A.R.; Casas-Godoy, L.; Sandoval, G.; Hernández, L.; Sassaki, G.L.; Alencar de Menezes, L.R.; Campos-Terán, J.; Reyes-Duarte, D.; Arrizon, J. Regioselective synthesis of 6″-O-lauroyl-1-kestose and 6‴-O-lauroylnystose by sequential enzymatic reactions of transfructosylation and acylation. Biocatal. Biotransform. 2022, 40, 133–143. [Google Scholar] [CrossRef]
- Harvey, D.J. Analysis of carbohydrates and glycoconjugates by matrix-assisted laser desorption/ionization mass spectrometry: An update for 2015–2016. Mass Spectrom. Rev. 2021, 40, 408–565. [Google Scholar] [CrossRef]
- Cheng, H.S.; Ton, S.H.; Phang, S.C.W.; Tan, J.B.L.; Abdul Kadir, K. Increased susceptibility of post-weaning rats on high-fat diet to metabolic syndrome. J. Adv. Res. 2017, 8, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Côme, M.; Ulmann, L.; Chini Zittelli, G.; Faraloni, C.; Nazih, H.; Ouguerram, K.; Chénais, B.; Mimouni, V. Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats. Nutrients 2019, 11, 1069. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Consumption 1/Diet 2 | SD | HD | HDF | HDFP | HDFL |
|---|---|---|---|---|---|
| Water (mL/d) | 23.48 | 16.74 | 14.89 | 13.59 | 12.66 |
| Feed (g/d) | 17.31 | 15.69 | 15.68 | 15.00 | 15.53 |
| Parameter 2/Diet 3 | SD | HD | HDF | HDFP | HDFL |
|---|---|---|---|---|---|
| HDL | 26.74 ± 5.02 | 28.43 ± 3.53 | 30.32 ± 5.64 | 27.69 ± 2.94 | 26.69 ± 2.45 |
| LDL | 22.71 ± 5.16 | 16.82 ± 7.37 | 9.02 ± 4.35 | 16.94 ± 6.00 | 16.07 ± 5.93 |
| Total cholesterol | 57.41 ± 10.17 | 53.95 ± 5.59 | 49.51 ± 5.81 | 56.86 ± 5.35 | 55.35 ± 5.07 |
| Triglycerides | 39.76 ± 13.25 | 43.45 ± 13.9 | 50.84 ± 16.07 | 61.19 ± 19.91 | 62.96 ± 19.91 * |
| Component | Percentage |
|---|---|
| Corn starch | 2.448 |
| Anhydrous butter (clarified) | 32.00 |
| Casein | 23.50 |
| Maltodextrin | 16.50 |
| Sucrose | 9.00 |
| Mineral premix AIN-93G | 4.59 |
| Soybean oil | 3.5 |
| Microcrystalline cellulose | 3.25 |
| Carboxymethyl cellulose | 3.25 |
| AIN-93 vitamin premix | 1.31 |
| L-cystine | 0.39 |
| Choline bitartrate | 0.26 |
| Butylhydroquinone | 0.002 |
| Macronutrient | SD 1 | HD |
|---|---|---|
| Fat | 13.6 | 58.9 |
| Carbohydrates | 60.0 | 26.25 |
| Proteins | 26.0 | 14.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padilla-Camberos, E.; Arrizon, J.; Sandoval, G. Effect of Agave Fructan Bioconjugates on Metabolic Syndrome Parameters in a Murine Model. Pharmaceuticals 2023, 16, 412. https://doi.org/10.3390/ph16030412
Padilla-Camberos E, Arrizon J, Sandoval G. Effect of Agave Fructan Bioconjugates on Metabolic Syndrome Parameters in a Murine Model. Pharmaceuticals. 2023; 16(3):412. https://doi.org/10.3390/ph16030412
Chicago/Turabian StylePadilla-Camberos, Eduardo, Javier Arrizon, and Georgina Sandoval. 2023. "Effect of Agave Fructan Bioconjugates on Metabolic Syndrome Parameters in a Murine Model" Pharmaceuticals 16, no. 3: 412. https://doi.org/10.3390/ph16030412