
In Vitro Efficacy of Extracts and Isolated Bioactive Compounds from Ascomycota Fungi in the Treatment of Colorectal Cancer: A Systematic Review
 , and
, and
Abstract
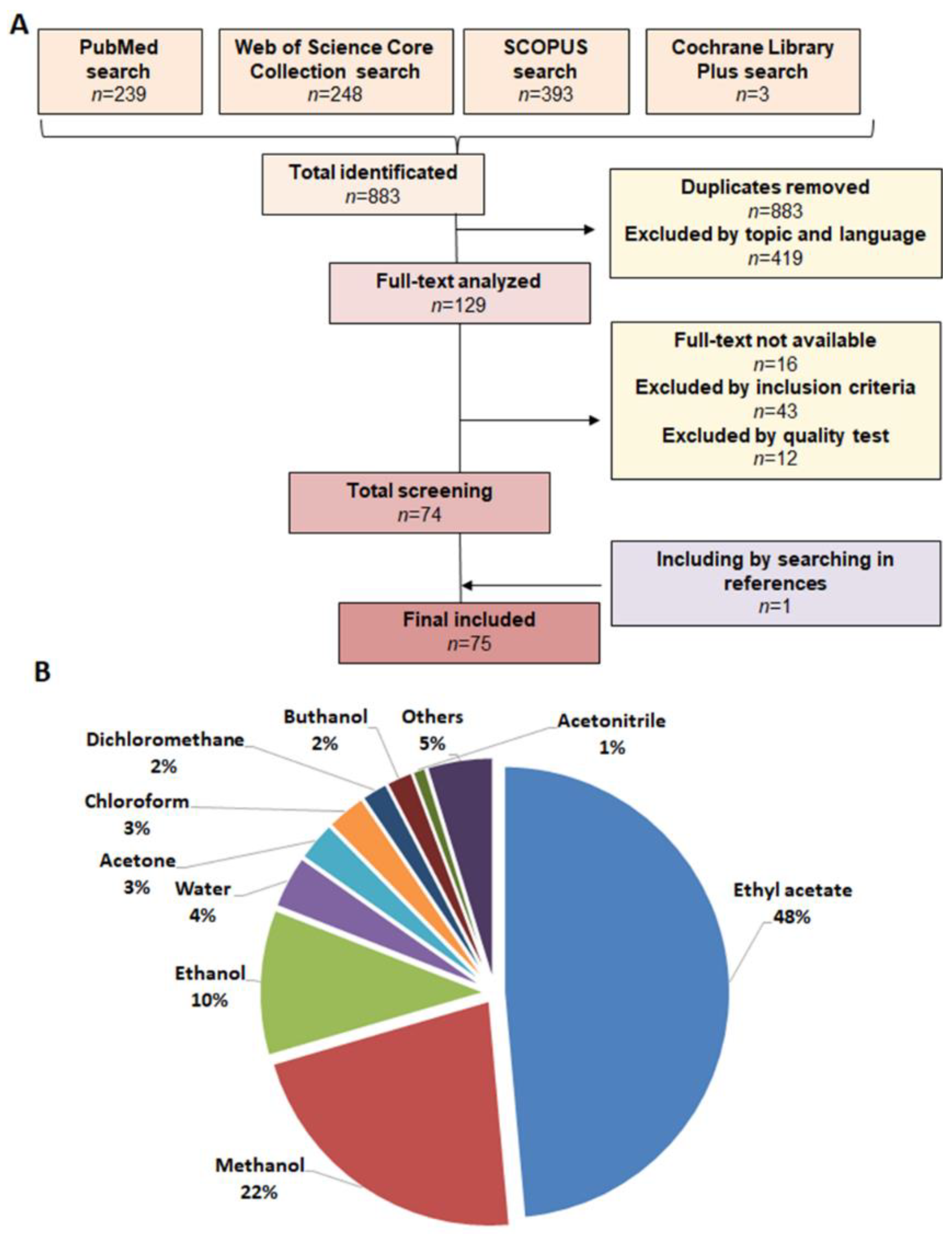
:1. Introduction
2. Results and Discussion
2.1. Order Eurotiales
2.1.1. Genus Aspergillus
2.1.2. Genus Penicillium
2.1.3. Genera Neosartorya, Byssochlamys and Talaromyces
2.2. Order Hypocreales
2.2.1. Genera Cordyceps, Fusarium and Trichoderma
2.2.2. Other Genera
2.3. Orders Pleosporales and Sordariales
2.4. Minoritary Orders

{kind=link}
{kind=link}
| Order | Genus | Isolated from | Extraction | Isolated Compounds | Cell Line/Administration/Cytotoxicity Assay | Compound and IC50 or Cell Death (%) | Mechanism of Action | Reference |
|---|---|---|---|---|---|---|---|---|
| Capnodiales | Zasmidium | Foliage | Ethyl acetate (EtOAc) | 8,8′-Bijuglone | HCT-116 DMSO MTT | 8,8′-Bijuglone: 45 μg/mL | - | [87] |
| Capnodiales Incertae sedis | Cladosporium Hansfordia | C. procera Vernonia amygdalina | EtOAc | - | HT-29 and HCT-116 - MTT | C. cladosporioides extracts 1 (HT-29: 77.7 μg/mL; HCT-116: 45.6 μg/mL), 2 (HT-29 and HCT-116: >100 μg/mL); H. sinuosae extract: HT-29 (47.6 μg/mL), HCT-116 (>100 μg/mL) | - | [39] |
| Capnodiales | Cladosporium | Medicinal plants | Sodium bicarbonate | Taxol | HCT-15 Methanol MTT | Taxol: 3.5 μM | - | [86] |
| Helotiales | Sclerotinia | Contaminated soybean seed | Ethanol | - | HCT-8 - MTT | Fractions ethyl acetate (48.03 μg/mL), F3 (250.50 μg/mL) | - | [91] |
| Helotiales | Sclerotinia | Soybeans | Trituration and boiled | - | CCD-18Co and HT-29 Alone Electronic counter | Aqueous extract: CCD-18Co (11%), HT-29 (58%) | Aqueous extract induces ROS and extrinsic pathway | [90] |
| Helotiales | Lachnum | - | Ethanol | Exopolysaccharide LEP-2b | CT-26 DMSO MTT | LEP-2b: 8816.27 μg/mL; PLEP-2b: 85.78 μg/mL; SLEP_2b: 154.52 μg/mL | - | [89] |
| Diaporthales | Phomopsis | Acanthus ilicifolius | Methanol | Phomolactonexanthone A (1), B (2) and C (3) Dicerandrol A (4), B (5) and C (6) Deacetylphomoxanthone B (7) Penexanthone A (8) | HCT-116 Alone MTT | (1) and (2): >50 μM; (3): 44.06 μM; (4): 2.64 μM; (5): 3.94 μM; (6): 42.63 μM; (7): 7.12 μM; (8): 6.92 μM | - | [92] |
| Diaporthales | Phomopsis | Miquelia dentata | Methanol EtOAc | - | SW-480 and HCT-116 - Hoechst 33342 | Methanol extract: HCT-116 (24.47 μg/mL), SW-480 (14.45 μg/mL) Ethyl acetate extract: HCT-116 (5.63 μg/mL), SW-480 (23.5 μg/mL) | - | [77] |
| Pezizales | Morchella | - | Hot water | Polysaccharides | CaCo-2 Alone Methylene Blue Assay | PMEP: 1.840 mg/mL; Ac-PMEP 1 (2.094 mg/mL), 2 (2.827 mg/mL), 3 (1.229 mg/mL) | - | [94] |
| Pezizales | Morchella | - | Pulsed electric field | - | HT-29 Alone MTT | M2 fraction: 54.29% | M2 fraction induces apoptosis | [93] |
| Xylariales | Xylaria | Aegle marmelos | EtOAc | 5-methylmellein | HCT-116 Alone SRB | 5-methylmellein: 2.0 μg/mL; 5-methylmellein nanoparticle: <0.5 μg/mL | 5-methylmellein nanoparticle induces apoptosis, ROS and mitochondrial membrane potential loss | [95] |
| Xylariales | Daldinia | Tenodera aridifolia | EtOAc | Daldinone F (1) Nodulisporin G (2) Dalmanol C (3) | SW-480 and HCT-116 Alone MTT | (1): SW-480 (9.59 μM), HCT-116 (>20 μM); (2) and (3): SW-480 and HCT-116 (>20 μM) | - | [96] |
| Boliniales | Camarops | Alibertia macrophylla | EtOAc | Xylarenone C and D | HCT-8 Alone MTT | Xylarenones C (1.9 μg/mL), D (1.5 μg/mL) | Xylarenone D shows weak AChE inhibitory activity | [34] |
| Leotiales | Halenospora | Wood | Greensporone A (1), C (2) Dechlorogreensporone A (3), D (4) O-Desmethylgreensporone C (5) | HT-29 DMSO CellTiter 96 Aqueous One Solution Cell Proliferation Assay | (1) and (3): >20 μM; (2): 7.5 μM; (4): 25.4 μM; (5): 13.8 μM | - | [97] | |
| Venturiales | Ochroconis | - | EtOAc | - | HCT-116 DMSO SRB | Ochroconis sp. extract: 70.5 μg/mL | - | [49] |
3. Materials and Methods
3.1. Study Eligibility
3.2. Inclusion Criteria
3.3. Exclusion Criteria
3.4. Data Sources
3.5. Study Selection
3.6. Data Extraction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CRC | Colorectal cancer |
| EGFR | Epidermal growth factor receptor |
| VEGF | Vascular endothelial growth factor |
| DMSO | Dimethyl sulfoxide |
| MTT | 3-(4,5-dimethytlthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| Asp. A | Asperphenins A |
| Asp. B | Asperphenins B |
| SRB | Sulforhodamine B |
| ROS | Reactive oxygen species |
| TXL | Paclitaxel |
| EtOAc | Ethyl acetate |
| CH2Cl2 extract | Sequential ethyl acetate, methanol, and dichloromethane extract |
| Dox | Doxorubicin |
| CCK8 | Cell counting kit-8 |
| WST-1 | 4-[3-(4-Iodo-phenyl)-2-(4-nitrophenyl)-2H-5 tetrazolio]-1,3-benzene disulphonate |
| RTCA-DP | Real-Time Cell Analysis System |
| EAE | Ethyl acetate extract |
| MTS | ((3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium)) assay |
| Mrna | Messenger RNA |
| IC50 | Half maximal inhibitory concentration |
| GSC | Cordyceps militaris cultivated on germinated soybeans |
| EE | Ethanol extract |
| HCT-116/HT-29 oxa | Oxaliplatin resistant cell |
| HCT-116/HT-29 5FU | 5-fluorouracil resistant cell |
| HCT-116/HT-29 SN-38 | SN-38 resistant cell |
| CK01 | Pleosporales sp. Extract |
| SN-38 | 7-Ethyl-10-hydroxycamptothecin |
| AMPK | AMP-activated protein kinase |
| PARP | Poly (ADP-ribose) polymerase |
| PMEP | Polysaccharides extracted from Morchella angusticepes Peck |
| Ac-PMEP | Acetylated derivatives of PMEP |
