
Chemical and Biological Investigations of Allium scorodoprasum L. Flower Extracts

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Composition of A. scorodoprasum Flower Extracts as Influenced by the Extraction Solvents
2.2. Identification of Microbes and Antimicrobial Activity
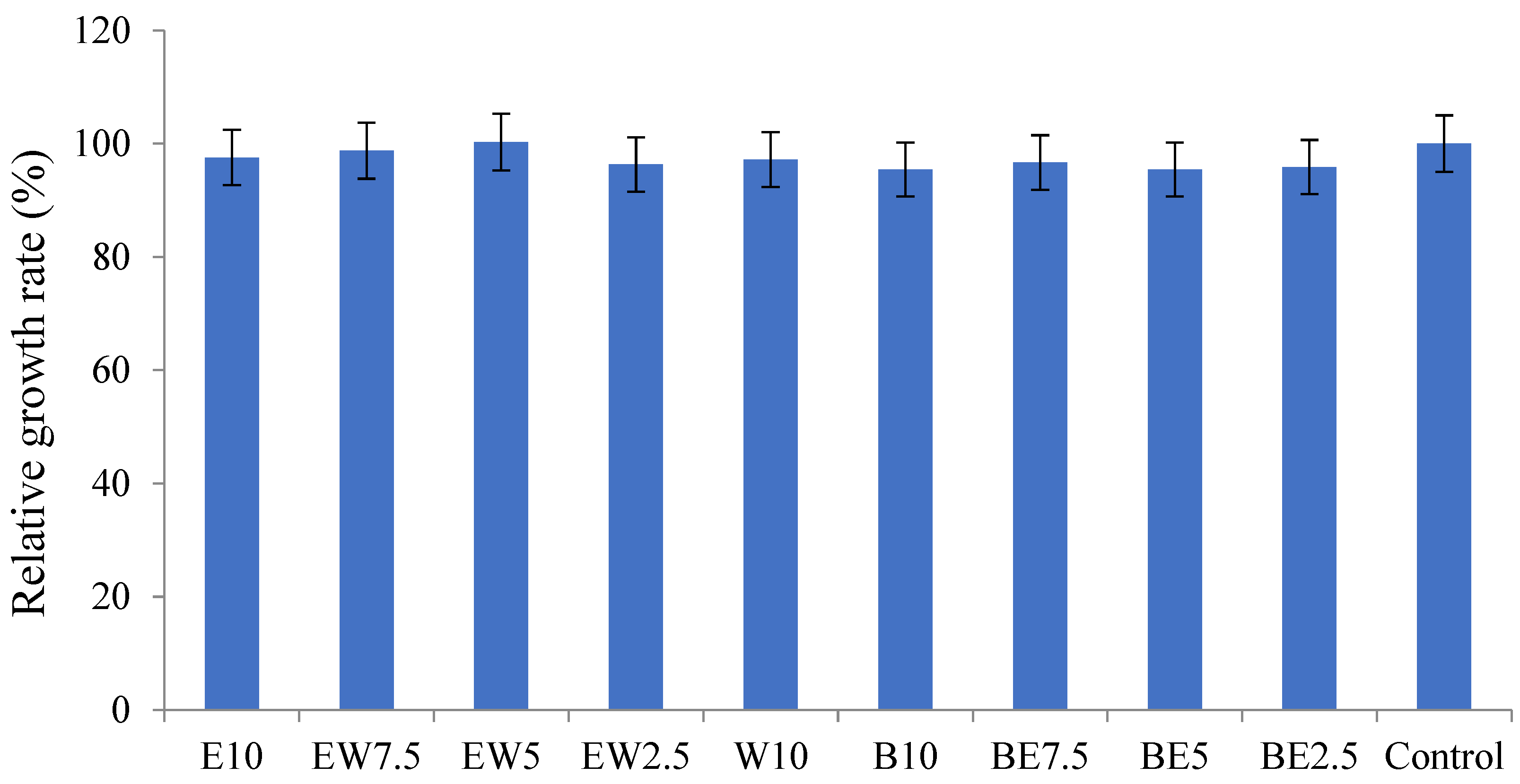
2.3. Cytotoxicity and Wound-Healing Properties of A. scorodoprasum
2.4. Antioxidant Properties of the Extracts
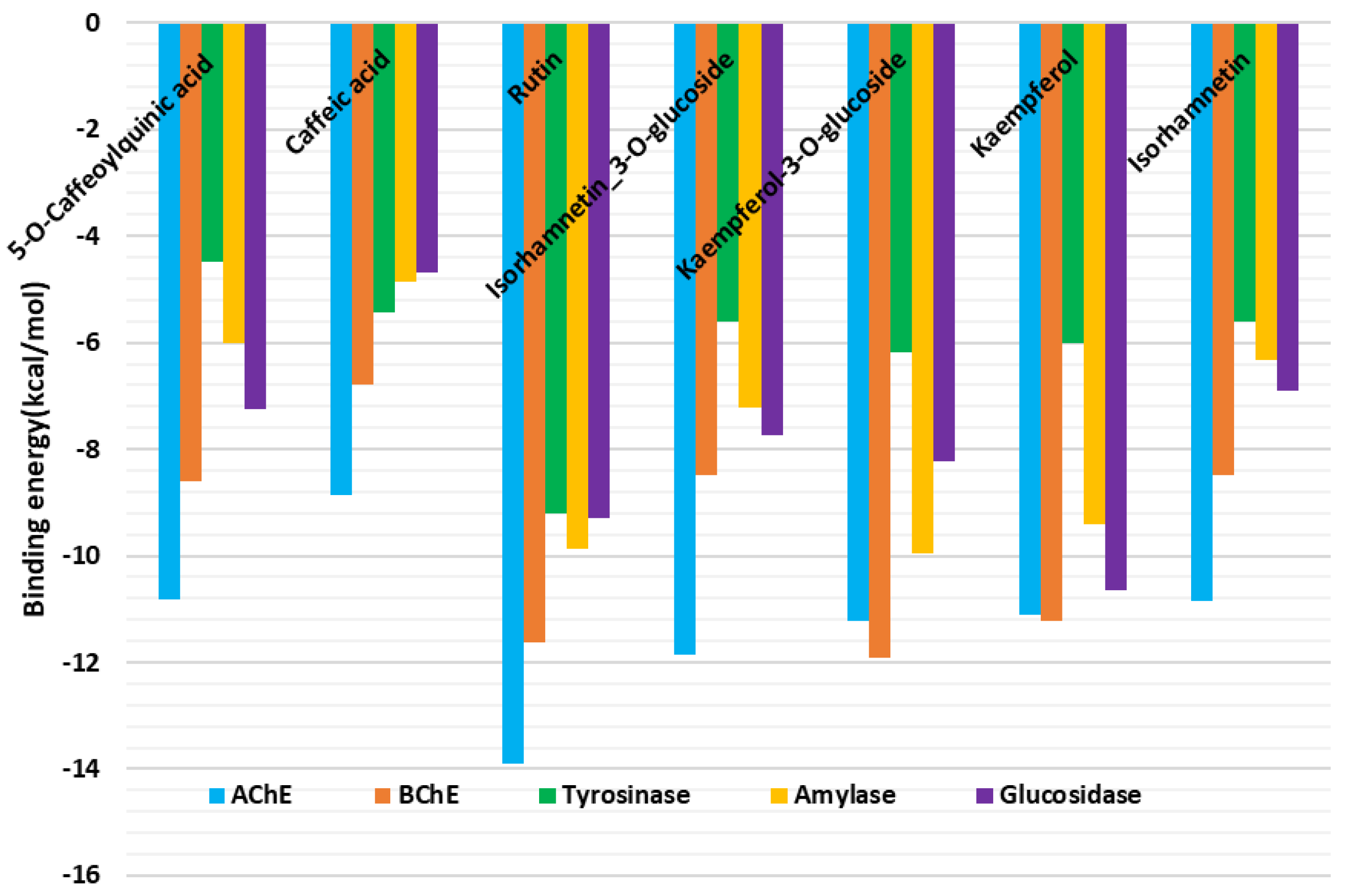
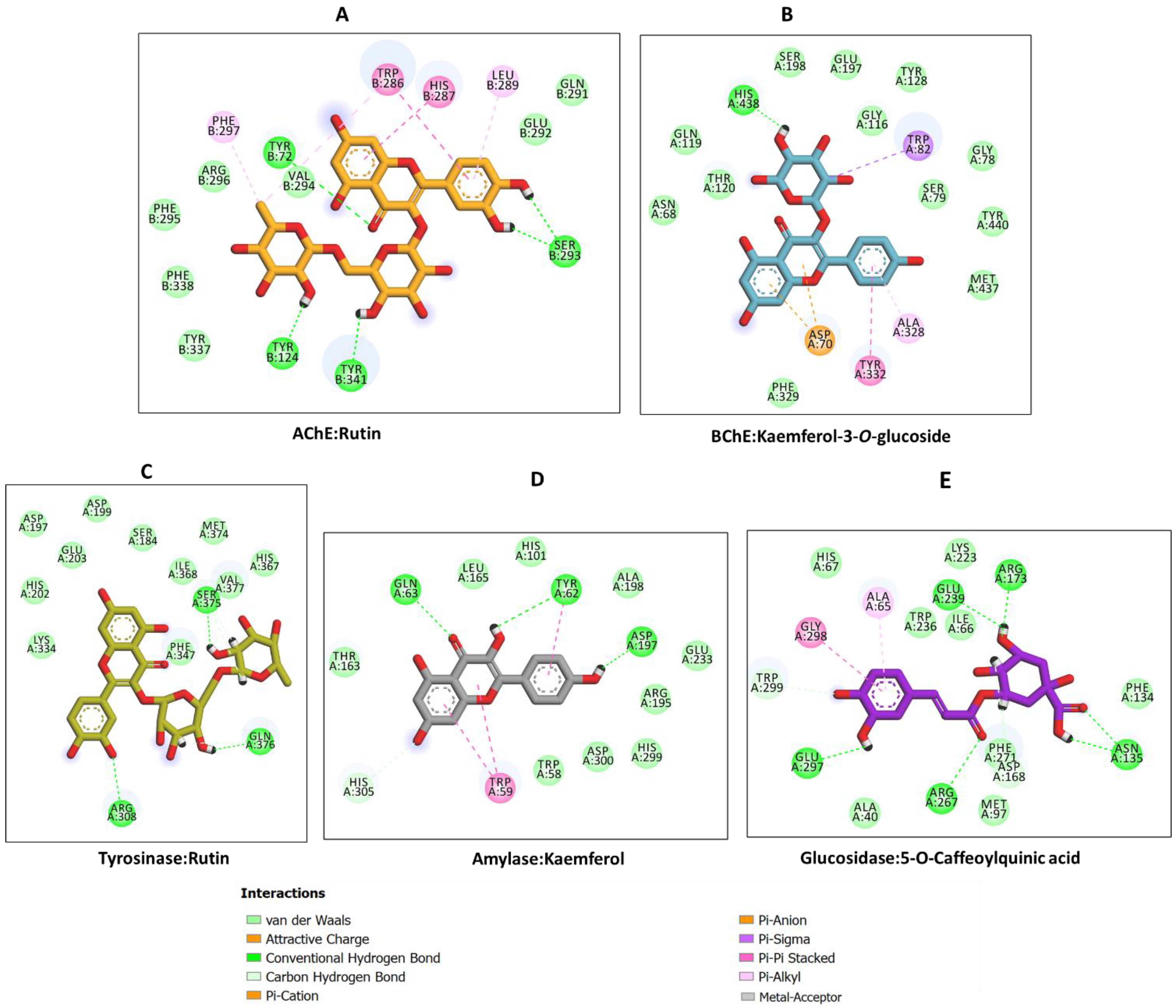
2.5. Inhibitory Activity of Allium scorodoprasum Extracts on Enzymes Linked with the Treatment of Neurodegenerative Diseases and Type 2 Diabetes and Molecular Docking
3. Materials and Methods
3.1. Plant Collection
3.2. Extractions
3.3. UHPLC(−)HESI–QqQ-MS/MS Targeted Metabolomics Analysis
3.4. Antimicrobial Assays
3.4.1. Isolation and Identification of Pathogenic Bacteria and Fungi Isolated from Infected Skin and Corneal Formations of Human Subjects
3.4.2. Antibacterial and Anticandidal Activity
3.4.3. Antifungal Activity
3.4.4. Activity of the Extracts against Biofilm Formation in S. lugdunensis
3.4.5. The Activity of Extracts against Formed Biofilms of S. lugdunensis
3.5. Cytotoxicity towards HaCaT Cell Line
3.6. In Vitro Wound-Healing Assay on HaCaT Cell Line
3.7. Antioxidant Activity Assays
3.8. Anti-Enzymatic Activities
3.9. Molecular Modeling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jarić, S.; Mitrović, M.; Djurdjević, L.; Kostić, O.; Gajić, G.; Pavlović, D.; Pavlović, P. Phytotherapy in medieval Serbian medicine according to the pharmacological manuscripts of the Chilandar Medical Codex (15-16th centuries). J. Ethnopharmacol. 2011, 137, 601–619. [Google Scholar] [CrossRef] [PubMed]
- Dusanka, P.; Dragan, S. The Chilander Medical Codex: Its importance for the medieval Serbian pharmacy. Pharm. Hist. Aust. 2001, 14, 12854602. [Google Scholar]
- Djordjević, S.P. Hilandarski medicinski kodeks. Srp. Arh. Celok. Lek. 1980, 108, 961–967. [Google Scholar] [PubMed]
- Barile, E.; Capasso, R.; Izzo, A.A.; Lanzotti, V.; Sajjadi, S.E.; Zolfaghari, B. Structure-activity relationships for saponins from Allium hirtifolium and Allium elburzense and their antispasmodic activity. Planta Med. 2005, 71, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Elgorashi, E.E.; Van Staden, J. Bioactivity and bioactive compounds of African amaryllidaceae. ACS Symp. Ser. 2009, 1021, 151–170. [Google Scholar] [CrossRef]
- Fenwick, G.R.; Hanley, A.B. The genus Allium–Part 1. Crit. Rev. Food Sci. Nutr. 1985, 22, 199–271. [Google Scholar] [CrossRef]
- Fenwick, G.R.; Hanley, A.B. The genus Allium–Part 2. Crit. Rev. Food Sci. Nutr. 1985, 22, 273–377. [Google Scholar] [CrossRef]
- Teshika, J.D.; Zakariyyah, A.M.; Zaynab, T.; Zengin, G.; Rengasamy, K.R.; Pandian, S.K.; Fawzi, M.M. Traditional and modern uses of onion bulb (Allium cepa L.): A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59, S39–S70. [Google Scholar] [CrossRef]
- Tasci, B.; Koca, I. Use of Allium scorodoprasum L. subsp. rotundum as food. Acta Hortic. 2016, 1143, 153–158. [Google Scholar] [CrossRef]
