
Exposure to Doxorubicin Modulates the Cardiac Response to Isoproterenol in Male and Female Mice

Abstract
:1. Introduction
2. Results
2.1. Baseline Cardiac Function
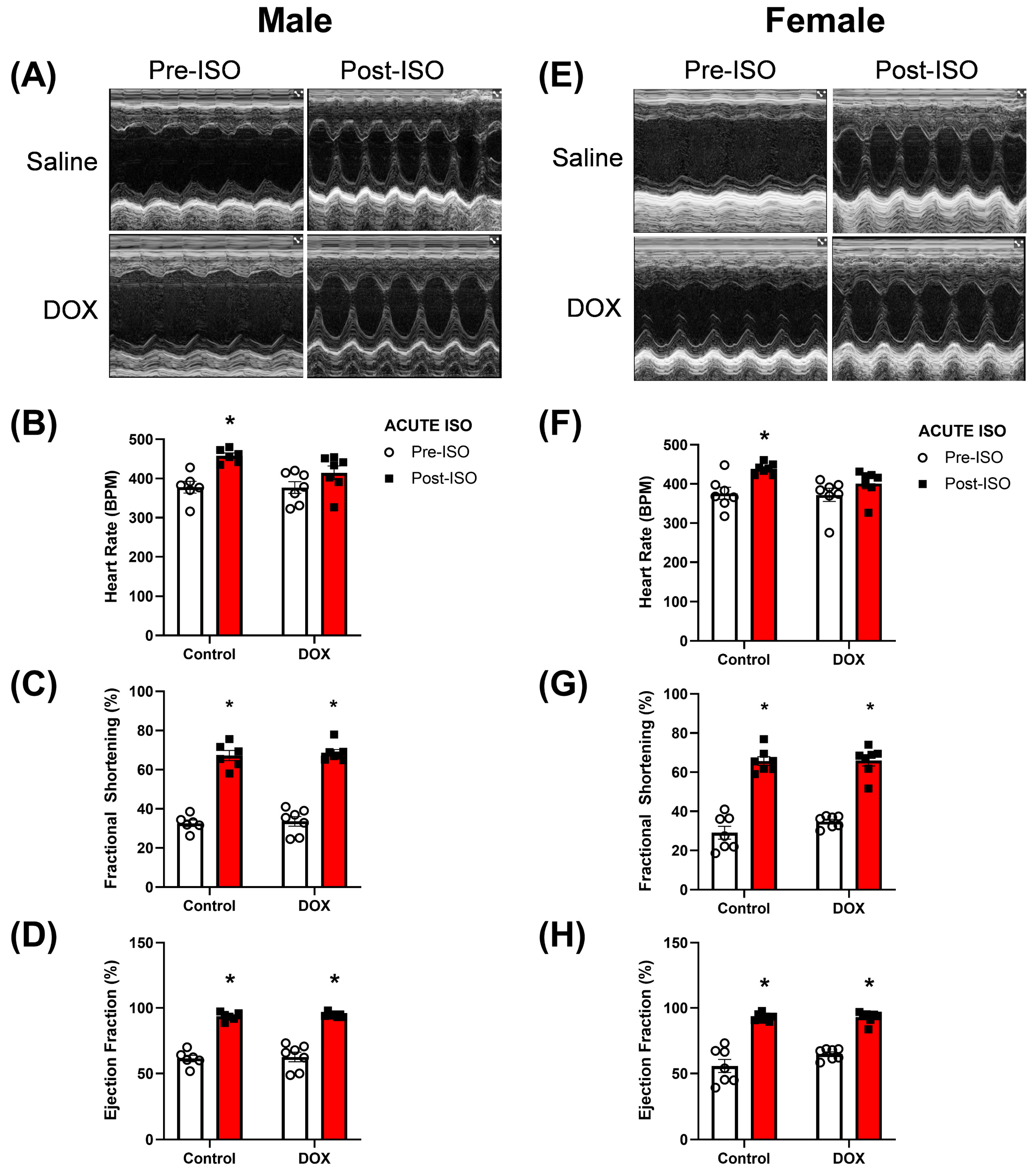
2.2. Acute ISO (Stress) Echocardiography
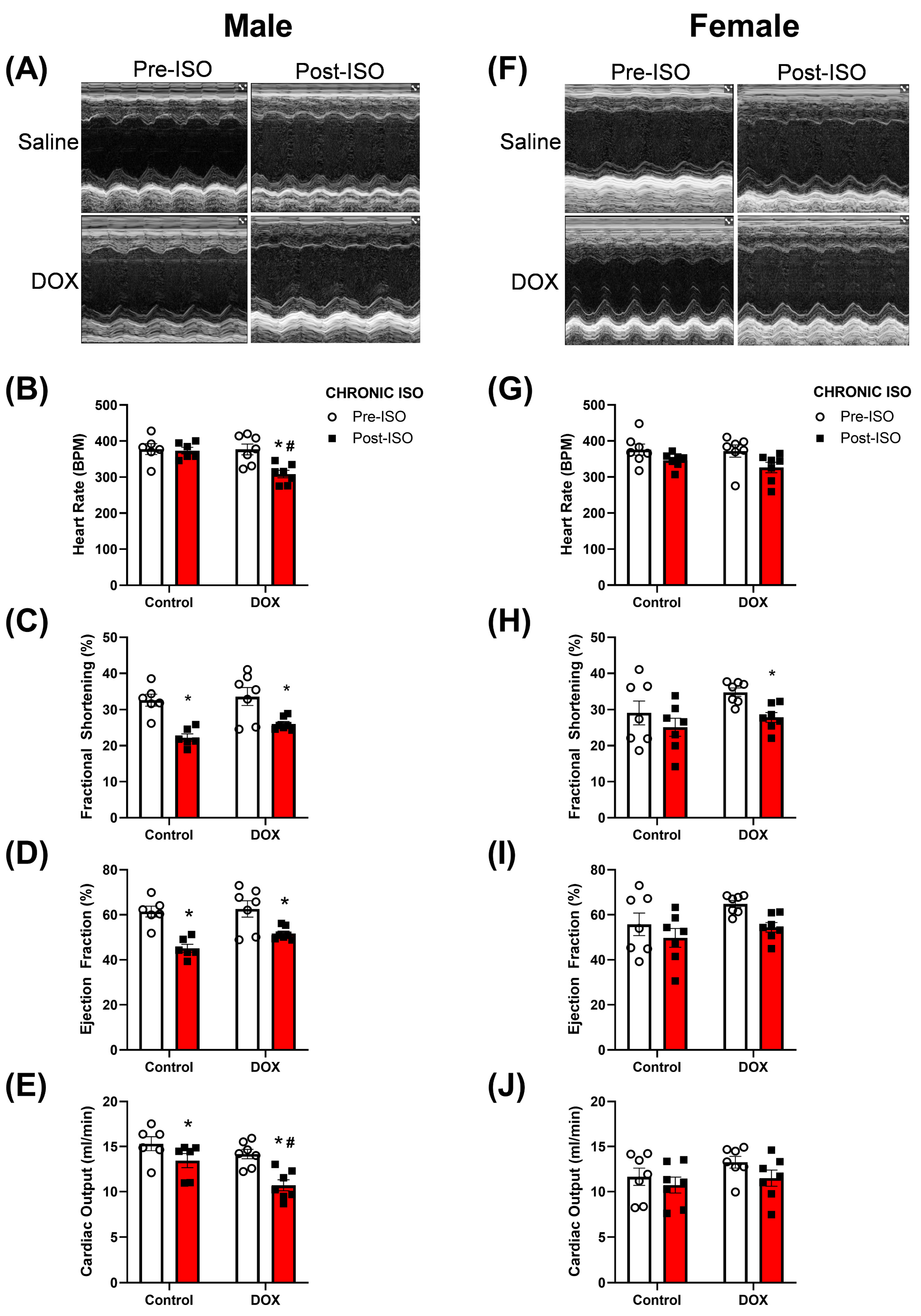
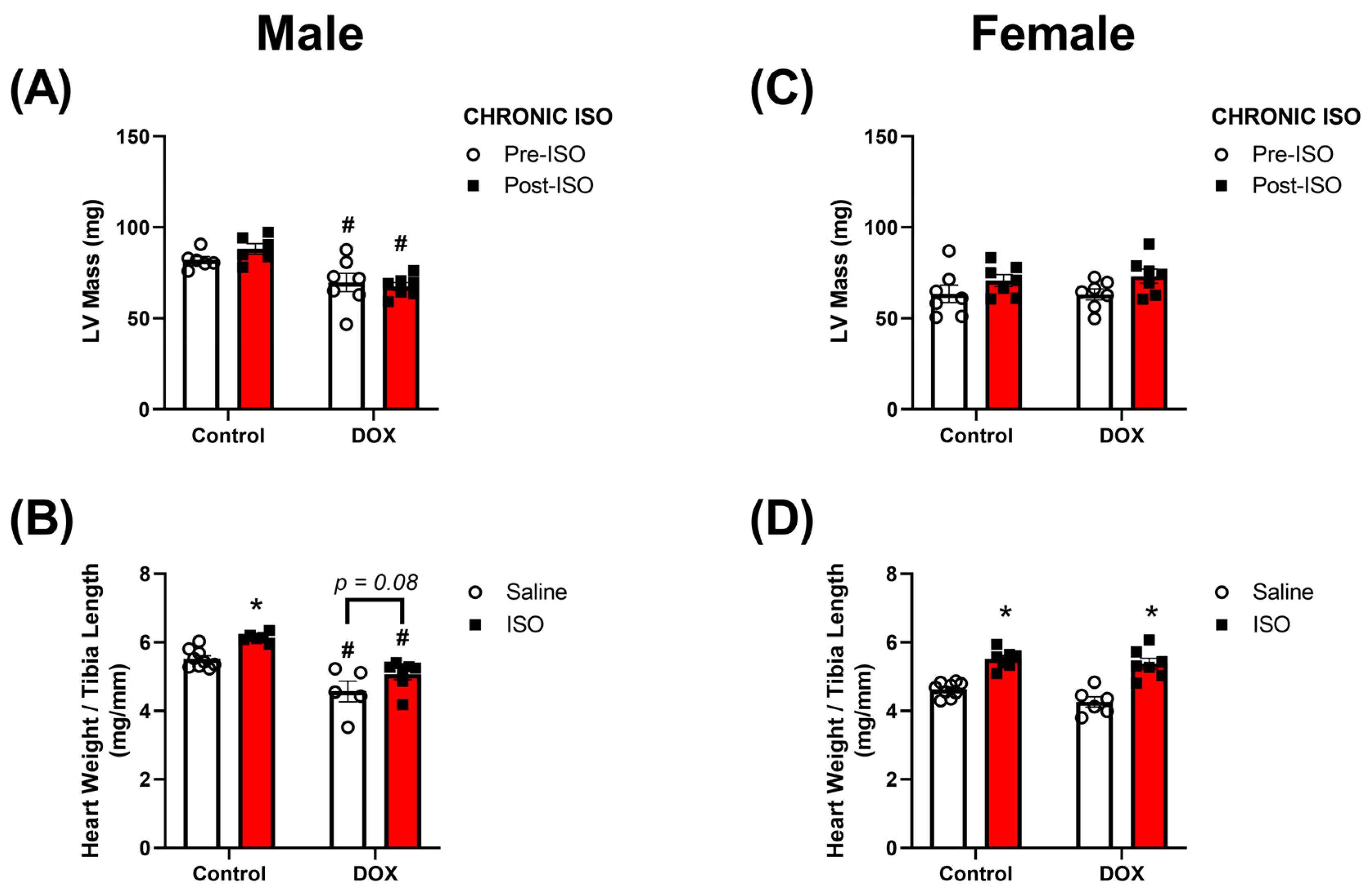
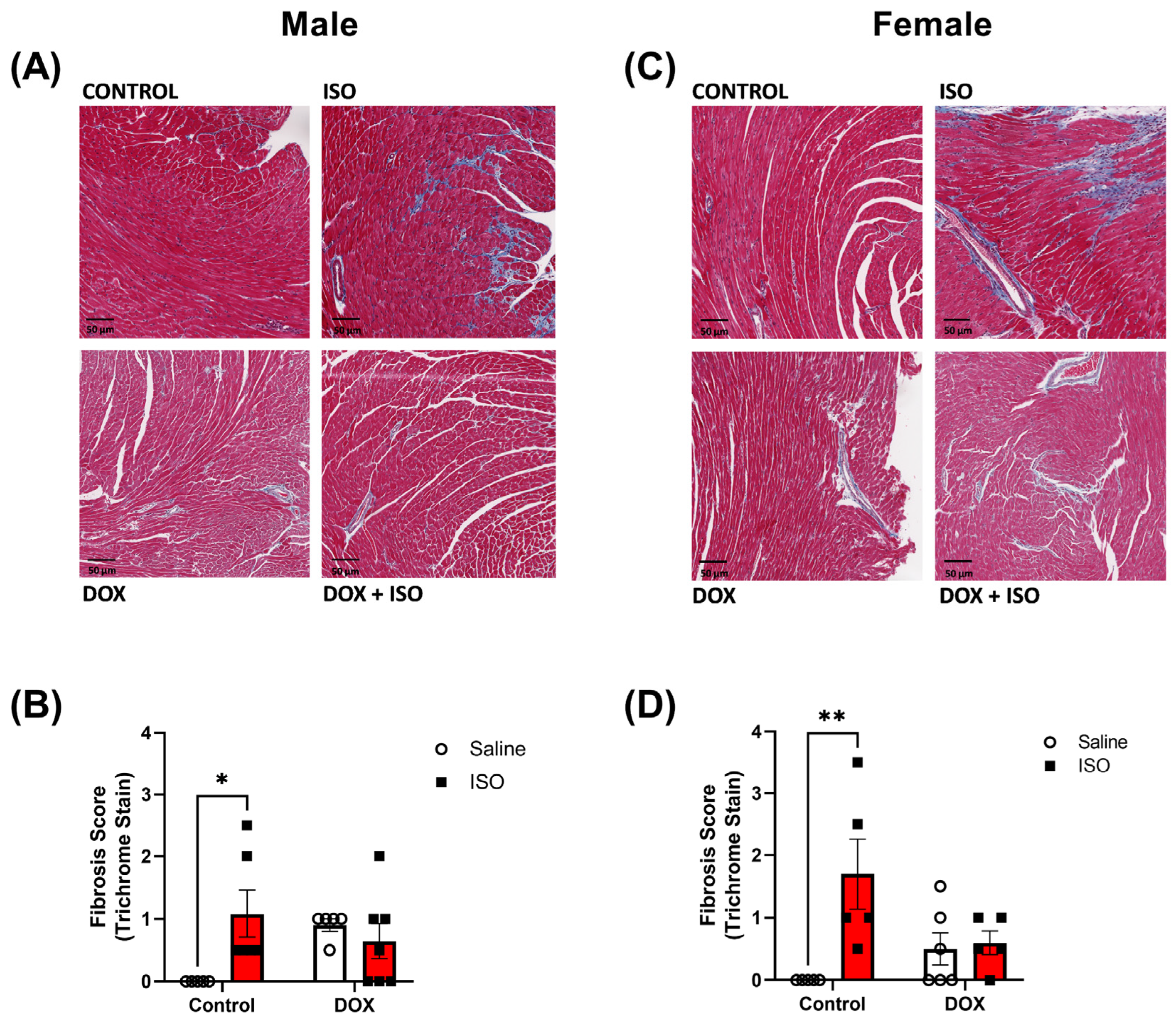
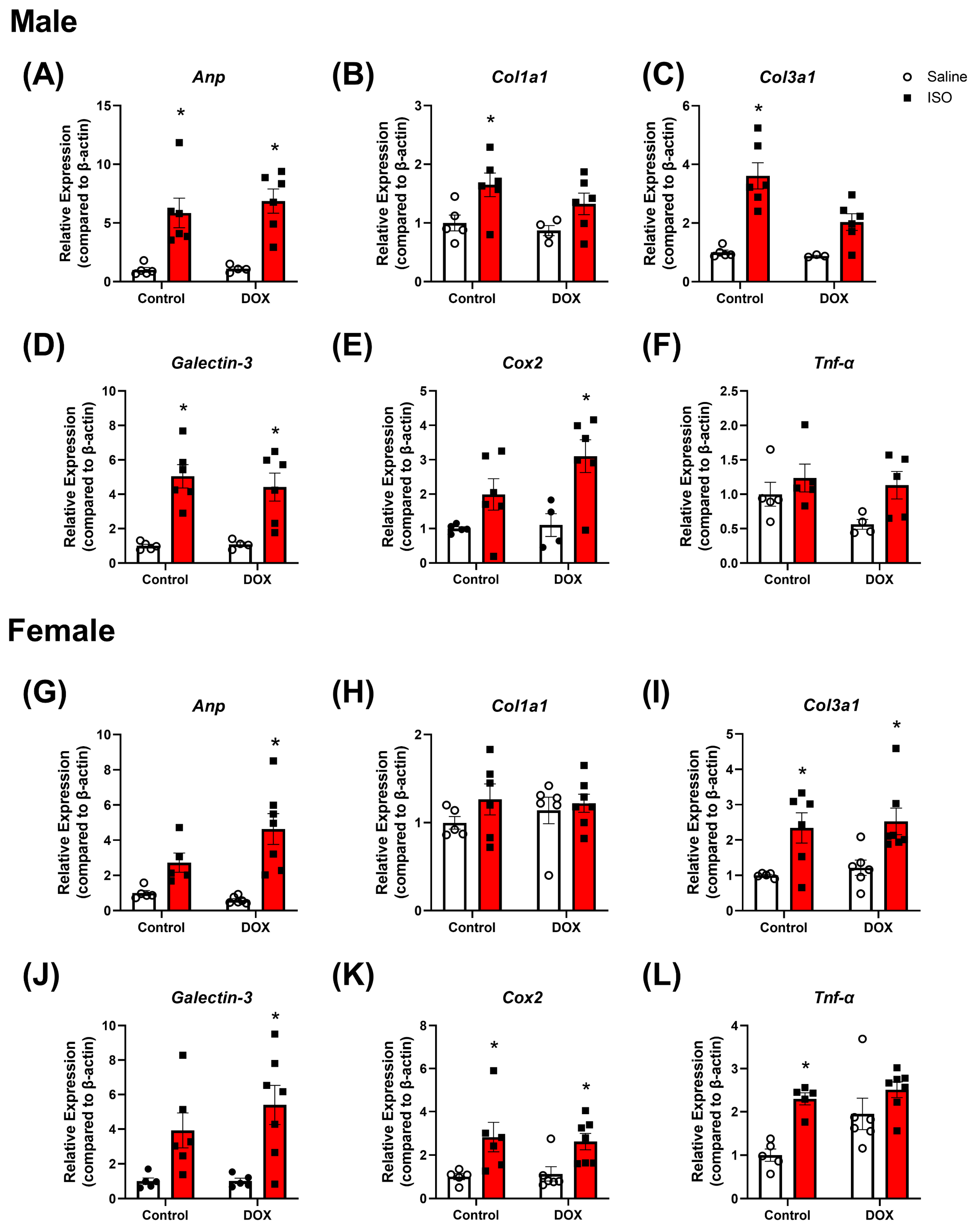
2.3. Response to Chronic ISO Administration in Intact Mice
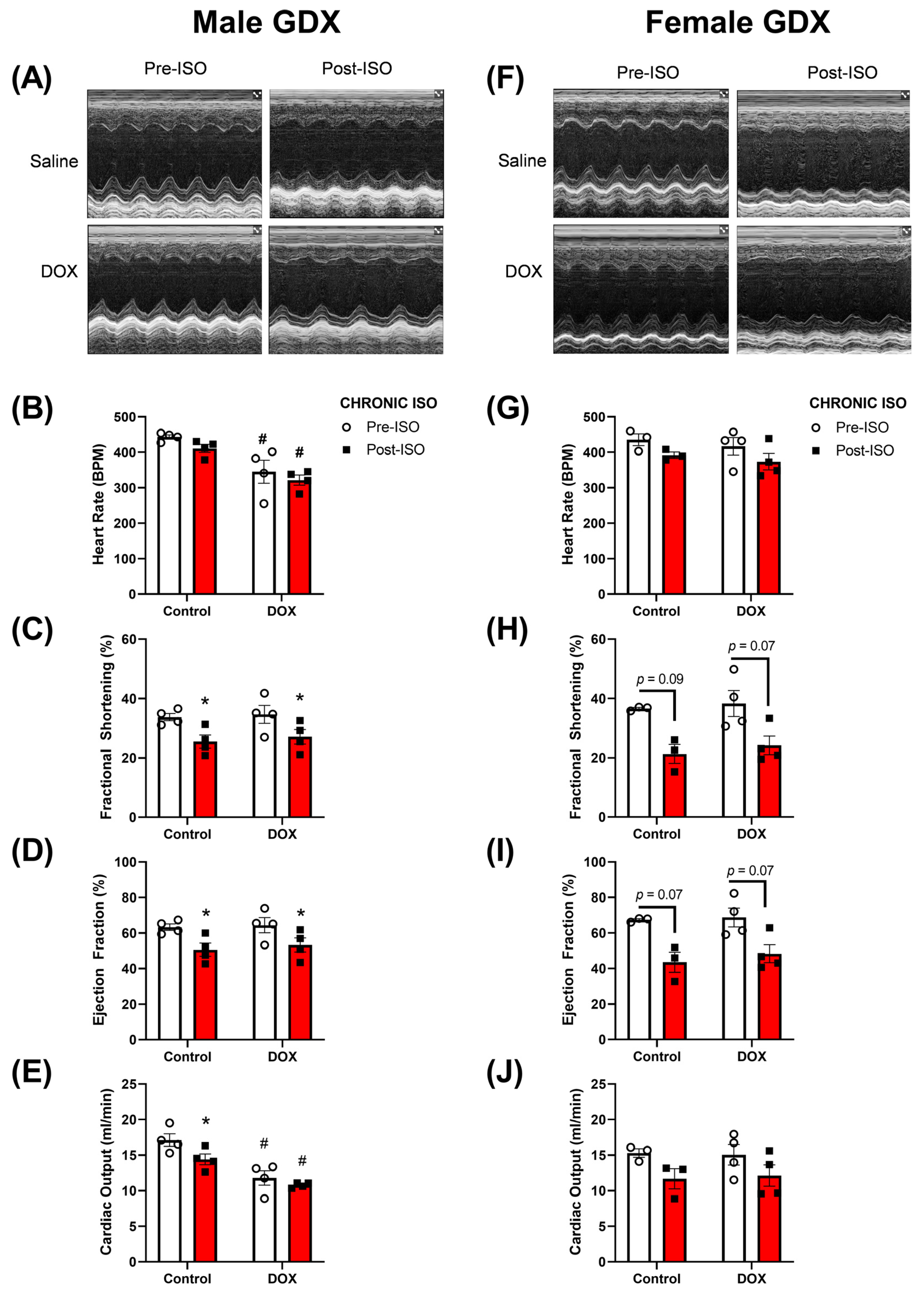
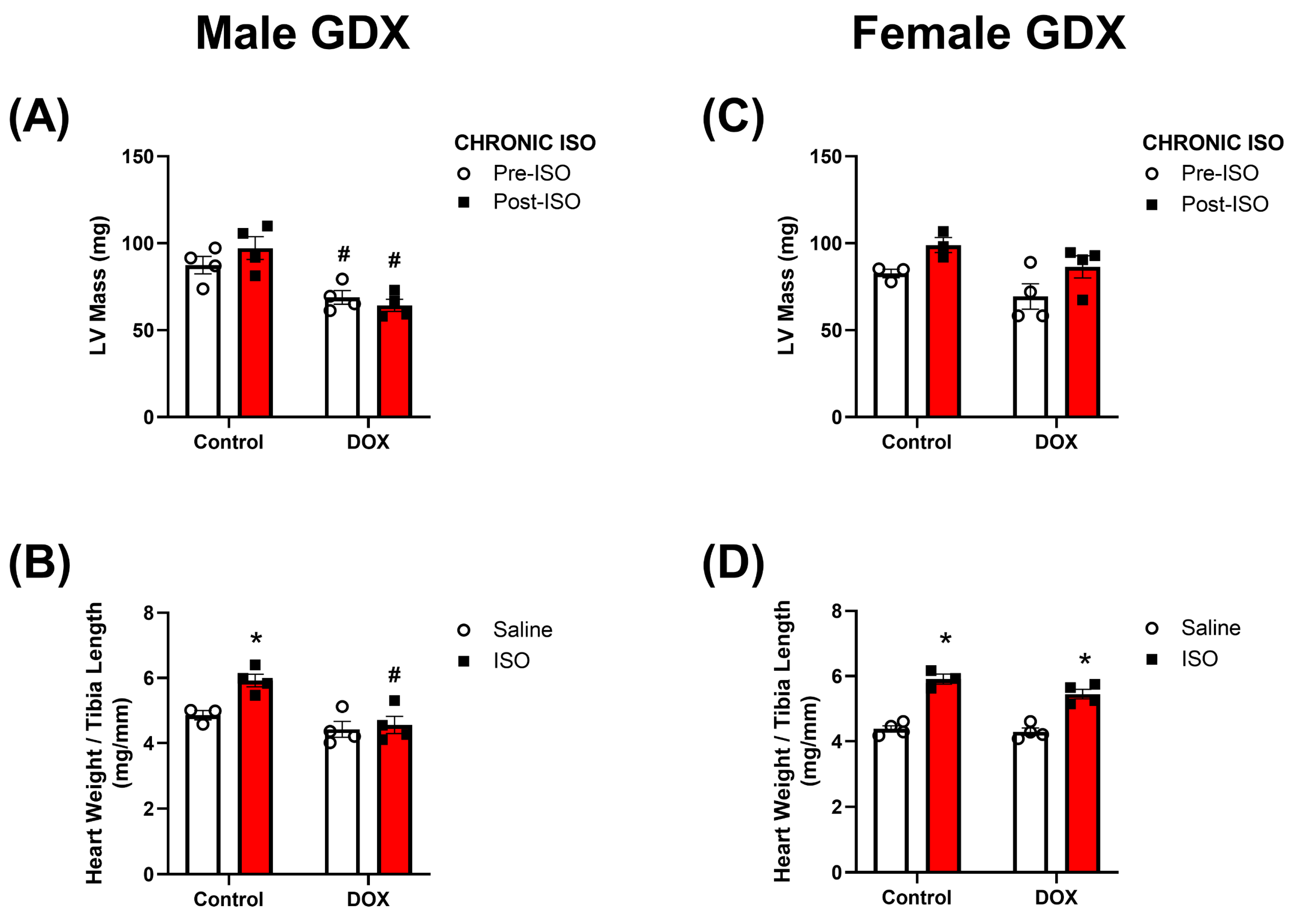
2.4. Response to Chronic ISO Administration in Gonadectomized Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Echocardiography
4.3. Histopathology
4.4. RNA Extraction and Real-Time PCR
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chatterjee, K.; Zhang, J.; Honbo, N.; Karliner, J.S. Doxorubicin cardiomyopathy. Cardiology 2010, 115, 155–162. [Google Scholar] [CrossRef]
- Lipshultz, S.E.; Diamond, M.B.; Franco, V.I.; Aggarwal, S.; Leger, K.; Santos, M.V.; Sallan, S.E.; Chow, E.J. Managing chemotherapy-related cardiotoxicity in survivors of childhood cancers. Paediatr. Drugs 2014, 16, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, S.E.; Sambatakos, P.; Maguire, M.; Karnik, R.; Ross, S.W.; Franco, V.I.; Miller, T.L. Cardiotoxicity and cardioprotection in childhood cancer. Acta Haematol. 2014, 132, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, S.E.; Cochran, T.R.; Franco, V.I.; Miller, T.L. Treatment-related cardiotoxicity in survivors of childhood cancer. Nat. Rev. Clin. Oncol. 2013, 10, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Desai, V.G.; Vijay, V.; Han, T.; Moland, C.L.; Phanavanh, B.; Lee, T.; Davis, K.J.; Muskhelishvili, L.; Stine, K.C.; Fuscoe, J.C. Doxorubicin-induced delayed-onset subclinical cardiotoxicity in mice. J. Appl. Toxicol. 2021, 42, 778–792. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.T.; Oeffinger, K.C.; Chen, Y.; Kawashima, T.; Yasui, Y.; Leisenring, W.; Stovall, M.; Chow, E.J.; Sklar, C.A.; Mulrooney, D.A.; et al. Modifiable risk factors and major cardiac events among adult survivors of childhood cancer. J. Clin. Oncol. 2013, 31, 3673–3680. [Google Scholar] [CrossRef]