| AChE | Acetylcholinesterase |
| PLEP | Phosphorylated polysaccharide |
| SLEP | Sulfated polysaccharide |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Recio-Boiles, A.; Cagir, B. Colon Cancer. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Hervieu, C.; Christou, N.; Battu, S.; Mathonnet, M. The Role of Cancer Stem Cells in Colorectal Cancer: From the Basics to Novel Clinical Trials. Cancers 2021, 13, 1092. [Google Scholar] [CrossRef] [PubMed]
- Sang, R.; Stratton, B.; Engel, A.; Deng, W. Liposome technologies towards colorectal cancer therapeutics. Acta Biomater. 2021, 127, 24–40. [Google Scholar] [CrossRef]
- Redondo-Blanco, S.; Fernández, J.; Gutiérrez-del-Río, I.; Villar, C.J.; Lombó, F. New Insights toward Colorectal Cancer Chemotherapy Using Natural Bioactive Compounds. Front. Pharm. 2017, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, Y.; Zhang, L.; Tian, Q. Mushroom polysaccharide lentinan for treating different types of cancers: A review of 12 years clinical studies in China. Prog. Mol. Biol. Transl. Sci. 2019, 163, 297–328. [Google Scholar]
- Mesas, C.; Fuel, M.; Martínez, R.; Prados, J.; Melguizo, C.; Porres, J.M. In vitro evidence of the antitumor capacity of Solanaceae and Cucurbitaceae in colon cancer: A systematic review. Crit. Rev. Food Sci. Nutr. 2021, 62, 1–22. [Google Scholar]
- Dai, R.; Liu, M.; Nik Nabil, W.N.; Xi, Z.; Xu, H. Mycomedicine: A Unique Class of Natural Products with Potent Anti-tumour Bioactivities. Molecules 2021, 26, 1113. [Google Scholar] [CrossRef]
- Contreras-Ochoa, C.O.; Maza-Lopez, J.; Mendoza de Gives, P.; Aguilar-Marcelino, L.; Mojica-Cardoso, C.; Dimas-González, J.; Fernández-Coto, D.L.; Reyna-Figueroa, J.; López-Arellano, M.E.; Lagunas-Martínez, A. Cell death induction by mycelium extracts from Pleurotus spp. on cervical cancer cell lines. Nat. Prod. Res. 2022, 36, 6091–6609. [Google Scholar] [CrossRef]
- El-Bondkly, E.A.M.; El-Bondkly, A.A.M.; El-Bondkly, A.A.M. Marine endophytic fungal metabolites: A whole new world of pharmaceutical therapy exploration. Heliyon 2021, 7, e06362. [Google Scholar] [CrossRef] [PubMed]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, J.K.; Maharachchikumbura, S.S.N.; Ekanayaka, A.H.; Tian, Q.; Phookamsak, R. Outline of Ascomycota: 2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Rupa, E.J.; Li, J.F.; Arif, M.H.; Yaxi, H.; Puja, A.M.; Chan, A.J.; Hoang, V.; Kaliraj, L.; Yang, D.C.; Kang, S.C. Cordyceps militaris Fungus Extracts-Mediated Nanoemulsion for Improvement Antioxidant, Antimicrobial, and Anti-Inflammatory Activities. Molecules 2020, 25, 5733. [Google Scholar] [CrossRef] [PubMed]
- Ukwatta, K.M.; Lawrence, J.L.; Wijayarathne, C.D. Antimicrobial, anti-cancer, anti-filarial and anti-inflammatory activities of Cowabenzophenone A extracted from the endophytic fungus Aspergillus terreus isolated from a mangrove plant Bruguiera gymnorrhyza. Mycology 2020, 11, 297–305. [Google Scholar] [CrossRef] [Green Version]
- El-Gendy, M.M.A.A.; Awad, M.F.; El-Shenawy, F.S.; El-Bondkly, A.M.A. Production, purification, characterization, antioxidant and antiproliferative activities of extracellular L-asparaginase produced by Fusarium equiseti AHMF4. Saudi J. Biol. Sci. 2021, 28, 2540–2548. [Google Scholar] [CrossRef]
- Opattova, A.; Horak, J.; Vodenkova, S.; Kostovcikova, K.; Cumova, A.; Macinga, P.; Galanova, N.; Rejhova, A.; Vodickova, L.; Kozics, K.; et al. Ganoderma Lucidum induces oxidative DNA damage and enhances the effect of 5-Fluorouracil in colorectal cancer in vitro and in vivo. Mutat. Res. Gen Tox. Environ. 2019, 845, 403065. [Google Scholar] [CrossRef]
- Qi, W.; Zhou, X.; Wang, J.; Zhang, K.; Zhou, Y.; Chen, S.; Nie, S.; Xie, M. Cordyceps sinensis polysaccharide inhibits colon cancer cells growth by inducing apoptosis and autophagy flux blockage via mTOR signaling. Carbohydr. Polym. 2020, 237, 116113. [Google Scholar] [CrossRef]
- Abd El-Hady, F.K.; Shaker, K.H.; Souleman, A.M.A.; Fayad, W.; Abdel-Aziz, M.S.; Hamed, A.A.; Iodice, C.; Tommonaro, G. Comparative Correlation Between Chemical Composition and Cytotoxic Potential of the Coral-Associated Fungus Aspergillus sp. 2C1-EGY Against Human Colon Cancer Cells. Curr. Microbiol. 2017, 74, 1294–1300. [Google Scholar] [CrossRef]
- Bae, S.Y.; Liao, L.; Park, S.H.; Kim, W.K.; Shin, J.; Lee, S.K. Antitumor Activity of Asperphenin, A., a Lipopeptidyl Benzophenone from Marine-Derived Aspergillus sp. Fungus, by Inhibiting Tubulin Polymerization in Colon Cancer Cells. Mar. Drugs 2020, 18, 110. [Google Scholar] [CrossRef] [Green Version]
- Perlatti, B.; Lan, N.; Earp, C.E.; AghaAmiri, S.; Vargas, S.H.; Azhdarinia, A.; Bills, G.F.; Gloer, J.B. Arenicolins: C-Glycosylated Depsides from Penicillium arenicola. J. Nat. Prod. 2020, 83, 668–674. [Google Scholar] [CrossRef]
- Shoeb, M.; Thoo-Lin, P.K.; Nahar, N. Anti-colon cancer activity of endophytic fungal strains from Terminalia chebula Rezt. Bangladesh. J. Pharm. 2012, 7, 47–49. [Google Scholar] [CrossRef] [Green Version]
- Alasmary, F.A.S.; Awaad, A.S.; Kamal, M.; Alqasoumi, S.I.; Zain, M.E. Antitumor activity of extract and isolated compounds from Drechslera rostrata and Eurotium tonophilum. Saudi Pharm. J. 2018, 26, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.; Abdelwahab, M. Antimicrobial and Cytotoxic Activities of Plant-Derived Endophytic Metabolites from Northern Border Region, KSA. Indo Am. 2018, 5, 1122–1132. [Google Scholar]