- Oosthuizen, C.B.; Reid, A.M.; Lall, N. Garlic (Allium sativum) and Its Associated Molecules, as Medicine. In Medicinal Plants for Holistic Health and Well-Being; Lall, N., Ed.; Academic Press: London, UK, 2018; pp. 277–295. ISBN 9780128124758. [Google Scholar] [CrossRef]
- FIRAT, M. The Ethnobotanical Usage of Some East Anatolian (Turkey) Allium L. Species. Manas J. Agric. Vet. Life Sci. 2015, 5, 80–86. [Google Scholar]
- Putnik, P.; Gabrić, D.; Roohinejad, S.; Barba, F.J.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.M.; Bursać Kovačević, D. An overview of organosulfur compounds from Allium spp.: From processing and preservation to evaluation of their bioavailability, antimicrobial, and anti-inflammatory properties. Food Chem. 2019, 276, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Hansen, E.A.; Folts, J.D.; Goldman, I.L. Steam-cooking rapidly destroys and reverses onion-induced antiplatelet activity. Nutr. J. 2012, 11, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyung, K.H. Antimicrobial properties of allium species. Curr. Opin. Biotechnol. 2012, 23, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Poojary, M.M.; Putnik, P.; Bursać Kovačević, D.; Barba, F.J.; Lorenzo, J.M.; Dias, D.A.; Shpigelman, A. Stability and extraction of bioactive sulfur compounds from Allium genus processed by traditional and innovative technologies. J. Food Compos. Anal. 2017, 61, 28–39. [Google Scholar] [CrossRef]
- Snyder, H.M.; Perlman, W.R.; Egge, R.; Carrillo, M.C. Alzheimer’s disease public-private partnerships: Update 2017. Alzheimer’s Dement. 2018, 14, 522–531. [Google Scholar] [CrossRef]
- Pugazhenthi, S.; Qin, L.; Reddy, P.H. Common neurodegenerative pathways in obesity, diabetes, and Alzheimer’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1037–1045. [Google Scholar] [CrossRef]
- Spampinato, S.F.; Caruso, G.I.; De Pasquale, R.; Sortino, M.A.; Merlo, S. The Treatment of Impaired Wound Healing in Diabetes: Looking among Old Drugs. Pharmaceuticals 2020, 13, 60. [Google Scholar] [CrossRef] [Green Version]
- Delamaire, M.; Maugendre, D.; Moreno, M.; Le Goff, M.-C.; Allannic, H.; Genetet, B. Impaired Leucocyte Functions in Diabetic Patients. Diabet. Med. 1997, 14, 29–34. [Google Scholar] [CrossRef]
- Dubey, R.; Prabhakar, P.K.; Gupta, J. Epigenetics: Key to improve delayed wound healing in type 2 diabetes. Mol. Cell. Biochem. 2022, 477, 371–383. [Google Scholar] [CrossRef]
- Kaye, K.S.; Petty, L.A.; Shorr, A.F.; Zilberberg, M.D. Current Epidemiology, Etiology, and Burden of Acute Skin Infections in the United States. Clin. Infect. Dis. 2019, 68, S193–S199. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Noviello, S.; Leone, S. Epidemiology and microbiology of skin and soft tissue infections. Curr. Opin. Infect. Dis. 2016, 29, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Božunović, J.; Gašić, U.; Drakulić, D.; Stevanović, M.; Rajčević, N.; Stojković, D. Bioactivities of Salvia nemorosa L. inflorescences are influenced by the extraction solvents. Ind. Crops Prod. 2022, 175, 114260. [Google Scholar] [CrossRef]