- Matsumura, N.; Zordoky, B.N.; Robertson, I.M.; Hamza, S.M.; Parajuli, N.; Soltys, C.M.; Beker, D.L.; Grant, M.K.; Razzoli, M.; Bartolomucci, A.; et al. Co-administration of resveratrol with doxorubicin in young mice attenuates detrimental late-occurring cardiovascular changes. Cardiovasc. Res. 2018, 114, 1350–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maayah, Z.H.; Alam, A.S.; Takahara, S.; Soni, S.; Ferdaoussi, M.; Matsumura, N.; Zordoky, B.N.; Eisenstat, D.D.; Dyck, J.R.B. Resveratrol reduces cardiac NLRP3-inflammasome activation and systemic inflammation to lessen doxorubicin-induced cardiotoxicity in juvenile mice. FEBS Lett. 2021, 595, 1681–1695. [Google Scholar] [CrossRef]
- Agostinucci, K.; Grant, M.K.O.; Seelig, D.; Yücel, D.; van Berlo, J.; Bartolomucci, A.; Dyck, J.R.B.; Zordoky, B.N. Divergent Cardiac Effects of Angiotensin II and Isoproterenol Following Juvenile Exposure to Doxorubicin. Front. Cardiovasc. Med. 2022, 9, 742193. [Google Scholar] [CrossRef]
- Cottin, Y.; L’Huillier, I.; Casasnovas, O.; Geoffroy, C.; Caillot, D.; Zeller, M.; Solary, E.; Guy, H.; Wolf, J.E. Dobutamine stress echocardiography identifies anthracycline cardiotoxicity. Eur. J. Echocardiogr. 2000, 1, 180–183. [Google Scholar] [CrossRef] [Green Version]
- De Wolf, D.; Suys, B.; Maurus, R.; Benoit, Y.; Verhaaren, H.; Matthijs, D.; Otten, J. Dobutamine stress echocardiography in the evaluation of late anthracycline cardiotoxicity in childhood cancer survivors. Pediatr. Res. 1996, 39, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Hamada, H.; Ohkubo, T.; Maeda, M.; Ogawa, S. Evaluation of cardiac reserved function by high-dose dobutamine-stress echocardiography in asymptomatic anthracycline-treated survivors of childhood cancer. Pediatr. Int. 2006, 48, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Klewer, S.E.; Goldberg, S.J.; Donnerstein, R.L.; Berg, R.A.; Hutter, J.J., Jr. Dobutamine stress echocardiography: A sensitive indicator of diminished myocardial function in asymptomatic doxorubicin-treated long-term survivors of childhood cancer. J. Am. Coll. Cardiol. 