- Mohamed, H.F. Molecular analysis and anticancer properties of two identified isolates, Fusarium solani and Emericella nidulans isolated from Wady El-Natron soil in Egypt against Caco-2 (ATCC) cell line. Asian Pac. J. Trop. Biomed. 2012, 2, 863–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montenegro, T.G.C.; Rodrigues, F.A.R.; Jimenez, P.C.; Angelim, A.L.; Melo, V.M.M.; Rodrigues Filho, E.; de Oliveira, M.; Costa-Lotufo, L. Cytotoxic Activity of Fungal Strains Isolated from the Ascidian Eudistoma vannamei. Chem. Biodivers. 2012, 9, 2203–2209. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, H.; Wu, N.; Liu, M.; Wei, J.; Zhang, Y.; Lin, X. Characterization of the high cytochalasin E and rosellichalasin producing-Aspergillus sp. nov. F1 isolated from marine solar saltern in China. World J. Microbiol. Biotechnol. 2013, 29, 11–17. [Google Scholar] [CrossRef]
- Ye, P.; Shen, L.; Jiang, W.; Ye, Y.; Chen, C.T.A.; Wu, X.; Wu, B. Zn-Driven Discovery of a Hydrothermal Vent Fungal Metabolite Clavatustide, C, and an Experimental Study of the Anti-Cancer Mechanism of Clavatustide B. Mar. Drugs 2014, 12, 3203–3217. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhang, X.; Nong, X.; Wang, J.; Qi, S. Brevianamides and Mycophenolic Acid Derivatives from the Deep-Sea-Derived Fungus Penicillium brevicompactum DFFSCS025. Mar. Drugs 2017, 15, 43. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Feng, Y.; Kramer, M.; Essmann, F.; Grond, S. A New Acetylenic Compound and Other Bioactive Metabolites from a Shark Gill-derived Penicillium Strain. Rec. Nat. Prod. 2017, 11, 31–36. [Google Scholar]
- Phainuphong, P.; Rukachaisirikul, V.; Phongpaichit, S.; Sakayaroj, J.; Kanjanasirirat, P.; Borwornpinyo, S.; Akrimajirachoote, N.; Yimnual, C.; Muanprasat, C. Depsides and depsidones from the soil-derived fungus Aspergillus unguis PSU-RSPG204. Tetrahedron 2018, 74, 5691–5699. [Google Scholar] [CrossRef]
- Asfour, H.Z.; Awan, Z.A.; Bagalagel, A.A.; Elfaky, M.A.; Abdelhameed, R.F.A.; Elhady, S.S. Large-Scale Production of Bioactive Terrein by Aspergillus terreus Strain S020 Isolated from the Saudi Coast of the Red Sea. Biomolecules 2019, 9, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artasasta, M.; Yanwirasti, T.M.; Djamaan, A.; Handayani, D. Cytotoxic and antibacterial activities of marine sponge-derived fungus Aspergillus nomius NC06. Rasayan. J. Chem. 2019, 12, 1463–1469. [Google Scholar] [CrossRef]
- Mandelare, P.E.; Adpressa, D.A.; Kaweesa, E.N.; Zakharov, L.N.; Loesgen, S. Coculture of Two Developmental Stages of a Marine-Derived Aspergillus alliaceus Results in the Production of the Cytotoxic Bianthrone Allianthrone A. J. Nat. Prod. 2018, 81, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zhang, C.; Huang, J.; Zhang, J.; Liu, D.; Huang, J.; Proksch, P.; Lin, W. Violaceimides A–E, sulfur-containing metabolites from a sponge-associated fungus Aspergillus violaceus. Tetrahedron. Lett. 2018, 59, 3157–3160. [Google Scholar] [CrossRef]
- Ali, M.; AbdElfattah, A.; Khalil, N.; Sayed, M. Biological Activities of the Alkaloid Quinazoline Extracted from Aspergillus nomius. Egypt J. Bot. 2017, 57, 565–582. [Google Scholar] [CrossRef]
- Artasasta, M.A.; Yanwirasti, D.A.; Handayani, S. Cytotoxic activity screening of ethyl acetate fungal extracts derived from the marine sponge Neopetrosia chaliniformis AR-01. J. Appl. Pharm. Sci. 2017, 7, 174–178. [Google Scholar]
- Dikmen, M.; Canturk, Z.; Artagan, O.; Ozturk, N. Antioxidant, Antiproliferative and Apoptotic Effects of Secondary Metabolites of Halotolerant Aspergillus terreus on Colon Adenocarcinoma Caco-2 Cells. Int. J. Pharm. 2017, 13, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Khiralla, A.; Mohamed, I.E.; Tzanova, T.; Schohn, H.; Slezack-Deschaumes, S.; Hehn, A.; André, P.; Carre, G.; Spina, R.; Lobstein, A.; et al. Endophytic fungi associated with Sudanese medicinal plants show cytotoxic and antibiotic potential. FEMS Microbiol. Lett. 2016, 363, fnw089. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Z.; Lam, W.; Gullen, E.A.; Yu, Z.; Wei, Y.; Wang, L.; Zeiss, C.; Beck, A.; Cheng, E.; et al. Study of malformin C, a Fungal Source Cyclic Pentapeptide, as an Anti-Cancer Drug. PLoS ONE 2015, 10, e0140069. [Google Scholar] [CrossRef]
- Gu, W.; Qiao, C. Furandiones from an endophytic Aspergillus terreus residing in Malus halliana. Chem. Pharm. Bull. 2012, 60, 1474–1477. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.J.; Park, J.S.; Kim, Y.J.; Jung, J.H.; Lee, J.K.; Kwon, H.C.; Yang, H.O. Apoptosis-inducing effect of diketopiperazine disulfides produced by Aspergillus sp. KMD 901 isolated from marine sediment on HCT116 colon cancer cell lines. J. Appl. Microbiol. 2011, 110, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Xu, M.Y.; Li, H.J.; Zeng, K.J.; Ma, W.Z.; Tian, G.B.; Xu, J.; Yang, D.; Lan, W. Diverse Secondary Metabolites from the Marine-Derived Fungus Dichotomomyces cejpii F31-1. Mar. Drugs 2017, 15, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, B.S.F.; Sahm, B.D.B.; Jimenez, P.C.; Pinto, F.C.L.; Mafezoli, J.; Mattos, M.C.; Rodrigues-Filho, E.; Pfenning, L.H.; Abreu, L.M.; Costa-Lotufo, L.; et al. Bioprospection of Cytotoxic Compounds in Fungal Strains Recovered from Sediments of the Brazilian Coast. Chem. Biodivers. 2015, 12, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Castro-Carvalho, B.; Ramos, A.A.; Prata-Sena, M.; Malhão, F.; Moreira, M.; Gargiulo, D.; Dethoup, T.; Buttachon, S.; Kijjoa, A.; Rocha, E. Marine-derived Fungi Extracts Enhance the Cytotoxic Activity of Doxorubicin in Nonsmall Cell Lung Cancer Cells A459. Pharm. Res. 2017, 9, S92–S928. [Google Scholar]