- Demasi, S.; Caser, M.; Donno, D.; Enri, S.R.; Lonati, M.; Scariot, V. Exploring wild edible flowers as a source of bioactive compounds: New perspectives in horticulture. Folia Hortic. 2021, 33, 27–48. [Google Scholar] [CrossRef]
- Emir, A.; Emir, C.; Yıldırım, H. Chemical and biological comparison of different parts of two Allium species: Allium paniculatum L. subsp. villosulum (Hal.) Stearn and Allium paniculatum L. subsp. paniculatum L. Chem. Pap. 2021, 75, 411–419. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Locatelli, M.; Picot-Allain, C.M.N.; Mahomoodally, M.F. Multidirectional investigations on different parts of Allium scorodoprasum L. subsp. rotundum (L.) Stearn: Phenolic components, in vitro biological, and in silico propensities. Food Res. Int. 2018, 108, 641–649. [Google Scholar] [CrossRef]
- Nowicka, D.; Chilicka, K.; Dzieńdziora-Urbińska, I. Host-Microbe Interaction on the Skin and Its Role in the Pathogenesis and Treatment of Atopic Dermatitis. Pathogens 2022, 11, 71. [Google Scholar] [CrossRef]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; Mullikin, J.; et al. Temporal shifts in the skin microbiome associated with disease flares and treatment in children with atopic dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Taiwo, F.O.; Oyedeji, O.; Osundahunsi, M.T. Antimicrobial and Antioxidant Properties of kaempferol-3-O-glucoside and 1-(4-Hydroxyphenyl)-3-phenylpropan-1-one Isolated from the Leaves of Annona muricata (Linn.). J. Pharm. Res. Int. 2019, 3, 1–13. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2017, 25, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Sathiya Deepika, M.; Thangam, R.; Sakthidhasan, P.; Arun, S.; Sivasubramanian, S.; Thirumurugan, R. Combined effect of a natural flavonoid rutin from Citrus sinensis and conventional antibiotic gentamicin on Pseudomonas aeruginosa biofilm formation. Food Control 2018, 90, 282–294. [Google Scholar] [CrossRef]
- Wang, Z.; Ding, Z.; Li, Z.; Ding, Y.; Jiang, F.; Liu, J. Antioxidant and antibacterial study of 10 flavonoids revealed rutin as a potential antibiofilm agent in Klebsiella pneumoniae strains isolated from hospitalized patients. Microb. Pathog. 2021, 159, 105121. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Flavones, flavonols, and glycosylated derivatives—Impact on candida albicans growth and virulence, expression of cdr1 and erg11, cytotoxicity. Pharmaceuticals 2021, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Al-Shabib, N.A.; Husain, F.M.; Ahmad, I.; Khan, M.S.; Khan, R.A.; Khan, J.M. Rutin inhibits mono and multi-species biofilm formation by foodborne drug resistant Escherichia coli and Staphylococcus aureus. Food Control 2017, 79, 325–332. [Google Scholar] [CrossRef]
- Stojković, D.; Dias, M.I.; Drakulić, D.; Barros, L.; Stevanović, M.; Ferreira, I.C.F.R.; Soković, M.D. Methanolic extract of the Herb ononis spinosa L. Is an antifungal agent with no cytotoxicity to primary human cells. Pharmaceuticals 2020, 13, 78. [Google Scholar] [CrossRef]
- Chen, L.Y.; Huang, C.N.; Liao, C.K.; Chang, H.M.; Kuan, Y.H.; Tseng, T.J.; Yen, K.J.; Yang, K.L.; Lin, H.C. Effects of Rutin on Wound Healing in Hyperglycemic Rats. Antioxidants 2020, 9, 1122. [Google Scholar] [CrossRef]