1992, 19, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Khouri, M.G.; Hornsby, W.E.; Risum, N.; Velazquez, E.J.; Thomas, S.; Lane, A.; Scott, J.M.; Koelwyn, G.J.; Herndon, J.E.; Mackey, J.R.; et al. Utility of 3-dimensional echocardiography, global longitudinal strain, and exercise stress echocardiography to detect cardiac dysfunction in breast cancer patients treated with doxorubicin-containing adjuvant therapy. Breast Cancer Res. Treat. 2014, 143, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Meiners, B.; Shenoy, C.; Zordoky, B.N. Clinical and preclinical evidence of sex-related differences in anthracycline-induced cardiotoxicity. Biol. Sex Differ. 2018, 9, 38. [Google Scholar] [CrossRef]
- Myrehaug, S.; Pintilie, M.; Tsang, R.; Mackenzie, R.; Crump, M.; Chen, Z.; Sun, A.; Hodgson, D.C. Cardiac morbidity following modern treatment for Hodgkin lymphoma: Supra-additive cardiotoxicity of doxorubicin and radiation therapy. Leuk. Lymphoma 2008, 49, 1486–1493. [Google Scholar] [CrossRef]
- Zhang, X.J.; Cao, X.Q.; Zhang, C.S.; Zhao, Z. 17β-estradiol protects against doxorubicin-induced cardiotoxicity in male Sprague-Dawley rats by regulating NADPH oxidase and apoptosis genes. Mol. Med. Rep. 2017, 15, 2695–2702. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Knapton, A.; Lipshultz, S.E.; Cochran, T.R.; Hiraragi, H.; Herman, E.H. Sex-related differences in mast cell activity and doxorubicin toxicity: A study in spontaneously hypertensive rats. Toxicol. Pathol. 2014, 42, 361–375. [Google Scholar] [CrossRef]
- Grant, M.K.; Seelig, D.M.; Sharkey, L.C.; Zordoky, B.N. Sex-dependent alteration of cardiac cytochrome P450 gene expression by doxorubicin in C57Bl/6 mice. Biol. Sex Differ. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zordoky, B.N.; Radin, M.J.; Heller, L.; Tobias, A.; Matise, I.; Apple, F.S.; McCune, S.A.; Sharkey, L.C. The interplay between genetic background and sexual dimorphism of doxorubicin-induced cardiotoxicity. Cardiooncology 2016, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipshultz, S.E.; Colan, S.D.; Gelber, R.D.; Perez-Atayde, A.R.; Sallan, S.E.; Sanders, S.P. Late cardiac effects of doxorubicin therapy for acute lymphoblastic leukemia in