- Prata-Sena, M.; Ramos, A.A.; Buttachon, S.; Castro-Carvalho, B.; Marques, P.; Dethoup, T.; Kijjoa, A.; Rocha, E. Cytotoxic activity of Secondary Metabolites from Marine-derived Fungus Neosartorya siamensis in Human Cancer Cells. Phytother. Res. 2016, 30, 1862–1871. [Google Scholar] [CrossRef]
- Ramos, A.A.; Castro-Carvalho, B.; Prata-Sena, M.; Dethoup, T.; Buttachon, S.; Kijjoa, A.; Rocha, E. Crude Extracts of Marine-derived and Soil Fungi of the Genus Neosartorya Exhibit Selective Anticancer Activity by Inducing Cell Death in Colon, Breast and Skin Cancer Cell Lines. Pharm. Res. 2016, 8, 8–15. [Google Scholar]
- Liang, W.L.; Le, X.; Li, H.J.; Yang, X.L.; Chen, J.X.; Xu, J.; Liu, H.; Wang, L.; Wang, K.; Hu, K.; et al. Exploring the Chemodiversity and Biological Activities of the Secondary Metabolites from the Marine Fungus Neosartorya pseudofischeri. Mar. Drugs 2014, 12, 5657–5676. [Google Scholar] [CrossRef]
- Lim, S.M.; Agatonovic-Kustrin, S.; Lim, F.T.; Ramasamy, K. High-performance thin layer chromatography-based phytochemical and bioactivity characterisation of anticancer endophytic fungal extracts derived from marine plants. J. Pharm. Biomed. Anal. 2021, 193, 113702. [Google Scholar] [CrossRef]
- Chen, L.; Fan, D.M.; Tang, J.W.; An, T.; Li, X.; Kong, L.M.; Puno, P.; Li, Y. Discovery of isopenicin A, a meroterpenoid as a novel inhibitor of tubulin polymerization. Biochem. Biophys. Res. Commun. 2020, 525, 303–307. [Google Scholar] [CrossRef]
- Vinale, F.; Salvatore, M.M.; Nicoletti, R.; Staropoli, A.; Manganiello, G.; Venneri, T.; Borrelli, F.; DellaGreca, M.; Salvatore, F.; Andolfi, A. Identification of the Main Metabolites of a Marine-Derived Strain of Penicillium brevicompactum Using LC and GC MS Techniques. Metabolites 2020, 10, 55. [Google Scholar] [CrossRef] [Green Version]
- Abou Zied, A.M.; Abd El-Zaher, E.H.; Elsharquawy, H.A.; Hammad, T. Exopolysaccharides Production and Characterization from Marine-Derived Penicillium commune KP942881.1 with Some Medical Potential Applications. Egypt. J. Bot. 2017, 57, 17–30. [Google Scholar] [CrossRef]
- Ali, T.; Inagaki, M.; Chai, H.; Wieboldt, T.; Rapplye, C.; Rakotondraibe, L.H. Halogenated Compounds from Directed Fermentation of Penicillium concentricum, an Endophytic Fungus of the Liverwort Trichocolea tomentella. J. Nat. Prod. 2017, 80, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Dikmen, M.; Canturk, Z.; Tilki, E.K.; Engur, S. Evaluation of Antiangiogenic and Antimetastatic Effects of Penicillium chrysogenum Secondary Metabolites. Indian J. Pharm. Sci. 2017, 79, 49–57. [Google Scholar] [CrossRef]
- Liu, Z.G.; Bao, L.; Liu, H.W.; Ren, J.W.; Wang, W.Z.; Wang, L.; Li, W.; Yin, W. Chemical diversity from the Tibetan Plateau fungi Penicillium kongii and P. brasilianum. Mycology 2018, 9, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canturk, Z.; Artagan, Ö.; Dıkmen, M. Anticancer effects of secondary metabolites of Penicillium chrysogenum var. chrysogenum on colon adenocarcinoma cells. Fresenius Environ. Bull. 2016, 25, 6190–6197. [Google Scholar]
- Li, C.S.; Li, X.M.; Gao, S.S.; Lu, Y.H.; Wang, B.G. Cytotoxic Anthranilic Acid Derivatives from Deep Sea Sediment-Derived Fungus Penicillium paneum SD-44. Mar. Drugs 2013, 11, 3068–3076. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, G.C.; Lakshmanan, M.D.; Bp, A.; Rs, H.K.; Th, A.K.; Ananthaswamy, K.; C, J. Extract of Penicillium sclerotiorum an endophytic fungus isolated from Cassia fistula L. induces cell cycle arrest leading to apoptosis through mitochondrial membrane depolarization in human cervical cancer cells. Biomed. Pharm. 2018, 105, 1062–1071. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Lee, J.S.; Qian, Z.J.; Li, Y.X.; Kim, K.N.; Heo, S.J.; Heo, S.J.; Jeon, Y.J.; Park, W.S.; Choi, I.W.; et al. Gliotoxin Isolated from Marine Fungus Aspergillus sp. Induces Apoptosis of Human Cervical Cancer and Chondrosarcoma Cells. Mar. Drugs 2013, 12, 69–87. [Google Scholar] [CrossRef]
- Tang, W.; Liu, Z.L.; Mai, X.Y.; Qi, X.; Li, D.H.; Gu, Q.Q. Identification of Gliotoxin isolated from marine fungus as a new pyruvate kinase M2 inhibitor. Biochem. Biophys. Res. Commun. 2020, 528, 594–600. [Google Scholar] [CrossRef]
- Chen, J.; Lou, Q.; He, L.; Wen, C.; Lin, M.; Zhu, Z.; Wang, F.; Huang, L.; Lan, W.; Iwamoto, A.; et al. Reduced-gliotoxin induces ROS-mediated anoikis in human colorectal cancer cells. Int. J. Oncol. 2018, 52, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Reis, F.S.; Barros, L.; Calhelha, R.C.; Cirić, A.; van Griensven, L.J.L.D.; Soković, M.; Ferreira, I. The methanolic extract of Cordyceps militaris (L.) Link fruiting body shows antioxidant, antibacterial, antifungal and antihuman tumor cell lines properties. Food Chem. Toxicol. 2013, 62, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šeklić, D.S.; Stanković, M.S.; Milutinović, M.G.; Topuzović, M.D.; Štajn, A.Š.; Marković, S.D. Cytotoxic, antimigratory, pro-and antioxidative activities of extracts from medicinal mushrooms on colon cancer cell lines. Arch. Biol. Sci. 2016, 68, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.H.; Lee, S.; Lee, K.; Shin, Y.S.; Kang, H.; Cho, H. Anti-cancer effect of Cordyceps militaris in human colorectal carcinoma RKO cells via cell cycle arrest and mitochondrial apoptosis. DARU J. Pharm. Sci. 2015, 23, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mollah, M.L.; Park, D.K.; Park, H.J. Cordyceps militaris Grown on Germinated Soybean Induces G2/M Cell Cycle Arrest through Downregulation of Cyclin B1 and Cdc25c in Human Colon Cancer HT-29 Cells. Evid. Based Complement. Alternat. Med. 2012, 2012, e249217. [Google Scholar] [CrossRef] [Green Version]
