
Antigen Unmasking Is Required to Clinically Assess Levels and Localisation Patterns of Phospholipase C Zeta in Human Sperm
 , , and
, , and 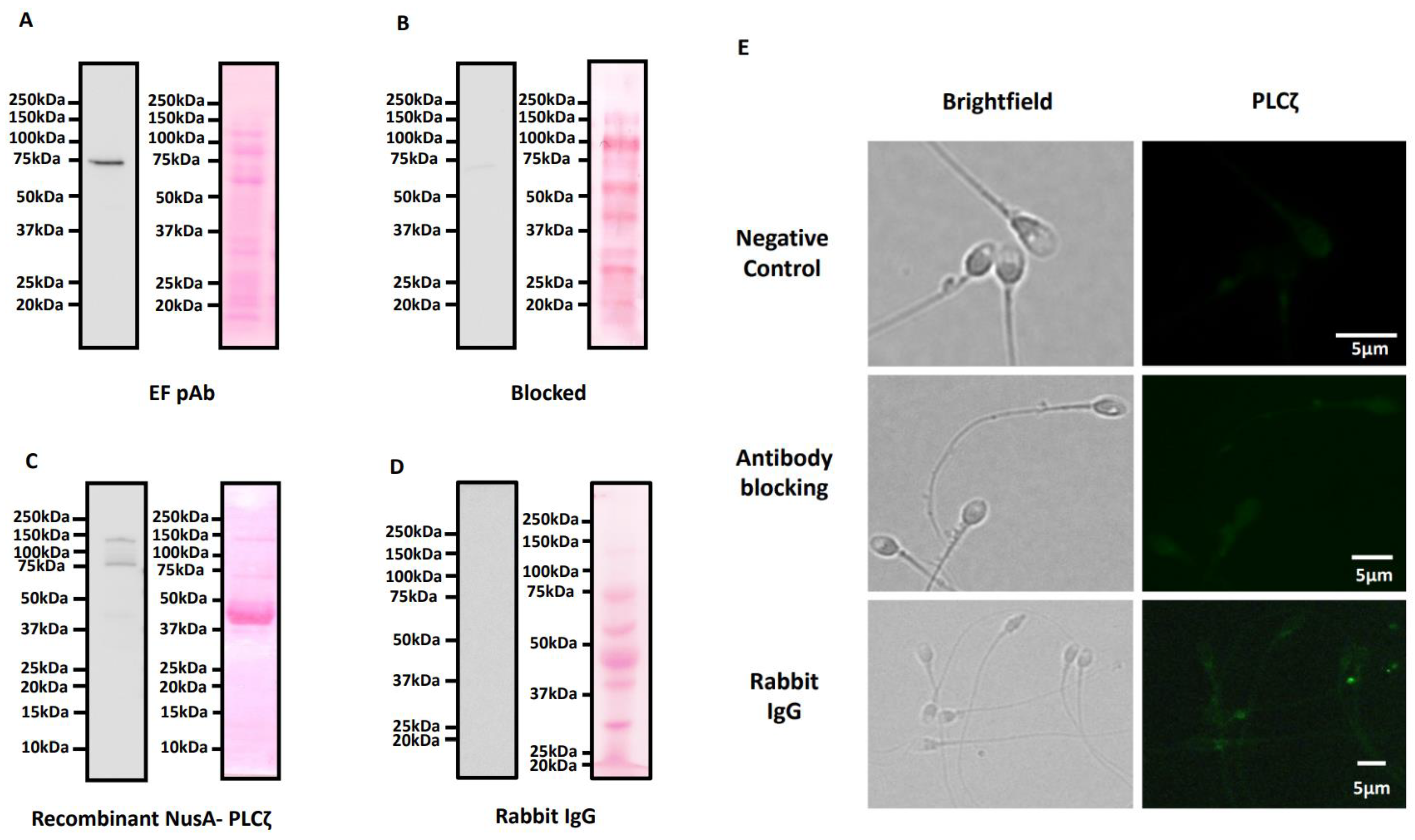{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
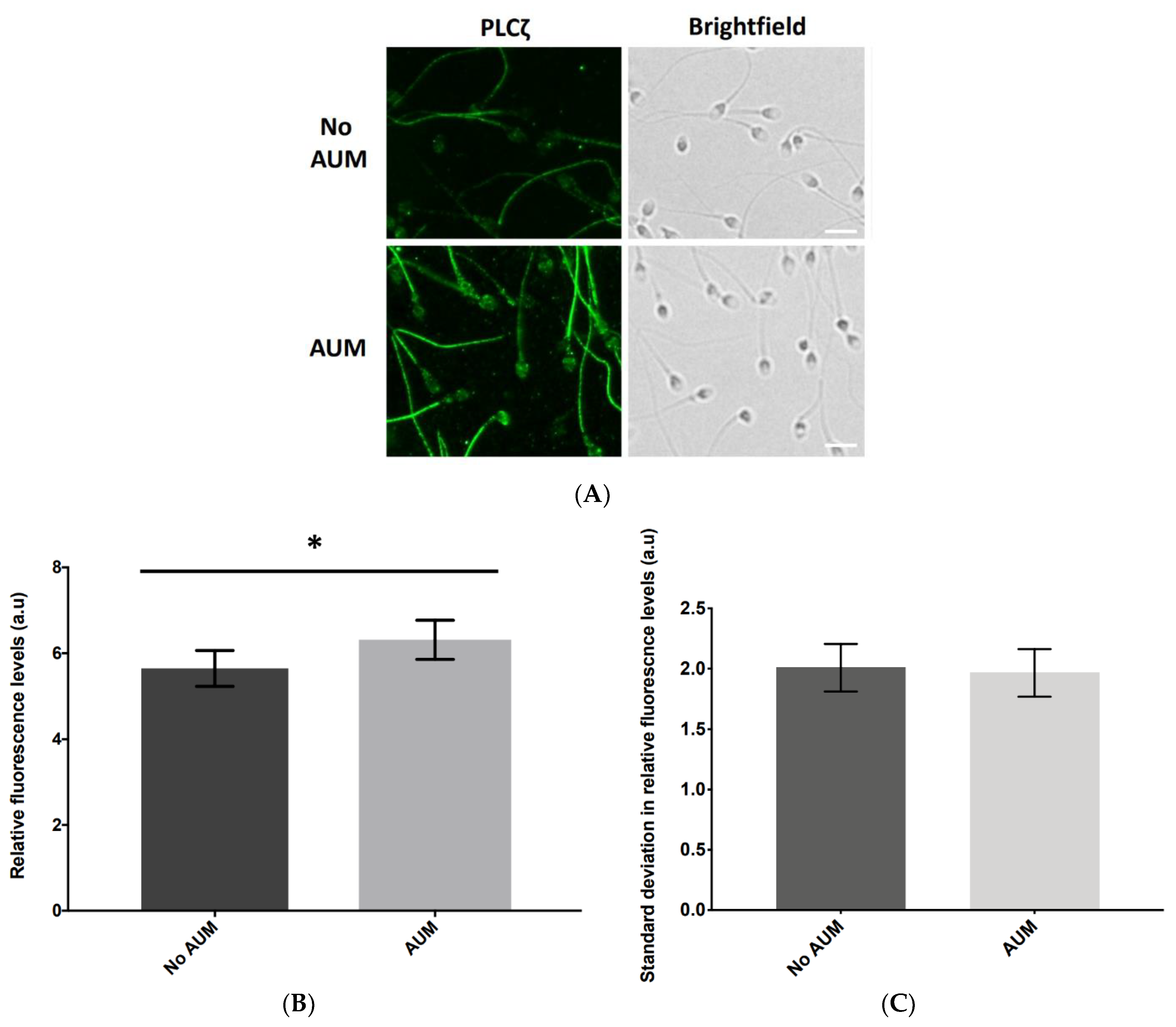
2.1. Antigen Unmasking (AUM) Differentially Enhances the Levels of Observable PLCζ in Sperm from Individual Human Patients
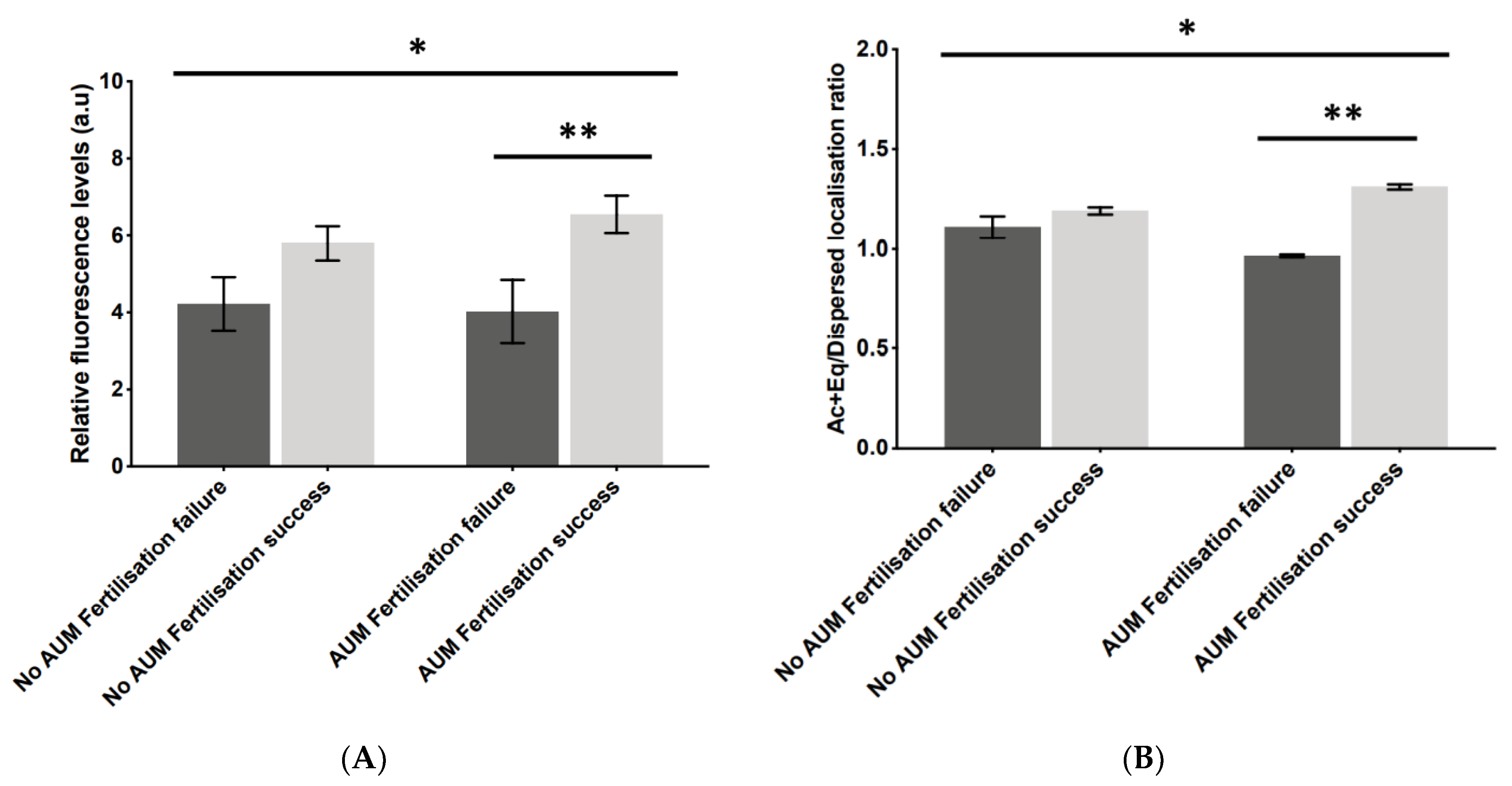
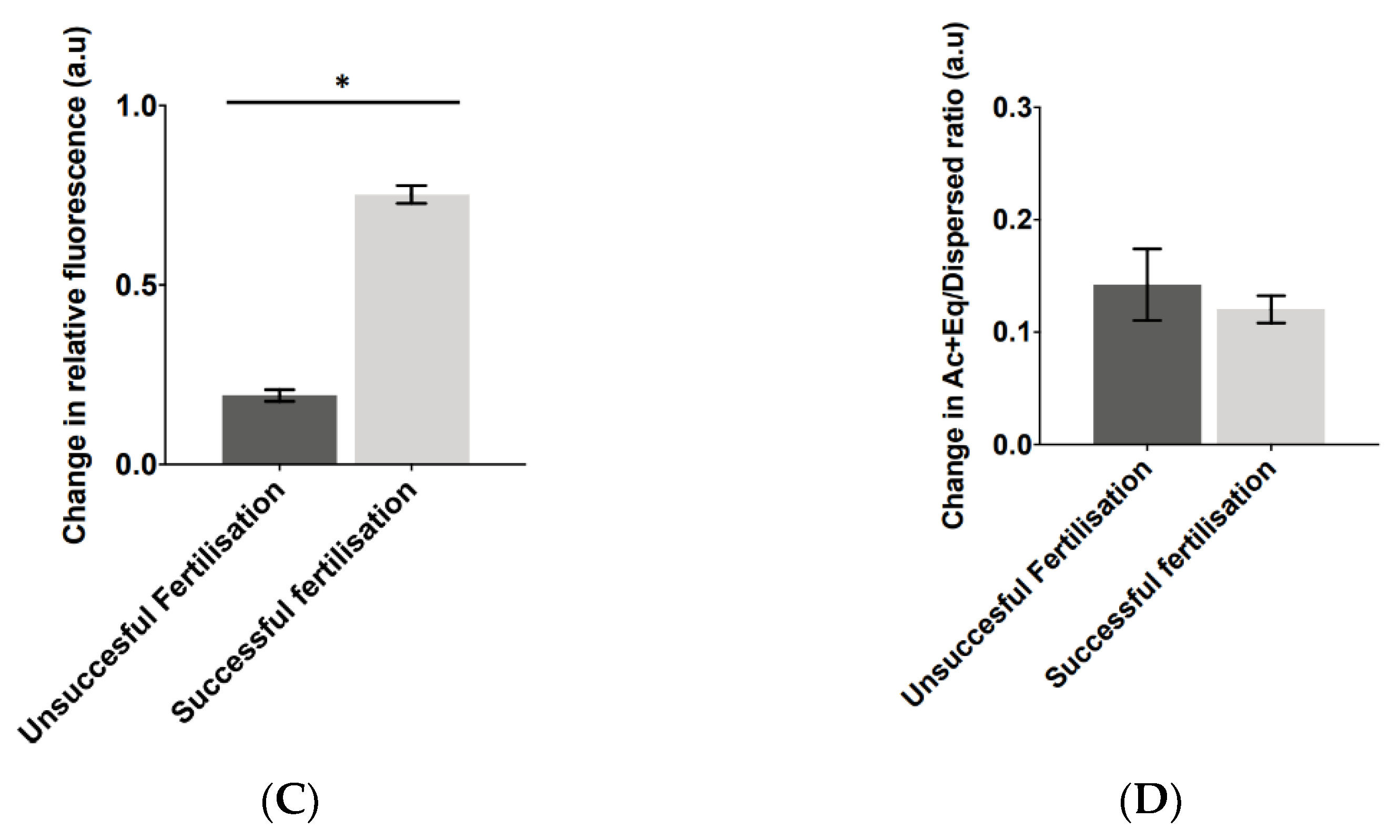
2.2. Application of AUM Is Required to Use PLCζ Fluorescence Levels and Localisation Patterns as Indicators of Fertilisation Outcome
2.3. AUM Treatment Did Not Affect Observed Proportions of PLCζ Localisation among Sperm Parameters Examined
2.4. Ac + Eq/Dispersed PLCζ Localisation Pattern Ratios Altered Significantly following AUM
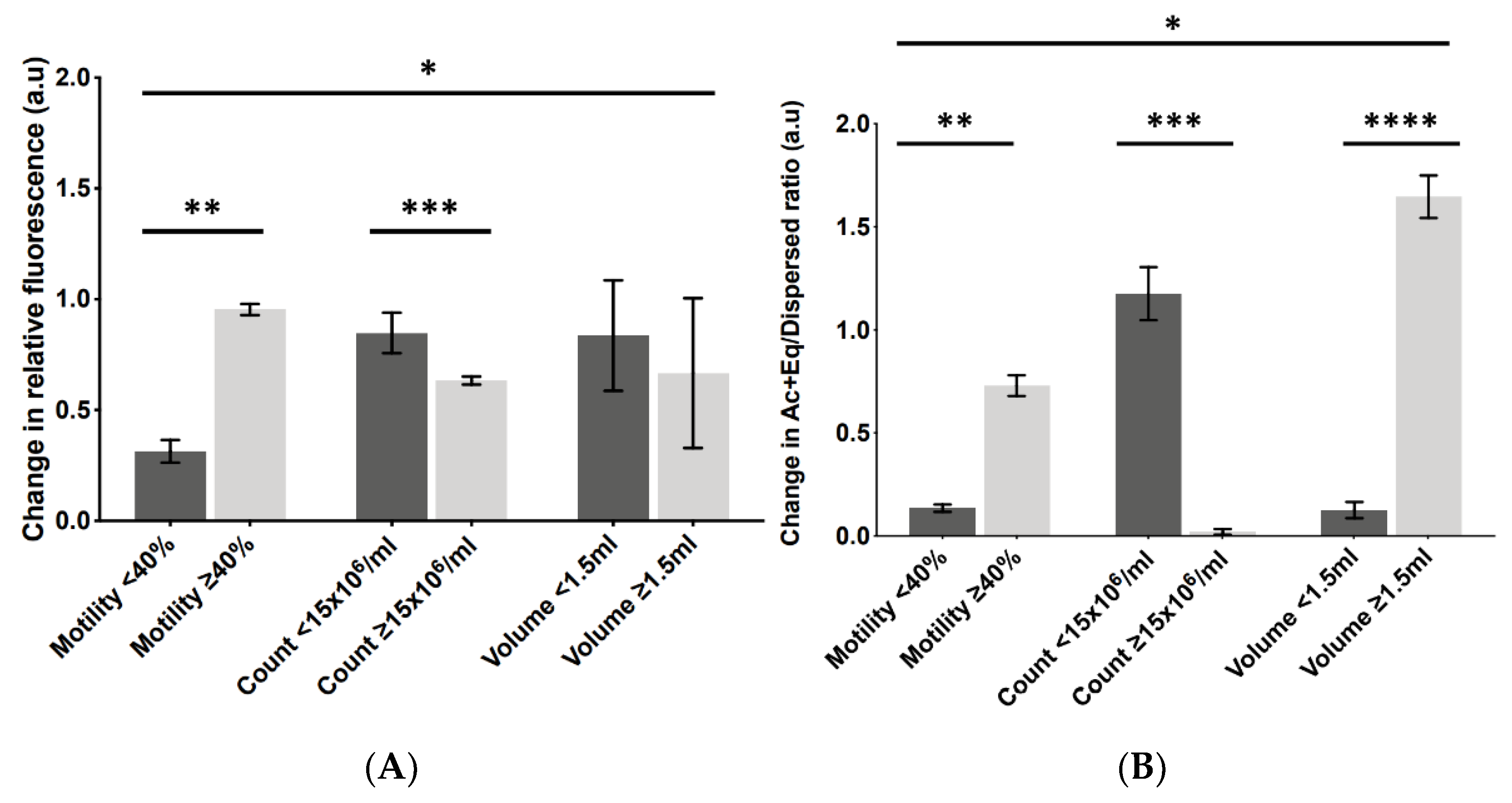
2.5. Levels of Change in PLCζ Parameters following AUM Corresponded to Optimal Sperm Parameters
3. Discussion
3.1. AUM Is Required to Reveal Significantly Higher PLCζ Parameters in Cases of Successful Fertilisation
3.2. AUM Altered the Ac + Eq/Dispersed PLCζ Ratio in Relation to Optimal Sperm Parameters
3.3. The Level of Change in PLCζ Parameters following AUM Is Related to Optimal Sperm Parameters and Fertilisation Success
4. Conclusions
5. Materials and Methods
5.1. Patient Recruitment and Semen/Sperm Processing
5.2. Immunofluorescence Microscopy
5.3. Immunoblotting and Antibody Validation
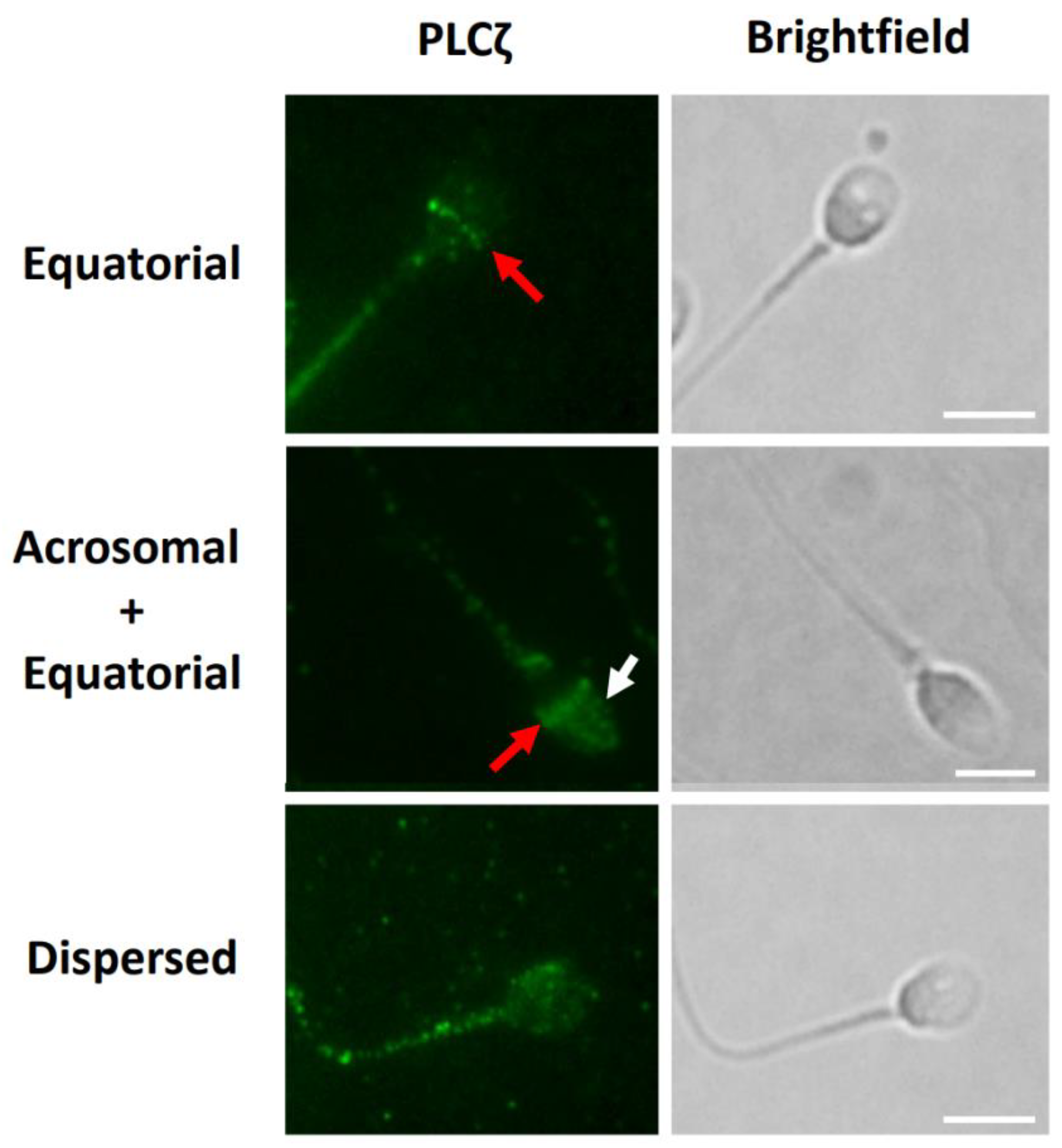
5.4. Sperm PLCζ Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kashir, J.; Heindryckx, B.; Jones, C.; De Sutter, P.; Parrington, J.; Coward, K. Oocyte activation, phospholipase C zeta and human infertility. Hum. Reprod. Update 2010, 16, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Harton, G.L.; Tempest, H.G. Chromosomal disorders and male infertility. Asian J. Androl. 2011, 14, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotaling, J.M. Genetics of Male Infertility. Urol. Clin. North Am. 2014, 41, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, A.; Giwercman, A.; Tournaye, H.; Diemer, T.; Kopa, Z.; Dohle, G.; Krausz, C. European Association of Urology Guidelines on Male Infertility: The 2012 Update. Eur. Urol. 2012, 62, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Nomikos, M.; Lai, F.A. Phospholipase C zeta and calcium oscillations at fertilisation: The evidence, applications, and further questions. Adv. Biol. Regul. 