childhood. N. Engl. J. Med. 1991, 324, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Leger, K.; Slone, T.; Lemler, M.; Leonard, D.; Cochran, C.; Bowman, W.P.; Bashore, L.; Winick, N. Subclinical cardiotoxicity in childhood cancer survivors exposed to very low dose anthracycline therapy. Pediatr. Blood Cancer 2015, 62, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Vandecruys, E.; Mondelaers, V.; De Wolf, D.; Benoit, Y.; Suys, B. Late cardiotoxicity after low dose of anthracycline therapy for acute lymphoblastic leukemia in childhood. J. Cancer Surviv. 2012, 6, 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira-Rodrigues, M.; Graça, A.L.; Ferreira, M.; Afonso, J.; Serrão, P.; Morato, M.; Ferreirinha, F.; Correia-de-Sá, P.; Ebert, S.N.; Moura, D. Attenuated aortic vasodilation and sympathetic prejunctional facilitation in epinephrine-deficient mice: Selective impairment of β2-adrenoceptor responses. J. Pharmacol. Exp. Ther. 2014, 351, 243–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leblais, V.; Delannoy, E.; Fresquet, F.; Bégueret, H.; Bellance, N.; Banquet, S.; Allières, C.; Leroux, L.; Desgranges, C.; Gadeau, A.; et al. beta-adrenergic relaxation in pulmonary arteries: Preservation of the endothelial nitric oxide-dependent beta2 component in pulmonary hypertension. Cardiovasc. Res. 2008, 77, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.; Della Torre, P.; Allievi, C.; Morisetti, A.; Al-Fayoumi, S.; Singer, J.W. Tolerability and toxicological profile of pixantrone (Pixuvri®) in juvenile mice. Comparative study with doxorubicin. Reprod. Toxicol. 2014, 46, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Moulin, M.; Piquereau, J.; Mateo, P.; Fortin, D.; Rucker-Martin, C.; Gressette, M.; Lefebvre, F.; Gresikova, M.; Solgadi, A.; Veksler, V.; et al. Sexual dimorphism of doxorubicin-mediated cardiotoxicity: Potential role of energy metabolism remodeling. Circ. Heart Fail. 2015, 8, 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, G.R.; Lee, T.; Moland, C.L.; Vijay, V.; Herman, E.H.; Lewis, S.M.; Davis, K.J.; Muskhelishvili, L.; Kerr, S.; Fuscoe, J.C.; et al. Sex-related differential susceptibility to doxorubicin-induced cardiotoxicity in B6C3F1 mice. Toxicol. Appl. Pharmacol. 