- Katoch, M.; Singh-Ranger, G.; Sharma, S.; Gupta, N.; Sangwan, P.L.; Saxena, A.K. Cytotoxic and antimicrobial activities of endophytic fungi isolated from Bacopa monnieri (L.) Pennell (Scrophulariaceae). BMC Complement. Altern. Med. 2014, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.E.; Sonbol, H.; Alwakeel, S.S.; Alotaibi, M.O.; Alotaibi, S.; Alothman, N.; Suliman, R.S.; Ahmedah, H.T.; Ali, R. Investigation of biological activity of soil fungal extracts and LC/MS-QTOF based metabolite profiling. Sci. Rep. 2021, 11, 4760. [Google Scholar] [CrossRef]
- Venugopalan, A.; Potunuru, U.R.; Dixit, M.; Srivastava, S. Reprint of: Effect of fermentation parameters, elicitors and precursors on camptothecin production from the endophyte Fusarium solani. Bioresour. Technol. 2016, 213, 311–318. [Google Scholar] [CrossRef]
- Riera, P.; Páez, D. Elucidating the role of pharmacogenetics in irinotecan efficacy and adverse events in metastatic colorectal cancer patients. Expert. Opin. Drug Metab. Toxicol. 2021, 17, 1157–1163. [Google Scholar] [CrossRef]
- Setyowati, E.P.; Pratiwi, S.U.T.; Purwantini, I. In-vitro cytotoxicity and apoptosis mechanism of ethyl acetate extract from Trichoderma reesei strain TV221 associated with marine sponge: Stylissa flabelliformis. J. Appl. Pharm. Sci. 2018, 8, 151–157. [Google Scholar]
- Cui, J.; Shang, R.Y.; Sun, M.; Li, Y.X.; Liu, H.Y.; Lin, H.W.; Jiao, W.H. Trichodermaloids A–C, Cadinane Sesquiterpenes from a Marine Sponge Symbiotic Trichoderma sp. SM16 Fungus. Chem. Biodivers. 2020, 17, e2000036. [Google Scholar] [CrossRef]
- Yamazaki, H.; Rotinsulu, H.; Kaneko, T.; Murakami, K.; Fujiwara, H.; Ukai, K.; Namikoshi, M. A New Dibenz[b,e]oxepine Derivative, 1-Hydroxy-10-methoxy-dibenz[b,e]oxepin-6,11-dione, from a Marine-Derived Fungus, Beauveria bassiana TPU942. Mar. Drugs 2012, 10, 2691–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Guo, S.; Zhu, H. Statistical optimization of culture medium for production of exopolysaccharide from endophytic fungus Bionectria ochroleuca and its antitumor effect in vitro. EXCLI J. 2016, 15, 211–220. [Google Scholar] [PubMed]
- Dornetshuber-Fleiss, R.; Heffeter, P.; Mohr, T.; Hazemi, P.; Kryeziu, K.; Seger, C.; Berger, W.; Lemmens-Gruber, R. Destruxins: Fungal-derived cyclohexadepsipeptides with multifaceted anticancer and antiangiogenic activities. Biochem. Pharm. 2013, 86, 361–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Khawaga, M.A.; El-Aziz, M.M.A.; Hegazi, G. Identification and bioactive potential of endophytic fungi isolated from Calotropis procera (Ait.) R. Br. Life Sci. J. 2013, 10, 2804–2814. [Google Scholar]
- Heilos, D.; Rodríguez-Carrasco, Y.; Englinger, B.; Timelthaler, G.; Van Schoonhoven, S.; Sulyok, M.; Boecker, S.; Süssmuth, R.; Heffeter, P.; Lemmens-Gruber, R.; et al. The Natural Fungal Metabolite Beauvericin Exerts Anticancer Activity In Vivo: A Pre-Clinical Pilot Study. Toxins 2017, 9, 258. [Google Scholar] [CrossRef] [Green Version]
- Shweta, S.; Gurumurthy, B.R.; Ravikanth, G.; Ramanan, U.S.; Shivanna, M.B. Endophytic fungi from Miquelia dentata Bedd., produce the anti-cancer alkaloid, camptothecine. Phytomedicine 2013, 20, 337–342. [Google Scholar] [CrossRef]
- Fang, Z.F.; Yu, S.S.; Zhou, W.Q.; Chen, X.G.; Ma, S.G.; Li, Y.; Qu, J. A new isocoumarin from metabolites of the endophytic fungus Alternaria tenuissima (Nees & T. Nees: Fr.) Wiltshire. Chin. Chem. Lett. 2012, 23, 317–320. [Google Scholar]
- El-Elimat, T.; Figueroa, M.; Raja, H.A.; Graf, T.N.; Swanson, S.M.; Falkinham, J.O.; Wani, M.C.; Pearce, C.J.; Oberlies, N.H. Biosynthetically Distinct Cytotoxic Polyketides from Setophoma terrestris. Eur. J. Org. Chem. 2015, 2015, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Dezaire, A.; Marchand, C.H.; Vallet, M.; Ferrand, N.; Chaouch, S.; Mouray, E.; Larsen, A.K.; Sabbah, M.; Lemaire, S.D.; Prado, S.; et al. Secondary Metabolites from the Culture of the Marine-derived Fungus Paradendryphiella salina PC 362H and Evaluation of the Anticancer Activity of Its Metabolite Hyalodendrin. Mar. Drugs 2020, 18, 191. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xiao, J.; Gao, Y.Q.; Tang, J.; Zhang, A.L.; Gao, J.M. Chaetoglobosins from Chaetomium globosum, an Endophytic Fungus in Ginkgo biloba, and Their Phytotoxic and Cytotoxic Activities. J. Agric. Food Chem. 2014, 62, 3734–3741. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, J.; Hu, S.; Ma, H.; Zhu, H.; Tong, Q.; Cheng, L.; Hao, X.; Zhang, G.; Zhang, Y. Secondary metabolites from endophytic fungus Chaetomium sp. induce colon cancer cell apoptotic death. Fitoterapia 2017, 121, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yin, J.; Yan, S.; Hu, P.; Huang, J.; Zhang, G.; Wang, F.; Tong, Q.; Zhang, Y. Chaetocochin J, an epipolythiodioxopiperazine alkaloid, induces apoptosis and autophagy in colorectal cancer via AMPK and PI3K/AKT/mTOR pathways. Bioorganic. Chem. 2021, 109, 104693. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Ren, F.; Niu, S.; Liu, X.; Che, Y. Dioxatricyclic and Oxabicyclic Polyketides from Trichocladium opacum. J. Nat. Prod. 2014, 77, 9–14. [Google Scholar] [CrossRef] [PubMed]