2018, 67, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.J.; Larman, M.G.; Saunders, C.M.; Hashimoto, K.; Swann, K.; Lai, F.A. Sperm phospholipase Czeta from humans and cynomolgus monkeys triggers Ca2+ oscillations, activation and development of mouse oocytes. Reproduction 2002, 124, 611–623. [Google Scholar] [CrossRef]
- Fujimoto, S.; Yoshida, N.; Fukui, T.; Amanai, M.; Isobe, T.; Itagaki, C.; Izumi, T.; Perry, A.C.F. Mammalian phospholipase Cζ induces oocyte activation from the sperm perinuclear matrix. Dev. Biol. 2004, 274, 370–383. [Google Scholar] [CrossRef]
- Kurokawa, M.; Sato, K.-I.; Wu, H.; He, C.; Malcuit, C.; Black, S.J.; Fukami, K.; Fissore, R.A. Functional, biochemical, and chromatographic characterization of the complete [Ca2+]i oscillation-inducing activity of porcine sperm. Dev. Biol. 2005, 285, 376–392. [Google Scholar] [CrossRef] [Green Version]
- Kouchi, Z.; Fukami, K.; Shikano, T.; Oda, S.; Nakamura, Y.; Takenawa, T.; Miyazaki, S. Recombinant Phospholipase Cζ Has High Ca2+ Sensitivity and Induces Ca2+ Oscillations in Mouse Eggs. J. Biol. Chem. 2004, 279, 10408–10412. [Google Scholar] [CrossRef] [Green Version]
- Knott, J.G.; Kurokawa, M.; Fissore, R.A.; Schultz, R.M.; Williams, C.J. Transgenic RNA Interference Reveals Role for Mouse Sperm Phospholipase Cζ in Triggering Ca2+ Oscillations During Fertilization1. Biol. Reprod. 2005, 72, 992–996. [Google Scholar] [CrossRef]
- Hachem, A.; Godwin, J.; Ruas, M.; Lee, H.C.; Buitrago, M.F.; Ardestani, G.; Bassett, A.; Fox, S.; Navarrete, F.; de Sutter, P.; et al. PLCζ is the physiological trigger of the Ca2+ oscillations that induce embryogenesis in mammals but offspring can be conceived in its absence. Development 2017, 144, 2914–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozawa, K.; Satouh, Y.; Fujimoto, T.; Oji, A.; Ikawa, M. Sperm-borne phospholipase C zeta-1 ensures monospermic fertilization in mice. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.