- Özay, Y.; Güzel, S.; Yumrutaş, Ö.; Pehlivanoğlu, B.; Erdoğdu, İ.H.; Yildirim, Z.; Türk, B.A.; Darcan, S. Wound Healing Effect of Kaempferol in Diabetic and Nondiabetic Rats. J. Surg. Res. 2019, 233, 284–296. [Google Scholar] [CrossRef]
- Ambiga, S.; Narayanan, R.; Gowri, D.; Sukumar, D.; Madhavan, S. Evaluation of wound healing activity of flavonoids from ipomoea carnea Jacq. Anc. Sci. Life 2007, 26, 45–51. [Google Scholar]
- Demir, T.; Akpınar, Ö.; Kara, H.; Güngör, H. Phenolic profile and investigation of biological activities of Allium scorodoprasum L. subsp. rotundum. Food Biosci. 2022, 46, 101548. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K.; Zapp, J. Activity-guided isolation of cholinesterase inhibitors quercetin, rutin and kaempferol from Prunus persica fruit. Z. Naturforsch. C 2020, 75, 87–96. [Google Scholar] [CrossRef]
- Khan, M.T.H.; Orhan, I.; Şenol, F.S.; Kartal, M.; Şener, B.; Dvorská, M.; Šmejkal, K.; Šlapetová, T. Cholinesterase inhibitory activities of some flavonoid derivatives and chosen xanthone and their molecular docking studies. Chem. Biol. Interact. 2009, 181, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Jakimiuk, K.; Sari, S.; Milewski, R.; Supuran, C.T.; Şöhretoğlu, D.; Tomczyk, M. Flavonoids as tyrosinase inhibitors in in silico and in vitro models: Basic framework of SAR using a statistical modelling approach. J. Enzyme Inhib. Med. Chem. 2022, 37, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Ai, B.; Zheng, L.; Zheng, X.; Xu, Z.; Shen, Y.; Jin, Z. Inhibitory activities of kaempferol, galangin, carnosic acid and polydatin against glycation and α-amylase and α-glucosidase enzymes. Int. J. Food Sci. Technol. 2018, 53, 755–766. [Google Scholar] [CrossRef]
- Yin, P.; Yang, L.; Xue, Q.; Yu, M.; Yao, F.; Sun, L.; Liu, Y. Identification and inhibitory activities of ellagic acid- and kaempferol-derivatives from Mongolian oak cups against α-glucosidase, α-amylase and protein glycation linked to type II diabetes and its complications and their influence on HepG2 cells’ viability. Arab. J. Chem. 2018, 11, 1247–1259. [Google Scholar] [CrossRef]
- Dubey, S.; Ganeshpurkar, A.; Ganeshpurkar, A.; Bansal, D.; Dubey, N. Glycolytic enzyme inhibitory and antiglycation potential of rutin. Futur. J. Pharm. Sci. 2017, 3, 158–162. [Google Scholar] [CrossRef]
- Smiljkovic, M.; Dias, M.I.; Stojkovic, D.; Barros, L.; Bukvički, D.; Ferreira, I.C.F.R.; Sokovic, M. Characterization of phenolic compounds in tincture of edible Nepeta nuda: Development of antimicrobial mouthwash. Food Funct. 2018, 9, 5417–5425. [Google Scholar] [CrossRef] [Green Version]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef]
- Gerlits, O.; Ho, K.Y.; Cheng, X.; Blumenthal, D.; Taylor, P.; Kovalevsky, A.; Radić, Z. A new crystal form of human acetylcholinesterase for exploratory room-temperature crystallography studies. Chem. Biol. Interact. 2019, 309, 108698. [Google Scholar] [CrossRef]