- El-Elimat, T.; Raja, H.; Figueroa, M.; Swanson, S.M.; Falkinham, J.O., III; Lucas, D.M.; Grever, M.R.; Wani, M.C.; Pearce, C.J.; Oberlies, N.H. Sorbicillinoid analogs with cytotoxic and selective anti-Aspergillus activities from Scytalidium album. J. Antibiot. 2015, 68, 191–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gokul Raj, K.; Manikandan, R.; Arulvasu, C.; Pandi, M. Anti-proliferative effect of fungal taxol extracted from Cladosporium oxysporum against human pathogenic bacteria and human colon cancer cell line HCT 15. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 138, 667–674. [Google Scholar] [CrossRef] [PubMed]
- González-Montiel, G.A.; Kaweesa, E.N.; Feau, N.; Hamelin, R.C.; Stone, J.K.; Loesgen, S. Chemical, Bioactivity, and Biosynthetic Screening of Epiphytic Fungus Zasmidium pseudotsugae. Molecules 2020, 25, 2358. [Google Scholar] [CrossRef]
- Yang, M.H.; Baek, S.H.; Ha, I.J.; Um, J.Y.; Ahn, K.S. Brassinin enhances the anticancer actions of paclitaxel by targeting multiple signaling pathways in colorectal cancer cells. Phytother. Res. 2021, 35, 3875–3885. [Google Scholar] [CrossRef]
- He, Y.; Ye, M.; Jing, L.; Du, Z.; Surhio, M.M.; Xu, H.; Li, J. Preparation, characterization and bioactivities of derivatives of an exopolysaccharide from Lachnum. Carbohydr. Polym. 2015, 117, 788–796. [Google Scholar] [CrossRef]
- Azevedo, L.; Chagas-Paula, D.A.; Kim, H.; Roque, A.C.M.; Días, K.S.T.; Machado, J.C.; Azevedo, M.G.; Mertens-Talcott, S.U. White mold (Sclerotinia sclerotirum), friend or foe: Cytotoxic and mutagenic activities in vitro and in vivo. Food Res. Int. 2016, 80, 27–35. [Google Scholar] [CrossRef]
- Pressete, C.G.; Giannini, L.S.V.; de Paula, D.A.C.; do Carmo, M.A.V.; Assis, D.M.; Santos, M.F.C.; Machado, J.C.; Marques, M.J.; Soares, M.G.; Azevedo, L. Sclerotinia Sclerotiorum (White Mold): Cytotoxic, Mutagenic, and Antimalarial Effects In Vivo and In Vitro. J. Food Sci. 2019, 84, 3866–3875. [Google Scholar] [CrossRef]
- Ding, B.; Yuan, J.; Huang, X.; Wen, W.; Zhu, X.; Liu, Y.; Li, H.; Lu, Y.; He, L.; Tan, H.; et al. New Dimeric Members of the Phomoxanthone Family: Phomolactonexanthones, A, B and Deacetylphomoxanthone C Isolated from the Fungus Phomopsis sp. Mar. Drugs 2013, 11, 4961–4972. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, Y.; Mao, Q.; Guo, X.; Li, P.; Liu, Y.; Xu, N. Characteristics and Antitumor Activity of Morchella esculenta Polysaccharide Extracted by Pulsed Electric Field. Int. J. Mol. Sci. 2016, 17, 986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Chen, J.; Li, F.; Yang, Y.; Wu, S.; Ming, J. Antioxidant and Antiproliferative Activities of Modified Polysaccharides Originally Isolated from Morchella Angusticepes Peck. J. Food Sci. 2019, 84, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Arora, D.; Kumar, A.; Gupta, P.; Chashoo, G.; Jaglan, S. Preparation, characterization and cytotoxic evaluation of bovine serum albumin nanoparticles encapsulating 5-methylmellein: A secondary metabolite isolated from Xylaria psidii. Bioorg. Med. Chem. Lett. 2017, 27, 5126–5130. [Google Scholar] [CrossRef]
- Wang, G.; Fan, J.Y.; Zhang, W.J.; Hua, C.P.; Chen, C.J.; Yan, W.; Ge, H.M.; Jiao, R.H.; Tan, R.X. Polyketides from Mantis-Associated Fungus Daldinia eschscholzii IFB-TL01. Chem. Biodivers. 2015, 12, 1349–1355. [Google Scholar] [CrossRef]
- Gubiani, J.R.; Nogueira, C.R.; Pereira, M.D.P.; Young, M.C.M.; Ferreira, P.M.P.; de Moraes, M.O.; Pessoa, C.; Bolzani, V.S.; Araujo, A.R. Rearranged Sesquiterpenes and Branched Polyketides Produced by the Endophyte Camarops sp. Phytochem. Lett. 2016, 17, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Száva-Kováts, E. Unfounded attribution of the “half-life” index-number of literature obsolescence to Burton and Kebler: A literature science study. J. Am. Soc. Inf. Sci. 2002, 53, 1098–1105. [Google Scholar] [CrossRef]
- Cohen, J. Weighted kappa: Nominal scale agreement with provision for scaled disagreement or partial credit. Psychol. Bull. 1968, 70, 213–220. [Google Scholar] [CrossRef]


| Genus | Isolated from | Extraction | Isolated Compounds | Cell Line/Administration/Cytotoxicity Assay | Compound and IC50 or Cell Death (%) | Mechanism of Action | Reference |
|---|---|---|---|---|---|---|---|
| Aspergillus | Submerged decaying wood | Methanol/ Dichloromethane | Asperphenins A (Asp. A) Asperphenins B (Asp. B) | RKO Alone and in combination with irinotecan and TXL SRB | Asp. A: 0.84 µM Asp. B: 1.26 µM Asp. A + Irinotecan at 1.25, 2.5, 5 and 10 µM (0.652, 0.811, 0.756, 0.694 and 0.652 μg/mL, respectively) Asp. A + TXL at 0.16, 0.8, 4 and 20µM (1.673, 1.925, 0.742 and 1.185 μg/mL, respectively) | Asp. A inhibits tubulin polymerization, generates ROS and induce G2/M arrest and apoptosis Asp. A and Irinotecan show synergism | [20] |
| Aspergillus | Neopetrosia chaliniformis | Ethyl acetate (EtOAc) | - | HCT-116 - MTT | Fractions I (193.64 μg/mL), II (5.28 μg/mL), III (15.82 μg/mL), IV (10.27 μg/mL), V (45.57 μg/mL) | - | [33] |
| Aspergillus Penicillium | Stylissa carteri Sediment Hyrtios erectus Marine sediment | Liquid medium: EtOAc Mycelia: Methanol | - | HCT-116 DMSO SRB | Penicillium Mycelia (M): between 15.00 and 92.60 μg/mL; Broth (B): between 74.20 and >100 μg/mL Aspergillus M: between 48.00 and 97.00 μg/mL; B: between 42.75 and 90.00 μg/mL | - | [32] |