-Y.; Jellerette, T.; Salicioni, A.M.; Lee, H.C.; Yoo, M.-S.; Coward, K.; Parrington, J.; Grow, D.; Cibelli, J.B.; Visconti, P.E.; et al. Human sperm devoid of PLC, zeta 1 fail to induce Ca2+ release and are unable to initiate the first step of embryo development. J. Clin. Investig. 2008, 118, 3671–3681. [Google Scholar] [CrossRef] [Green Version]
- Escoffier, J.; Lee, H.C.; Yassine, S.; Zouari, R.; Martinez, G.; Karaouzène, T.; Coutton, C.; Kherraf, Z.-E.; Halouani, L.; Triki, C.; et al. Homozygous mutation of PLCZ1 leads to defective human oocyte activation and infertility that is not rescued by the WW-binding protein PAWP. Hum. Mol. Genet. 2015, 25, 878–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer-Vaquer, A.; Barragan, M.; Freour, T.; Vernaeve, V.; Vassena, R. PLCζ sequence, protein levels, and distribution in human sperm do not correlate with semen characteristics and fertilization rates after ICSI. J. Assist. Reprod. Genet. 2016, 33, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heytens, E.; Parrington, J.; Coward, K.; Young, C.; Lambrecht, S.; Yoon, S.Y.; Fissore, R.A.; Hamer, R.; Deane, C.M.; Ruas, M.; et al. Reduced amounts and abnormal forms of phospholipase C zeta (PLC ) in spermatozoa from infertile men. Hum. Reprod. 2009, 24, 2417–2428. [Google Scholar] [CrossRef] [Green Version]
- Kashir, J.; Jones, C.; Lee, H.C.; Rietdorf, K.; Nikiforaki, D.; Durrans, C.; Ruas, M.; Tee, S.T.; Heindryckx, B.; Galione, A.; et al. Loss of activity mutations in phospholipase C zeta (PLC ) abolishes calcium oscillatory ability of human recombinant protein in mouse oocytes. Hum. Reprod. 2011, 26, 3372–3387. [Google Scholar] [CrossRef] [Green Version]
- Kashir, J.; Jones, C.; Mounce, G.; Ramadan, W.M.; Lemmon, B.; Heindryckx, B.; de Sutter, P.; Parrington, J.; Turner, K.; Child, T.; et al. Variance in total levels of phospholipase C zeta (PLC-ζ) in human sperm may limit the applicability of quantitative immunofluorescent analysis as a diagnostic indicator of oocyte activation capability. Fertil. Steril. 2013, 99, 107–117.e103. [Google Scholar] [CrossRef]