2016, 310, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.K.O.; Abdelgawad, I.Y.; Lewis, C.A.; Seelig, D.; Zordoky, B.N. Lack of sexual dimorphism in a mouse model of isoproterenol-induced cardiac dysfunction. PLoS ONE 2020, 15, e0232507. [Google Scholar] [CrossRef]
- Zhu, W.; Soonpaa, M.H.; Chen, H.; Shen, W.; Payne, R.M.; Liechty, E.A.; Caldwell, R.L.; Shou, W.; Field, L.J. Acute doxorubicin cardiotoxicity is associated with p53-induced inhibition of the mammalian target of rapamycin pathway. Circulation 2009, 119, 99–106. [Google Scholar] [CrossRef]
- Xia, P.; Chen, J.; Liu, Y.; Fletcher, M.; Jensen, B.C.; Cheng, Z. Doxorubicin induces cardiomyocyte apoptosis and atrophy through cyclin-dependent kinase 2-mediated activation of forkhead box O1. J. Biol. Chem. 2020, 295, 4265–4276. [Google Scholar] [CrossRef]
- Antoniak, S.; Phungphong, S.; Cheng, Z.; Jensen, B.C. Novel Mechanisms of Anthracycline-Induced Cardiovascular Toxicity: A Focus on Thrombosis, Cardiac Atrophy, and Programmed Cell Death. Front. Cardiovasc. Med. 2021, 8, 817977. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Yan, J.; Yang, P.Z. Cardiomyocyte Atrophy, an Underestimated Contributor in Doxorubicin-Induced Cardiotoxicity. Front. Cardiovasc. Med. 2022, 9, 812578. [Google Scholar] [CrossRef]
- Cosper, P.F.; Leinwand, L.A. Cancer causes cardiac atrophy and autophagy in a sexually dimorphic manner. Cancer Res. 2011, 71, 1710–1720. [Google Scholar] [CrossRef] [Green Version]
- Rosa-Caldwell, M.E.; Greene, N.P. Muscle metabolism and atrophy: Let’s talk about sex. Biol. Sex Differ. 2019, 10, 43. [Google Scholar] [CrossRef]
- Abdelgawad, I.Y.; Sadak, K.T.; Lone, D.W.; Dabour, M.S.; Niedernhofer, L.J.; Zordoky, B.N. Molecular mechanisms and cardiovascular implications of cancer therapy-induced senescence. Pharmacol. Ther. 2021, 221, 107751. [Google Scholar] [CrossRef] [PubMed]
- Espitia-Corredor, J.A.; Shamoon, L.; Olivares-Silva, F.; Rimassa-Tare, C.; Munoz-Rodriguez, C.; Espinoza-Perez, C.; Sanchez-Ferrer, C.F.; Peiro, C.; Diaz-Araya, G. Resolvin E1 attenuates doxorubicin-induced cardiac fibroblast senescence: A key role for IL-1beta. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166525. [Google Scholar] [CrossRef] [PubMed]