| Aspergillus | Bruguiera gymnorrhyza | EtOAc | - | HCT-116 Alone MTT | EtOAc extract (EAE): 10.1 μM | - | [15] |
| Aspergillus | Achillea fragrantissima | Ethanol | - | HCT-116 DMSO MTT | Subfractions N. Hexane (76 μg/mL), EtOAc (26.3 μg/mL), Butanol (89.1 μg/mL) | - | [24] |
| Aspergillus | Marine Alga | EtOAc | Allianthrone A-C | HCT-116 DMSO MTT | Allianthrone A, B and C: >20 μM | - | [34] |
| Aspergillus | - | Liquid medium: Methanol Mycelia: EtOAc | Aspergiside B (1) Aspergisidone (2) Emeguisin A (3) Folipastatin (4) Aspergillusidone C (5) Unguinol (6) 2-Chlorounguinol (7) 2,4-Dichlorounguinol (8) Nidulin (9) | HCT-116 - MTT | (1): 3.98%; (2): 20.19%; (3): 23.5 µM; (4): 53.69%; (5): 7.84%; (6): 21.59%; (7): 4.92%; (8): 52.49%; (9): 3.58% | - | [31] |
| Aspergillus | Sponge | EtOAc | Violaceimide A-E | HCT-8 - MTT | Violaceimides A (1.5 μM), B (2.51 μM), C, D (>20 μM), E (>100 μM) | - | [35] |
| Aspergillus | Sinularia sp. | EtOAc Methanol Dichloromethane | - | HCT-116 DMSO MTT | CH2Cl2 extract: 15.8% Fractions 1 (29%), 2 (38%), 2c (88%), 2d (85%) | - | [19] |
| Aspergillus | Soil | EtOAc | - | HCT-116 DMSO SRB | A. niger (69.1%), A. nomius (68.2%), A. terreus (63.7%), A. fumigatus (60.3%), A. flavus (55.7%), A. candidus (55.1%), A. stellifer (51.9%), A. oryzae (50.7%), A. violaceus (50.1%) | - | [36] |
| Aspergillus | Neopetrosia chaliniformis | EtOAc | - | WiDr - MTT | EAE of NC01 (87.89%), NC02 (102.43%), NC03 (70.98%), A. nomius (29.69%), NC07 (85.96%), NC08 (4.48%), NC09 (53.96%) | EAE of NC01 induces apoptosis | [37] |
| Aspergillus | Lake | EtOAc | - | CaCo-2 DMSO WST-1 and RTCA | EAE: 185.9 μg/mL | - | [38] |
| Aspergillus Byssochlamys | Calotropis procera Catharanthus roseus Euphorbia prostrata Vernonia amygdalina | EtOAc | - | HT-29 and HCT-116 - MTT | A. terreus 1: HT-29 and HCT-116: >100 μg/mL A. terreus 2: HT-29: >100 μg/mL, HCT-116: 30.7 μg/mL Byssochlamys: HT-29: 56.3 μg/mL, HCT-116: 30.4 μg/mL | - | [39] |
| Aspergillus | Sand soil | N-butyl alcohol | Malformin C | MC-38 and HCT-116 - Methylene blue | Malformin C: 0.27 and 0.18 µM (MC-38 and HCT-116, respectively) | Malformin C induces G2/M arrest, DNA damage, apoptosis, autophapy and necrosis | [40] |
| Aspergillus | Xenograpsus testudinatus | Liquid medium: EtOAc Mycelia: Methanol | Clavatustide B | SW-480 - CCK-8 | Clavatustide B: 37% | Clavatustide B inhibits G1/S phase cell cycle transit | [28] |
| Aspergillus | Solar saltern | Liquid medium: EtOAc Mycelia: Ethanol | Ergosterol (1) Rosellichalasin (2) Cytochalasin E (3) | RKO DMSO MTT | (1): 3.3 μM; (2): 62.3 μM; (3): 37.3 μM | - | [27] |
| Aspergillus | Malus halliana | EtOAc | Asperterone B and C | SW-1116 - MTT | Asperterones B (57.5 µM), C (1.0 µM) | - | [41] |
| Aspergillus | Eudistoma vannamei | Liquid medium: EtOAc Mycelia: Methanol | Isocoumarin (R)-mellein (1) Penicillic acid (2) cis-4-hydroxymellein (3) trans-4-hydroxymellein (4) | HCT-8 DMSO MTT | (1), (3) and (4): >25 μg/mL; (2): 8.76 μg/ml | - | [26] |
| Aspergillus | Marine sediment | EtOAc | Acetylaranotin (1) Acetylapoaranotin (2) Deoxyapoaranotin (3) | HCT-116 - MTT | (1): 21.2 μmol/L; (2): 13.8 μmol/L; (3): 52 μmol/L | All compounds induce caspases 3-, 9- and 8-dependent apoptosis | [42] |
| Dichotomomyces (Aspergillus) | Lobophytum crassum | EtOAc | Pityriacitrin | HCT-116 Alone SRB | Pityriacitrin: 35.1 µM | - | [43] |
| Dichotomomyces (Aspergillus) | Marine sediment | EtOAc | Bis(dethio)bis(methylsulfanyl)gliotoxin (1) 6-acetylbis(dethio)bis(methylsulfanyl)gliotoxin (2) Acetylgliotoxin G (3) Gliotoxin (4) Acetylgliotoxin (5) Fiscalin B (6) | HCT-116 DMSO MTT | (1): 23.56 μg/mL; (2): 35.97 μg/mL; (3): 1.06 μg/mL; (4): 0.41 μg/mL; (5): >50 μg/mL; (6): 33.51 μg/mL | - | [44] |
| Emericella Aspergillus) | Soil | Sonication, centrifugation and lyophilization | - | CaCo-2 - MTT | Crude extract: 9.84 μg/mL | - | [25] |
| Eurotium (Aspergillus) | - | Ethanol | 1,8-Dihydroxy-3-methoxy-6-methyl- anthraquinone | HCT-116 DMSO MTT | Eurotium extract: 125.0 μg/mL 1,8-Dihydroxy-3-methoxy-6-methyl- anthraquinone: 18.6 μg/mL | - | [23] |
| Neosartorya Aspergillus Talaromyces | Aka coralliphaga Porites lutea Coastal forest soil Rhabdermia sp. Chondrilla australiensis Clathria reianwardii | EtOAc | - | HCT-116 and HT29 Alone or combined with Dox MTT | - | - | [45] |
| Neosartorya | Rumphella sp. | EtOAc | Chevalone C (1) Nortryptoquivaline (2) Tryptoquivaline H (3) Fiscalin A (4) epi-fiscalin A (5) and C (6) epi-neofiscalin A (7) | HCT-116 DMSO MTT | (1): 153 μM; (2): 114 μM; (3): 202 μM; (4): 123 μM; (5): 277 μM, (6): 86 μM; (7): 203 μM | - | [46] |
| Neosartorya | Aka coralliphaga Coastal forest soil Porites lutea | EtOAc | - | HCT-116 and HT-29 DMSO MTT | Extracts 1 (HCT-116 and HT-29: >200 μg/mL), 2 (HCT-116: 139 μg/mL, HT-29: 200 μg/mL), 3 (HCT-116: 189 μg/mL, HT29: 196 μg/mL) | - | [47] |
| Neosartorya | Acanthaster planci | EtOAc | 1,2,3,4-Tetrahydro-2-methyl-3-methylene-1,4-dioxopyrazino [1,2-a]índole (1) 1,2,3,4-Tetrahydro-2-methyl-1,3,4-trioxopyrazino [1,2-a]índole (2) Gliotoxin (3) Acetylgliotoxin (4) Reduced gliotoxin (5) 6-Acetylbis(methylthio)gliotoxin (6) Bisdethiobis(methylthio)gliotoxin (7) Didehydrobisdethiobis(methylthio)gliotoxin (8) Bis-N-norgliovictin (9) | HCT-116 and RKO DMSO MTS | (1): HCT-116: 10.34 μM, RKO: 33.56 μM; (2), (6), (8) and (9): HCT-116 and RKO: >50 μM; (3) and (4): HCT-116: 1.24 μM, RKO: 0.80 μM; (5): HCT-116: 0.89 μM, RKO: 1.24 μM; (7): HCT-116: 8.59 μM, RKO: 10.32 μM | - | [48] |