- Kashir, J.; Konstantinidis, M.; Jones, C.; Heindryckx, B.; De Sutter, P.; Parrington, J.; Wells, D.; Coward, K. Characterization of two heterozygous mutations of the oocyte activation factor phospholipase C zeta (PLCζ) from an infertile man by use of minisequencing of individual sperm and expression in somatic cells. Fertil. Steril. 2012, 98, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Konstantinidis, M.; Jones, C.; Lemmon, B.; Chang Lee, H.; Hamer, R.; Heindryckx, B.; Deane, C.M.; De Sutter, P.; Fissore, R.A.; et al. A maternally inherited autosomal point mutation in human phospholipase C zeta (PLC) leads to male infertility. Hum. Reprod. 2011, 27, 222–231. [Google Scholar] [CrossRef]
- Torra-Massana, M.; Cornet-Bartolomé, D.; Barragán, M.; Durban, M.; Ferrer-Vaquer, A.; Zambelli, F.; Rodriguez, A.; Oliva, R.; Vassena, R. Novel phospholipase C zeta 1 mutations associated with fertilization failures after ICSI. Hum. Reprod. 2019, 34, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Azad, N.; Nazarian, H.; Ghaffari Novin, M.; Masteri Farahani, R.; Piryaei, A.; Heidari, M.H. Phospholipase C zeta parameters in sperm from polymorphic teratozoospermic men. Ann. Anat. –Anat. Anz. 2018, 215, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Azad, N.; Nazarian, H.; Ghaffari Novin, M.; Masteri Farahani, R.; Piryaei, A.; Heidari, M.H.; Abdollahpour Alitappeh, M. Oligoasthenoteratozoospermic (OAT) men display altered phospholipase C ζ (PLCζ) localization and a lower percentage of sperm cells expressing PLCζ and post-acrosomal sheath WW domain-binding protein (PAWP). Bosn. J. Basic Med. Sci. 2017, 18, 178. [Google Scholar] [CrossRef] [Green Version]
- Escoffier, J.; Yassine, S.; Lee, H.C.; Martinez, G.; Delaroche, J.; Coutton, C.; Karaouzène, T.; Zouari, R.; Metzler-Guillemain, C.; Pernet-Gallay, K.; et al. Subcellular localization of phospholipase Cζ in human sperm and its absence in DPY19L2-deficient sperm are consistent with its role in oocyte activation. MHR Basic Sci. Reprod. Med. 2014, 21, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Nikiforaki, D.; Vanden Meerschaut, F.; De Gheselle, S.; Qian, C.; Van den Abbeel, E.; De Vos, W.H.; Deroo, T.; De Sutter, P.; Heindryckx, B. Sperm involved in recurrent partial hydatidiform moles cannot induce the normal pattern of calcium oscillations. Fertil. Steril. 