- Rosenberry, T.L.; Brazzolotto, X.; MacDonald, I.R.; Wandhammer, M.; Trovaslet-Leroy, M.; Darvesh, S.; Nachon, F. Comparison of the Binding of Reversible Inhibitors to Human Butyrylcholinesterase and Acetylcholinesterase: A Crystallographic, Kinetic and Calorimetric Study. Molecules 2017, 22, 2098. [Google Scholar] [CrossRef] [Green Version]
- Maurus, R.; Begum, A.; Williams, L.K.; Fredriksen, J.R.; Zhang, R.; Withers, S.G.; Brayer, G.D. Alternative catalytic anions differentially modulate human alpha-amylase activity and specificity. Biochemistry 2008, 47, 3332–3344. [Google Scholar] [CrossRef]
- Ielo, L.; Deri, B.; Germanò, M.P.; Vittorio, S.; Mirabile, S.; Gitto, R.; Rapisarda, A.; Ronsisvalle, S.; Floris, S.; Pazy, Y.; et al. Exploiting the 1-(4-fluorobenzyl)piperazine fragment for the development of novel tyrosinase inhibitors as anti-melanogenic agents: Design, synthesis, structural insights and biological profile. Eur. J. Med. Chem. 2019, 178, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Karade, S.S.; Hill, M.L.; Kiappes, J.L.; Manne, R.; Aakula, B.; Zitzmann, N.; Warfield, K.L.; Treston, A.M.; Mariuzza, R.A. N-Substituted Valiolamine Derivatives as Potent Inhibitors of Endoplasmic reticulum α-Glucosidases I and II with Antiviral Activity. J. Med. Chem. 2021, 64, 18010–18024. [Google Scholar] [CrossRef] [PubMed]
- Omer, H.A.A.; Caprioli, G.; Abouelenein, D.; Mustafa, A.M.; Uba, A.I.; Ak, G.; Ozturk, R.B.; Zengin, G.; Yagi, S. Phenolic Profile, Antioxidant and Enzyme Inhibitory Activities of Leaves from Two Cassia and Two Senna Species. Molecules 2022, 27, 5590. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rosell, G.; Giorgino, T.; De Fabritiis, G. PlayMolecule ProteinPrepare: A Web Application for Protein Preparation for Molecular Dynamics Simulations. J. Chem. Inf. Model. 2017, 57, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Ruth, H.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, R.B.; Zengin, G.; Sinan, K.I.; Montesano, D.; Zheleva-Dimitrova, D.; Gevrenova, R.; Uba, A.I.; Çakılcıoğlu, U.; Kaplan, A.; Jugreet, S.; et al. Which Extraction Solvents and Methods Are More Effective in Terms of Chemical Composition and Biological Activity of Alcea fasciculiflora from Turkey? Molecules 2022, 27, 5011. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample Code | Extraction Solvent | Yield % (v/v) |
|---|---|---|
| E10 | Ethanol, 100% | 3.87 |
| EW7.5 | Ethanol:Water, 75%:25% | 36.99 |
| EW5 | Ethanol:Water, 50%:50% | 6.63 |
| EW2.5 | Ethanol:Water, 25%:75% | 11.42 |
| W10 | Water, 100% | 9.38 |
| B10 | Butanol, 100% | 7.03 |
| BE7.5 | Butanol:Ethanol, 75%:25% | 13.21 |
| BE5 | Butanol:Ethanol, 50%:50% | 9.05 |
| BE2.5 | Butanol:Ethanol, 25%:75% | 11.76 |
| Identified Compounds in mg/kg dw | E10 | EW7.5 | EW5 | EW2.5 | W10 | B10 | BE7.5 | BE5 | BE2.5 |
|---|---|---|---|---|---|---|---|---|---|
| 3-O-Caffeoylquinic acid | 6.29 ± 0.34 | 3.41 ± 0.20 | 12.20 ± 1.63 | 40.90 ± 0.81 | 3.98 ± 0.55 | 17.38 ± 1.11 | 1.64 ± 0.13 | 3.90 ± 0.11 | 2.32 ± 0.30 |
| 5-O-Caffeoylquinic acid | 173.75 ± 9.90 | 54.47 ± 0.82 | 211.36 ± 5.57 | 225.16 ± 1.73 | 19.93 ± 0.70 | 541.11 ± 13.11 | 10.35 ± 0.15 | 83.30 ± 1.00 | 51.62 ± 1.55 |
| Caffeic acid | 303.76 ± 6.99 | 80.10 ± 1.29 | 219.73 ± 0.78 | 82.45 ± 1.84 | 65.01 ± 2.10 | 154.46 ± 5.28 | 61.82 ± 1.66 | 80.87 ± 4.80 | 58.02 ± 2.62 |