- Mancilla, T.R.; Davis, L.R.; Aune, G.J. Doxorubicin-induced p53 interferes with mitophagy in cardiac fibroblasts. PLoS ONE 2020, 15, e0238856. [Google Scholar] [CrossRef] [PubMed]
- Yusifov, A.; Chhatre, V.E.; Zumo, J.M.; Cook, R.F.; McNair, B.D.; Schmitt, E.E.; Woulfe, K.C.; Bruns, D.R. Cardiac response to adrenergic stress differs by sex and across the lifespan. Geroscience 2021, 43, 1799–1813. [Google Scholar] [CrossRef]
- Bujak, M.; Kweon, H.J.; Chatila, K.; Li, N.; Taffet, G.; Frangogiannis, N.G. Aging-related defects are associated with adverse cardiac remodeling in a mouse model of reperfused myocardial infarction. J. Am. Coll. Cardiol. 2008, 51, 1384–1392. [Google Scholar] [CrossRef] [Green Version]
- Phungphong, S.; Kijtawornrat, A.; Kampaengsri, T.; Wattanapermpool, J.; Bupha-Intr, T. Comparison of exercise training and estrogen supplementation on mast cell-mediated doxorubicin-induced cardiotoxicity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R829–R842. [Google Scholar] [CrossRef]
- Calvé, A.; Haddad, R.; Barama, S.N.; Meilleur, M.; Sebag, I.A.; Chalifour, L.E. Cardiac response to doxorubicin and dexrazoxane in intact and ovariectomized young female rats at rest and after swim training. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H2048–H2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Fu, L.; Tang, X.; Han, Y.; Ma, D.; Cao, J.; Kang, N.; Ji, H. Testosterone modulation of cardiac β-adrenergic signals in a rat model of heart failure. Gen. Comp. Endocrinol. 2011, 172, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, Y.; Zeng, Z.; Li, Y.; Su, H.; Li, Y.; Wang, R.; Zhang, M.; Yang, Y.; Deng, J. Influence of androgen on myocardial apoptosis and expression of myocardial IR and IRS1 in chronic heart failure rat models. Mol. Med. Rep. 2018, 17, 1057–1064. [Google Scholar] [CrossRef] [Green Version]
- Peter, A.K.; Walker, C.J.; Ceccato, T.; Trexler, C.L.; Ozeroff, C.D.; Lugo, K.R.; Perry, A.R.; Anseth, K.S.; Leinwand, L.A. Cardiac Fibroblasts Mediate a Sexually Dimorphic Fibrotic Response to beta-Adrenergic Stimulation. J. Am. Heart Assoc. 2021, 10, e018876. [Google Scholar] [CrossRef]