2014, 102, 581–588.e581. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, S.K.; Kim, J.; Kim, J.H.; Chang, J.H.; Jee, B.C.; Kim, S.H. Relationship between phospholipase C zeta immunoreactivity and DNA fragmentation and oxidation in human sperm. Obstet. Gynecol. Sci. 2015, 58, 232. [Google Scholar] [CrossRef]
- Tavalaee, M.; Kiani-Esfahani, A.; Nasr-Esfahani, M.H. Relationship between phospholipase C-zeta, semen parameters, and chromatin status. Syst. Biol. Reprod. Med. 2017, 63, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Tavalaee, M.; Nomikos, M.; Lai, F.A.; Nasr-Esfahani, M.H. Expression of sperm PLCζ and clinical outcomes of ICSI-AOA in men affected by globozoospermia due to DPY19L2 deletion. Reprod. BioMedicine Online 2018, 36, 348–355. [Google Scholar] [CrossRef]
- Yelumalai, S.; Yeste, M.; Jones, C.; Amdani, S.N.; Kashir, J.; Mounce, G.; Da Silva, S.J.M.; Barratt, C.L.; McVeigh, E.; Coward, K. Total levels, localization patterns, and proportions of sperm exhibiting phospholipase C zeta are significantly correlated with fertilization rates after intracytoplasmic sperm injection. Fertil. Steril. 2015, 104, 561–568.e564. [Google Scholar] [CrossRef]
- Kashir, J.; Mistry, B.V.; BuSaleh, L.; Abu-Dawas, R.; Nomikos, M.; Ajlan, A.; Abu-Dawud, R.; AlYacoub, N.; AlHassan, S.; Lai, F.A.; et al. Phospholipase C zeta profiles are indicative of optimal sperm parameters and fertilisation success in patients undergoing fertility treatment. Andrology 2020, 8, 1143–1159. [Google Scholar] [CrossRef]
- Kashir, J. Increasing associations between defects in phospholipase C zeta and conditions of male infertility: Not just ICSI failure? J. Assist. Reprod. Genet. 2020, 37, 1273–1293. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Buntwal, L.; Nomikos, M.; Calver, B.L.; Stamatiadis, P.; Ashley, P.; Vassilakopoulou, V.; Sanders, D.; Knaggs, P.; Livaniou, E.; et al. Antigen unmasking enhances visualization efficacy of the oocyte activation factor, phospholipase C zeta, in mammalian sperm. Mol. Hum. Reprod. 