| Isoorientin | NF | NF | NF | 128.19 ± 3.06 | NF | NF | NF | NF | NF |
| Rutin | 2504.35 ± 68.46 | 1309.00 ± 22.86 | 5921.72 ± 202.28 | 1117.89 ± 13.15 | 70.33 ± 1.80 | 1936.22 ± 47.94 | 140.85 ± 3.54 | 631.88 ± 5.25 | 477.32 ± 15.99 |
| Vitexin | 1.64 ± 0.13 | 0.44 ± 0.01 | 1.81 ± 0.25 | 24.18 ± 0.30 | 1.56 ± 0.16 | 1.76 ± 0.22 | 0.14 ± 0.01 | 0.44 ± 0.04 | 0.38 ± 0.07 |
| p-Coumaric acid | 215.96 ± 9.08 | 65.57 ± 0.76 | 192.52 ± 4.00 | 195.01 ± 3.31 | 180.16 ± 3.19 | 136.38 ± 4.16 | 14.67 ± 1.09 | 47.65 ± 2.53 | 43.39 ± 3.22 |
| Quercetin 3-O-glucoside | 136.42 ± 3.81 | 61.30 ± 0.81 | 347.91 ± 9.79 | 407.02 ± 1.29 | 17.26 ± 0.62 | 92.18 ± 3.88 | 5.49 ± 0.05 | 31.15 ± 1.26 | 26.46 ± 1.04 |
| Isorhamnetin 3-O-glucoside | 470.39 ± 0.19 | 191.56 ± 5.14 | 1673.95 ± 42.08 | 748.56 ± 16.46 | NF | 395.08 ± 17.90 | 34.78 ± 1.84 | 102.48 ± 2.68 | 133.64 ± 0.39 |
| Kaempferol 3-O-glucoside | 1774.76 ± 64.90 | 316.30 ± 3.54 | 1728.21 ± 11.31 | 1362.05 ± 30.59 | 27.60 ± 0.91 | 2348.39 ± 69.81 | 164.15 ± 5.49 | 526.13 ± 8.79 | 503.54 ± 7.91 |
| Eriodictyol | NF | 1.96 ± 0.03 | 7.27 ± 0.21 | NF | NF | NF | NF | NF | NF |
| Luteolin | 47.87 ± 3.46 | 7.94 ± 0.13 | 37.99 ± 1.95 | 105.17 ± 4.22 | 10.75 ± 0.42 | 23.85 ± 0.27 | 5.47 ± 0.20 | 11.00 ± 0.83 | 8.91 ± 0.92 |
| Quercetin | 21.52 ± 1.21 | 13.72 ± 0.16 | 50.85 ± 0.95 | 104.76 ± 4.17 | 180.37 ± 4.09 | 14.94 ± 0.89 | 1.73 ± 0.08 | 5.33 ± 0.04 | 2.92 ± 0.23 |
| Naringenin | 7.38 ± 0.51 | 1.53 ± 0.08 | 3.84 ± 0.22 | 1.51 ± 0.17 | 1.80 ± 0.14 | 4.86 ± 0.36 | 0.90 ± 0.10 | 2.25 ± 0.08 | 1.53 ± 0.17 |
| Apigenin | NF | NF | NF | 38.56 ± 2.14 | 129.01 ± 5.96 | NF | NF | NF | NF |
| Kaempferol | 1583.28 ± 111.10 | 703.46 ± 26.91 | 1427.17 ± 39.11 | 1121.81 ± 8.67 | 305.73 ± 11.30 | 1407.73 ± 58.77 | 278.41 ± 3.20 | 592.23 ± 21.26 | 383.78 ± 11.78 |
| Hispidulin | 96.59 ± 1.19 | 20.67 ± 1.59 | 58.93 ± 2.34 | 24.54 ± 0.28 | 30.78 ± 0.22 | 47.53 ± 1.94 | 16.79 ± 0.29 | 24.13 ± 0.31 | 17.34 ± 0.07 |
| Isorhamnetin | 266.40 ± 17.95 | 169.07 ± 8.97 | 565.76 ± 19.97 | 1201.72 ± 40.13 | 605.67 ± 14.12 | 282.41 ± 18.20 | 33.48 ± 1.44 | 110.04 ± 4.43 | 83.52 ± 2.14 |
| SUM | 7610.35 | 3000.50 | 12461.22 | 6929.48 | 1649.93 | 7404.29 | 770.68 | 2252.77 | 1794.68 |
| Species | Number of Isolates | Confidence Value | Label/Labels | |
|---|---|---|---|---|
| Bacteria | Staphylococcus epidermidis | 1 | 99.9 | B45 |
| Staphylococcus lugdunensis | 1 | 99.9 | B43 | |
| Proteus vulgaris | 1 | 99.9 | B44 | |
| Yeast | Candida albicans | 1 | 99.9 | Y177 |
| Candida krusei | 1 | 99.9 | Y454 | |
| Candida tropicalis | 1 | 99.9 | Y149 | |
| Candida kefyr | 1 | 99.9 | Y289 | |
| Dermatomycetes | Microsporum fulvum | 2 | 99.9 | D89 |
| D351 | ||||
| Trichophyton violaceum | 1 | 99.9 | D1182 | |
| Trichophyton mentagrophytes | 2 | 99.9 | D448 | |
| D465 | ||||
| Microsporum canis | 2 | 99.9 | D277 | |
| D371 | ||||
| Trichophyton rubrum | 2 | 99.9 | D460 | |
| D1026 |
| Extract mg/mL | Staphylococcus lugdunensis (B43) | Staphylococcus epidermidis (B45) | Proteus vulgaris (B44) | Candida albicans (Y177) | Candida krusei (Y454) | Candida tropicalis (Y149) | Candida kefyr (Y289) | |