- Milasin, J.; Muntoni, F.; Severini, G.M.; Bartoloni, L.; Vatta, M.; Krajinovic, M.; Mateddu, A.; Angelini, C.; Camerini, F.; Falaschi, A.; et al. A point mutation in the 5’ splice site of the dystrophin gene first intron responsible for X-linked dilated cardiomyopathy. Hum. Mol. Genet. 1996, 5, 73–79. [Google Scholar] [CrossRef]
- Snell, D.M.; Turner, J.M.A. Sex Chromosome Effects on Male-Female Differences in Mammals. Curr. Biol. 2018, 28, R1313–R1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, A.P. Y chromosome’s roles in sex differences in disease. Proc. Natl. Acad. Sci. USA 2017, 114, 3787–3789. [Google Scholar] [CrossRef] [Green Version]
- Le-Ha, C.; Beilin, L.J.; Burrows, S.; Keelan, J.A.; Hickey, M.; Mori, T.A. Prenatal Testosterone Associates with Blood Pressure in Young Adults: A Prospective Cohort Study. Hypertension 2021, 77, 1756–1764. [Google Scholar] [CrossRef]
- Shi, W.; Sheng, X.; Dorr, K.M.; Hutton, J.E.; Emerson, J.I.; Davies, H.A.; Andrade, T.D.; Wasson, L.K.; Greco, T.M.; Hashimoto, Y.; et al. Cardiac proteomics reveals sex chromosome-dependent differences between males and females that arise prior to gonad formation. Dev. Cell. 2021, 56, 3019–3034.e7. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 1-Week Post DOX | 5-Week Post DOX | ||||||
|---|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | |||||
| Saline | DOX | Saline | DOX | Saline | DOX | Saline | DOX | |
| CO (mL/min) | 15.56 ± 0.55 | 13.07 ± 0.64 | 13.21 ± 1.06 | 13.17 ± 0.89 | 15.31 ± 0.78 | 14.16 ± 0.52 | 11.65 ± 0.96 | 13.26 ± 0.65 |
| HR (BPM) | 363 ± 8 | 340 ± 14 | 366 ± 14 | 362 ± 13 | 378 ± 15 | 377 ± 15 | 376 ± 15 | 372 ± 17 |
| SV (µL) | 42.85 ± 1.41 | 38.48 ± 0.79 | 35.75 ± 1.84 | 36.13 ± 1.25 | 40.65 ± 1.84 | 37.72 ± 1.16 | 30.89 ± 2.13 | 35.68 ± 0.84 |
| FS (%) | 30.10 ± 1.82 | 31.73 ± 2.36 | 29.86 ± 1.56 | 32.15 ± 32.15 | 32.64 ± 1.64 | 33.59 ± 2.46 | 29.09 ± 3.30 | 34.81 ± 1.13 |
| EF (%) | 57.48 ± 2.82 | 59.90 ± 3.66 | 57.39 ± 2.36 | 60.93 ± 1.63 | 61.42 ± 2.41 | 62.59 ± 3.63 | 55.75 ± 4.99 | 64.80 ± 1.57 |