2016, 23, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M.; Jones, C.; Amdani, S.N.; Yelumalai, S.; Mounce, G.; da Silva, S.J.M.; Child, T.; Coward, K. Does advancing male age influence the expression levels and localisation patterns of phospholipase C zeta (PLCζ) in human sperm? Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, A.; Kashir, J.; Thanassoulas, A.; Safieh-Garabedian, B.; Lai, F.A.; Nomikos, M. Essential Role of Sperm-Specific PLC-Zeta in Egg Activation and Male Factor Infertility: An Update. Front Cell Dev. Biol. 2020, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Jones, C.; Melo, P.; Ross, C.; Mounce, G.; Child, T.; Coward, K. Antigen unmasking does not improve the visualization of phospholipase C zeta in human spermatozoa. Asian J. Androl. 2022, 24, 345–352. [Google Scholar] [CrossRef]
- Nomikos, M.; Stamatiadis, P.; Sanders, J.R.; Beck, K.; Calver, B.L.; Buntwal, L.; Lofty, M.; Sideratou, Z.; Swann, K.; Lai, F.A. Male infertility-linked point mutation reveals a vital binding role for the C2 domain of sperm PLCζ. Biochem. J. 2017, 474, 1003–1016. [Google Scholar] [CrossRef]
- Javadian-Elyaderani, S.; Ghaedi, K.; Tavalaee, M.; Rabiee, F.; Deemeh, M.R.; Nasr-Esfahani, M.H. Diagnosis of genetic defects through parallel assessment of PLCζ and CAPZA3 in infertile men with history of failed oocyte activation. Iran J. Basic Med. Sci. 2016, 19, 281–289. [Google Scholar]
- Dai, J.; Dai, C.; Guo, J.; Zheng, W.; Zhang, T.; Li, Y.; Lu, C.; Gong, F.; Lu, G.; Lin, G. Novel homozygous variations in PLCZ1 lead to poor or failed fertilization characterized by abnormal localization patterns of PLCζ in sperm. Clin. Genet. 2019, 97, 347–351. [Google Scholar] [CrossRef]
- Janghorban-Laricheh, E.; Ghazavi-Khorasgani, N.; Tavalaee, M.; Zohrabi, D.; Abbasi, H.; Nasr- Esfahani, M.H. An association between sperm PLCζ levels and varicocele? J. Assist. Reprod. Genet. 2016, 33, 1649–1655. [Google Scholar] [CrossRef]
- Tavalaee, M.; Nasr-Esfahani, M.H. Expression profile of PLCζ,PAWP, andTR-KIT in association with fertilization potential, embryo development, and pregnancy outcomes in globozoospermic candidates for intra-cytoplasmic sperm injection and artificial oocyte activation. Andrology 2016, 4, 850–856. [Google Scholar] [CrossRef] [Green Version]