|---|---|---|---|---|---|---|---|---|
| E10 | MIC | 1 | 0.5 | 2 | 0.06 | 1 | 1 | 1 |
| MBC/MFC | 2 | 1 | 4 | 0.125 | 2 | 2 | 2 | |
| EW7.5 | MIC | 0.5 | 0.5 | 2 | 0.125 | 2 | 0.125 | 0.125 |
| MBC/MFC | 1 | 1 | 4 | 0.25 | 4 | 0.25 | 0.25 | |
| EW5 | MIC | 0.5 | 0.5 | 1 | 0.06 | 1 | 0.06 | 0.5 |
| MBC/MFC | 1 | 1 | 2 | 0.125 | 2 | 0.125 | 1 | |
| EW2.5 | MIC | 0.5 | 1 | 2 | 0.125 | 2 | 0.25 | 0.5 |
| MBC/MFC | 1 | 2 | 4 | 0.25 | 4 | 0.5 | 1 | |
| W10 | MIC | 4 | 0.5 | 0.5 | 0.25 | 0.5 | 0.5 | 0.5 |
| MBC/MFC | 8 | 1 | 1 | 0.5 | 1 | 1 | 1 | |
| B10 | MIC | 4 | 0.5 | 0.125 | 0.5 | 0.5 | 0.5 | 0.5 |
| MBC/MFC | 8 | 1 | 0.25 | 1 | 1 | 1 | 1 | |
| BE7.5 | MIC | 1 | 0.5 | 1 | 0.5 | 0.5 | 0.5 | 0.5 |
| MBC/MFC | 2 | 1 | 2 | 1 | 1 | 1 | 1 | |
| BE5 | MIC | 4 | 0.5 | 1 | 0.5 | 0.5 | 0.5 | 0.5 |
| MBC/MFC | 8 | 1 | 2 | 1 | 1 | 1 | 1 | |
| BE2.5 | MIC | 4 | 0.5 | 4 | 0.5 | 4 | 4 | 4 |
| MBC/MFC | 8 | 1 | 8 | 1 | 8 | 8 | 8 | |
| Streptomycin | MIC | 0.003 | 0.1 | 0.003 | - | - | - | - |
| MBC | 0.006 | 0.2 | 0.006 | - | - | - | - | |
| Ketoconazole | MIC | - | - | - | 0.015 | 0.015 | 0.015 | 0.015 |
| MFC | - | - | - | 0.030 | 0.030 | 0.030 | 0.030 |
| Extract mg/mL | Microsporum fulvum (D89) | Microsporum fulvum (D351) | Trichophyton violaceum (D1182) | Trichophyton mentagrophytes (D448) | Trichophyton mentagrophytes (D465) | Microsporum canis (D277) | Microsporum canis (D371) | Trichophyton rubrum (D460) | Trichophyton rubrum (D1026) | |
|---|---|---|---|---|---|---|---|---|---|---|
| E10 | MIC | 2 | 1 | 2 | 2 | 2 | 1 | 1 | 1 | 1 |
| MFC | 4 | 2 | 4 | 4 | 4 | 2 | 2 | 2 | 2 | |
| EW7.5 | MIC | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 |
| MFC | 4 | 2 | 4 | 4 | 4 | 2 | 2 | 2 | 2 | |
| EW5 | MIC | 1 | 1 | 1 | 1 | 1 | 0.5 | 0.5 | 0.25 | 0.25 |
| MFC | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 0.5 | 0.5 | |
| EW2.5 | MIC | 1 | 2 | 2 | 1 | 1 | 0.5 | 0.5 | 0.25 | 0.25 |
| MFC | 2 | 4 | 4 | 2 | 2 | 1 | 1 | 0.5 | 0.5 | |
| W10 | MIC | 4 | 4 | 4 | 4 | 4 | 2 | 2 | 2 | 2 |
| MFC | 8 | 8 | 8 | 8 | 8 | 4 | 4 | 4 | 4 | |
| B10 | MIC | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| MFC | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | |
| BE7.5 | MIC | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| MIC | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | |
| BE5 | MIC | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| MFC | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | |
| BE2.5 | MIC | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| MFC | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | |
| Ketoconazole | MIC | 0.025 | 0.025 | 0.0125 | 0.0125 | 0.0125 | 0.25 | 0.25 | 0.0125 | 0.0125 |
| MFC | 0.050 | 0.050 | 0.0250 | 0.0250 | 0.0250 | 0.50 | 0.50 | 0.0250 | 0.0250 |
| Extract | Inhibition Percentage [%] | ||
|---|---|---|---|
| MIC/2 | MIC | MBC | |
| EW7.5 | 43.75% | 65.44% | 66.18% |
| EW5 | 46.32% | 63.97% | 70.59% |
| EW2.5 | 44.11% | 51.84% | 62.5% |
| Extract | Inhibition Procentage [%] | ||
|---|---|---|---|
| MIC | MBC | 2xMBC | |
| EW7.5 | NI | NI | NI |
| EW5 | NI | NI | NI |