| LVESV (µL) | 31.46 ± 4.22 | 26.59 ± 3.93 | 27.34 ± 2.63 | 23.75 ± 1.67 | 25.48 ± 4.17 | 24.65 ± 4.75 | 26.09 ± 4.58 | 19.43 ± 1.28 |
| LVEDV (µL) | 73.00 ± 3.72 | 63.44 ± 3.43 | 61.17 ± 3.00 | 59.37 ± 2.69 | 64.50 ± 5.55 | 60.87 ± 6.91 | 53.58 ± 5.06 | 54.45 ± 1.43 |
| LV Mass (mg) | 80.06 ± 4.03 | 66.85 ± 3.63 * | 66.69 ± 3.84 | 65.96 ± 2.81 | 82.02 ± 1.97 | 69.75 ± 5.01 | 63.49 ± 4.82 | 63.17 ± 2.97 |
| LVAW;s (mm) | 1.11 ± 0.092 | 1.04 ± 0.075 | 0.94 ± 0.061 | 0.90 ± 0.055 | 1.33 ± 0.076 | 1.11 ± 0.29 * | 0.92 ± 0.073 | 1.00 ± 0.044 |
| LVAW;d (mm) | 0.75 ± 0.037 | 0.71 ± 0.060 | 0.595 ± 0.040 | 0.580 ± 0.043 | 0.85 ± 0.036 | 0.70 ± 0.025 * | 0.68 ± 0.046 | 0.63 ± 0.037 |
| LVPW;s (mm) | 1.01 ± 0.057 | 0.96 ± 0.051 | 0.93 ± 0.057 | 0.99 ± 0.034 | 1.09 ± 0.046 | 1.00 ± 0.019 | 0.90 ± 0.053 | 1.02 ± 0.045 |
| LVPW;d (mm) | 0.64 ± 0.017 | 0.59 ± 0.032 | 0.69 ± 0.041 | 0.67 ± 0.021 | 0.66 ± 0.023 | 0.59 ± 0.048 | 0.57 ± 0.023 | 0.64 ± 0.026 |
| Gene ID | Forward Primer | Reverse Primer |
|---|---|---|
| Anp | GGAGCCTACGAAGATCCAGC | TCCAATCCTGTCAATCCTACCC |
| Col1a1 | CTGGCGGTTCAGGTCCAAT | TTCCAGGCAATCCACGAGC |
| Col3a1 | ATGGTGGTTTTCAGTTCAGCTATG | GCCCGGCTGGAAAGAAGT |
| Galectin-3 | TATCCTGCTGCTGGCCCTTAT | GTTTGCGTTGGGTTTCACTG |
| Cox2 | CTGGTGCCTGGTCTGATGATG | GGCAATGCGGTTCTGATACTG |
| Tnf-ɑ | CCAGACCCTCACACTCAGATCA | CACTTGGTGGTTTGCTACGAC |
| β-actin | TATTGGCAACGAGCGGTTCC | GGCATAGAGGTCTTTACGGATGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agostinucci, K.; Grant, M.K.O.; Melaku, W.; Nair, C.; Zordoky, B.N. Exposure to Doxorubicin Modulates the Cardiac Response to Isoproterenol in Male and Female Mice. Pharmaceuticals 2023, 16, 391. https://doi.org/10.3390/ph16030391
Agostinucci K, Grant MKO, Melaku W, Nair C, Zordoky BN. Exposure to Doxorubicin Modulates the Cardiac Response to Isoproterenol in Male and Female Mice. Pharmaceuticals. 2023; 16(3):391. https://doi.org/10.3390/ph16030391
Chicago/Turabian StyleAgostinucci, Kevin, Marianne K. O. Grant, Wongel Melaku, Chandini Nair, and Beshay N. Zordoky. 2023. "Exposure to Doxorubicin Modulates the Cardiac Response to Isoproterenol in Male and Female Mice" Pharmaceuticals 16, no. 3: 391. https://doi.org/10.3390/ph16030391