- Yeste, M.; Recuero, S.; Maside, C.; Salas-Huetos, A.; Bonet, S.; Pinart, E. Blocking NHE Channels Reduces the Ability of In Vitro Capacitated Mammalian Sperm to Respond to Progesterone Stimulus. Int. J. Mol. Sci. 2021, 22, 12646. [Google Scholar] [CrossRef]
- Aras-Tosun, D.; Cakar, Z.; Can, A.; Ozkavukcu, S.; Kaplanoglu, I.; Cinar, O. Phospholipase C-zeta levels are not correlated with fertilisation rates in infertile couples. Andrologia 2022, 54, e14269. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Sermondade, N.; Sifer, C.; Oo, S.L.; Jones, C.; Mounce, G.; Turner, K.; Child, T.; McVeigh, E.; Coward, K. Motile sperm organelle morphology evaluation-selected globozoospermic human sperm with an acrosomal bud exhibits novel patterns and higher levels of phospholipase C zeta. Hum. Reprod. 2012, 27, 3150–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sermondade, N.; Hafhouf, E.; Dupont, C.; Bechoua, S.; Palacios, C.; Eustache, F.; Poncelet, C.; Benzacken, B.; Levy, R.; Sifer, C. Successful childbirth after intracytoplasmic morphologically selected sperm injection without assisted oocyte activation in a patient with globozoospermia. Hum. Reprod. 2011, 26, 2944–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashir, J.; Nomikos, M.; Lai, F.A.; Swann, K. Sperm-induced Ca2+ release during egg activation in mammals. Biochem. Biophys. Res. Commun. 2014, 450, 1204–1211. [Google Scholar] [CrossRef]
- Auger, J.; Eustache, F.; Ducot, B.; Blandin, T.; Daudin, M.; Diaz, I.; Matribi, S.E.; Gony, B.; Keskes, L.; Kolbezen, M.; et al. Intra- and inter-individual variability in human sperm concentration, motility and vitality assessment during a workshop involving ten laboratories. Hum. Reprod. 2000, 15, 2360–2368. [Google Scholar] [CrossRef] [Green Version]
- Mossman, J.A.; Pearson, J.T.; Moore, H.D.; Pacey, A.A. Variation in mean human sperm length is linked with semen characteristics. Hum. Reprod. 2013, 28, 22–32. [Google Scholar] [CrossRef]
- Ozelci, R.; Yılmaz, S.; Dilbaz, B.; Akpınar, F.; Akdag Cırık, D.; Dilbaz, S.; Ocal, A. Seasonal variation of human sperm cells among 4,422 semen samples: A retrospective study in Turkey. Syst. Biol. Reprod. Med. 2016, 62, 379–386. [Google Scholar] [CrossRef]