| EW2.5 | NI | NI | 46.6% |
| Extract | Wound Closure (%) |
|---|---|
| E10 | 81.00 ± 0.97 |
| EW7.5 | 17.34 ± 1.74 |
| EW5 | 100.00 ± 0.00 |
| EW2.5 | 23.18 ± 4.58 |
| W10 | 23.16 ± 0.98 |
| B10 | 26.33 ± 1.22 |
| BE7.5 | 23.66 ± 2.05 |
| BE5 | 13.17 ± 2.88 |
| BE2.5 | 17.14 ± 1.11 |
| Control | 0.83 ± 0.08 |
| Extract | DPPH mg TE/g | ABTS mg TE/g | CUPRAC mg TE/g | FRAP mg TE/g | Metal Chelating mg EDTAE/g | Phosphomolybdenum mmol TE/g |
|---|---|---|---|---|---|---|
| Mean ± SD | ||||||
| E10 | 16.03 ± 0.45 | 32.56 ± 0.70 | 55.35 ± 0.61 | 49.34 ± 0.98 | 7.69 ± 0.32 | 1.79 ± 0.01 |
| EW7.5 | 16.33 ± 0.73 | 42.09 ± 0.53 | 50.86 ± 0.56 | 44.06 ± 0.04 | 16.64 ± 0.18 | 1.31 ± 0.05 |
| EW5 | 18.66 ± 0.49 | 44.04 ± 0.19 | 56.26 ± 1.37 | 48.83 ± 0.29 | 18.60 ± 0.55 | 1.83 ± 0.11 |
| EW2.5 | 15.50 ± 0.70 | 40.71 ± 1.30 | 45.27 ± 0.38 | 37.80 ± 0.64 | 16.33 ± 0.15 | 1.09 ± 0.10 |
| W10 | 12.05 ± 0.01 | 34.76 ± 0.37 | 41.31 ± 0.85 | 39.78 ± 0.77 | 12.40 ± 0.29 | 0.99 ± 0.08 |
| B10 | 12.33 ± 0.27 | 27.95 ± 0.37 | 48.17 ± 1.16 | 44.26 ± 0.89 | 8.67 ± 0.47 | 1.30 ± 0.04 |
| BE7.5 | 10.92 ± 0.32 | 27.28 ± 0.52 | 43.07 ± 0.81 | 40.03 ± 0.54 | 6.11 ± 0.70 | 1.13 ± 0.08 |
| BE5 | 2.50 ± 0.34 | 8.46 ± 0.86 | 20.94 ± 0.87 | 18.16 ± 0.17 | 16.76 ± 0.67 | 0.60 ± 0.02 |
| BE2.5 | 3.37 ± 0.72 | 10.35 ± 1.42 | 22.36 ± 0.72 | 19.72 ± 0.26 | 11.12 ± 1.04 | 0.56 ± 0.02 |
| Extract | AChE mg GALAE/g | BChE mg GALAE/g | Tyrosinase mg KAE/g | Amylase mmol ACAE/g | Glucosidase mmol ACAE/g |
|---|---|---|---|---|---|
| Mean ± SD na: not active | |||||
| E10 | 3.05 ± 0.03 | 3.50 ± 0.16 | 65.24 ± 3.20 | 0.28 ± 0.01 | 0.86 ± 0.02 |
| EW7.5 | 3.06 ± 0.03 | 2.61 ± 0.14 | 53.24 ± 2.29 | 0.24 ± 0.00 | 0.90 ± 0.01 |
| EW5 | 2.99 ± 0.07 | 1.81 ± 0.23 | 43.96 ± 5.15 | 0.20 ± 0.01 | 0.89 ± 0.01 |
| EW2.5 | 3.04 ± 0.01 | 2.13 ± 0.19 | 53.57 ± 2.30 | 0.10 ± 0.01 | 0.90 ± 0.00 |
| W10 | 2.98 ± 0.06 | 2.54 ± 0.19 | 41.23 ± 1.95 | 0.19 ± 0.00 | na |
| B10 | 3.08 ± 0.07 | 3.42 ± 0.14 | 75.66 ± 4.83 | 0.28 ± 0.01 | 0.90 ± 0.00 |
| BE7.5 | 2.83 ± 0.00 | 3.34 ± 0.02 | 65.34 ± 8.12 | 0.20 ± 0.15 | 0.89 ± 0.00 |
| BE5 | na | 3.96 ± 0.06 | 50.74 ± 6.33 | 0.25 ± 0.01 | 0.88 ± 0.00 |
| BE2.5 | na | 3.99 ± 0.08 | 52.77 ± 3.85 | 0.23 ± 0.01 | 0.88 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Đorđevski, N.; Uba, A.I.; Zengin, G.; Božunović, J.; Gašić, U.; Ristanović, E.; Ćirić, A.; Nikolić, B.; Stojković, D. Chemical and Biological Investigations of Allium scorodoprasum L. Flower Extracts. Pharmaceuticals 2023, 16, 21. https://doi.org/10.3390/ph16010021
Đorđevski N, Uba AI, Zengin G, Božunović J, Gašić U, Ristanović E, Ćirić A, Nikolić B, Stojković D. Chemical and Biological Investigations of Allium scorodoprasum L. Flower Extracts. Pharmaceuticals. 2023; 16(1):21. https://doi.org/10.3390/ph16010021
Chicago/Turabian StyleĐorđevski, Nikoleta, Abdullahi Ibrahim Uba, Gokhan Zengin, Jelena Božunović, Uroš Gašić, Elizabeta Ristanović, Ana Ćirić, Biljana Nikolić, and Dejan Stojković. 2023. "Chemical and Biological Investigations of Allium scorodoprasum L. Flower Extracts" Pharmaceuticals 16, no. 1: 21. https://doi.org/10.3390/ph16010021