- Organisation, W.H. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO Press: Geneva, Switzerland, 2010. [Google Scholar]
- Nomikos, M.; Sanders, J.R.; Parthimos, D.; Buntwal, L.; Calver, B.L.; Stamatiadis, P.; Smith, A.; Clue, M.; Sideratou, Z.; Swann, K.; et al. Essential Role of the EF-hand Domain in Targeting Sperm Phospholipase Cζ to Membrane Phosphatidylinositol 4,5-Bisphosphate (PIP2). J. Biol. Chem. 2015, 290, 29519–29530. [Google Scholar] [CrossRef] [Green Version]
- Nomikos, M.; Theodoridou, M.; Elgmati, K.; Parthimos, D.; Calver, B.L.; Buntwal, L.; Nounesis, G.; Swann, K.; Lai, F.A. Human PLC exhibits superior fertilization potency over mouse PLC in triggering the Ca2+ oscillations required for mammalian oocyte activation. Mol. Hum. Reprod. 2014, 20, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Nomikos, M.; Yu, Y.; Elgmati, K.; Theodoridou, M.; Campbell, K.; Vassilakopoulou, V.; Zikos, C.; Livaniou, E.; Amso, N.; Nounesis, G.; et al. Phospholipase Cζ rescues failed oocyte activation in a prototype of male factor infertility. Fertil. Steril. 2013, 99, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashir, J.; Mistry, B.V.; BuSaleh, L.; Nomikos, M.; Almuqayyil, S.; Abu-Dawud, R.; AlYacoub, N.; Hamdan, H.; AlHassan, S.; Lai, F.A.; et al. Antigen Unmasking Is Required to Clinically Assess Levels and Localisation Patterns of Phospholipase C Zeta in Human Sperm. Pharmaceuticals 2023, 16, 198. https://doi.org/10.3390/ph16020198
Kashir J, Mistry BV, BuSaleh L, Nomikos M, Almuqayyil S, Abu-Dawud R, AlYacoub N, Hamdan H, AlHassan S, Lai FA, et al. Antigen Unmasking Is Required to Clinically Assess Levels and Localisation Patterns of Phospholipase C Zeta in Human Sperm. Pharmaceuticals. 2023; 16(2):198. https://doi.org/10.3390/ph16020198
Chicago/Turabian StyleKashir, Junaid, Bhavesh V. Mistry, Lujain BuSaleh, Michail Nomikos, Sarah Almuqayyil, Raed Abu-Dawud, Nadya AlYacoub, Hamdan Hamdan, Saad AlHassan, F. Anthony Lai, and et al. 2023. "Antigen Unmasking Is Required to Clinically Assess Levels and Localisation Patterns of Phospholipase C Zeta in Human Sperm" Pharmaceuticals 16, no. 2: 198. https://doi.org/10.3390/ph16020198