| Penicillium | Sonneratia sp. | EtOAc | - | HCT-116 DMSO SRB | Eupenicillium sp.: 13.9 μg/mL P. decumbens: 0.2 μg/mL | - | [49] |
| Penicillium | Isodon | EtOAc | Isopenicin A | SW-480 DMSO MTT | Isopenicin A: 8.33 μmol/L | Isopenicin A regulates cycle progression from G2 to M and induces apoptosis | [50] |
| Penicillium | - | Methanol | Arenicolin A | HCT-116 - CellTiter-Glo | Arenicolin A: 7.3 μM | - | [21] |
| Penicillium | Anemonia sulcata | EtOAc | cis-bis(methylthio)silvatin | CaCo-2 and HCT-116 DMSO MTT | cis-bis(methylthio)silvatin: HCT-116 29.29 µM, CaCo-2 35.31 µM | - | [51] |
| Penicillium | Marine water | - | Exopolysaccharides | CaCo-2 - SRB | Exopolysaccharides: 3.21 mg/mL | - | [52] |
| Penicillium | Trichocolea tomentella | EtOAc | Epoxydon (1) 3,6,8-trihydroxy-1-methylxanthone (2) Gentisyl alcohol (3) (R,S)-1-phenyl-1,2-ethanediol (4) Dehydrodechlorogriseofulvin (5) Dechlorogriseofulvin (6) Griseofulvin (7) Ethylene glycol benzoate (8) Alternariol (9) Griseoxanthone C (10) Drimiopsin H (11) Griseophenone B (12) and C (13) | HT-29 - - | (1): 14.1 μM; (2), (4), (5), (6), (7), (8), (9), (10), (11), (12) and (13): >20 μM; (3): 6.4 μM | - | [53] |
| Penicillium | Lake | EtOAc | - | CaCo-2 DMSO WST-1 and RTCA-DP | EAE: 55.2 μg/mL | EAE decreases angiogenesis and metastasis gene expression, cell migration and invasively | [54] |
| Penicillium | Plant leaf | EtOAc | Kongiiline A (1), B (2) Pebrolide (3) 1-deoxypebrolide (4) Asperphenamate (5) Asperphenamate B (6), C (7) N-benzoyl-phenylalaninol (8) Orsellinic acid (9) Mycophenolic acid (10) 5,7-dihydroxy-4-methylphthalide (11) | HCT-116 DMSO MTT | (1), (2), (3), (4), (8), (9) and (11): 100 μM; (5): 88.16 μM; (6): 77.68 μM; (7): 91.72 μM; (10): 36.92 μM | - | [55] |
| Penicillium | Marine sediment | Liquid medium: Methanol Mycelia: 80% acetone/ H2O | (–)-Brevianamide C | HCT-116 DMSO SRB | (–)-Brevianamide C: 15.6 µM | - | [29] |
| Penicillium | Isurus oxyrinchus | Liquid medium: EtOAc Mycelia: Methanol | Fructigenine A (1) Verrucosidin (2) Norverrucosidin (3) | HCT-116 - Crystal violet | (1): 40.5 μg/mL; (2): 30.8 μg/mL; (3): 5.7 μg/ml | - | [30] |
| Penicillium | - | EtOAc | - | CaCo-2 DMSO WST-1 and RTCA-DP | EAE: 102 μg/mL | EAE induces apoptosis | [56] |
| Penicillium | Marine sediment | Acetone | Penipacids A and E | RKO - MTT | Penipacids A (8.4 μM), E (9.7 μM) | - | [57] |
| Penicillium | Terminalia chebula Retz | EtOAc Ethanol | - | CaCo-2 DMSO MTT | EAE of IR-4 (55 μg/mL), IR-6 (44 μg/mL), IR-7 (67 μg/mL) Ethanol extract of IR-6: 71 μg/mL | - | [22] |
| Genus | Isolated from | Extraction | Isolated Compounds | Cell Line/Administration/Cytotoxicity Assay | Compound and IC50 or Cell Death (%) | Mechanism of Action | Reference |
|---|---|---|---|---|---|---|---|
| Beauveria | Marine sponge | Acetone | 1-Hydroxy-10-methoxy-dibenz[b, e]oxepin-6,11-dione (1) Chrysazin (2) Globosuxanthone A (3) | HCT-15 - MTT | (1) and (2): >30 μM; (3): 10.7 μM | - | [72] |
| Bionectria | Psidium guajava | Ethanol | Exopolysaccharides | HT-29 Alone MTT | Exopolysaccharides (84.58%) | - | [73] |
| Cordyceps | - | Methanol | - | HCT-116 and SW-480 DMSO MTT | Extract: HCT-116: >250 μg/mL; SW-480: 178.70 μg/mL | Extract reduces cell migration and cytoplasmic β-catenin | [63] |
| Cordyceps | - | Ethanol | - | RKO Distilled water CCK-8 | - | Extract induces cell morphological changes, G2/M phase arrest and apoptosis | [64] |
| Cordyceps | - | Methanol:water (80:20 v/v) | - | HCT-15 - SRB | Extract: 72.57 μg/mL | - | [62] |
| Cordyceps | - | Methanol Buthanol | - | HT-29 Alone EZ-CyTox kit | GSC (46.56%) BuOH extract of C. militaris (36.23%) | GSC induces cell morphological changes and G2/M phase arrest, blocking the cyclin B1 and Cdc25c protein | [65] |
| Engyodontium | Terminalia sp. | Ethyl acetate (EtOAc) | - | HCT-116 DMSO SRB | EtOAc extract (EAE): 2.5 μg/mL | - | [49] |
| Fusarium | Soil | EtOAc | - | HCT-8 and HCT-116 DMSO CellTiter-Glo assay | EAE: HCT-8: 0.3779 μg/mL, HCT-116: 15.86 μg/mL | - | [67] |
| Fusarium | - | - | Camptothecin | CaCo-2 DMSO:Methanol (1:50) Alamar blue assay | Standard camptothecin: 2.41 μM; Crude camptothecin extract: 0.591 μM | - | [68] |
| Fusarium | Soil | Sonication, centrifugation and lyophilization | - | CaCo-2 - MTT | Crude extract: 6.24 μg/mL | - | [25] |
| Metarhizium | - | Liquid medium: Filtration Acetonitrile and sodium chloride | Destruxin A (1), B (2) and E (3) | CaCo-2 and HCT-116 DMSO MTT | (1): CaCo-2 (2.18 μM), HCT-116 (2.06 μM); (2): CaCo-2 (1.34 μM), HCT-116 (3.22 μM); (3): CaCo-2 (0.05 μM), HCT-116 (0.04 μM) | (3) activates apoptotic caspases and induces ROS All destruxins induce G0/G1 phase arrest in CaCo-2 cells, reduce cell migration, have antiangiogenic activity and interfered with the MAPK and/or PI3K/Akt signaling pathways | [74] |
| Myrothecium | Calotropis procera | Liquid medium: Chloroform and methanol (2:1 v/v) | - | HCT-116 DMSO Crystal violet staining | Extract: 380 ng/mL | - | [75] |
| Trichoderma | Dysidea sp. | EtOAc | Trichodermaloid A (1), B (2) and C (3) Aspergilloid G (4) Rhinomilisin E (5) and G (6) | SW-620 DMSO MTT | (1): 9.3 μM; (2): 8.6 μM; (3): 12.7 μM; (4) and (6): >32 μM; (5): 22.7 μM | - | [71] |
| Trichoderma | Polygonum aviculare | Ethanol | - | HCT-116 DMSO MTT | Fractions F2 (14.9 μg/mL), F4 (7.3 μg/mL), F5 (7.61 μg/mL) | - | [24] |
| Trichoderma | Stylissa flabelliformis | EtOAc | - | WiDr - MTT | EAE: 88.88 μg/mL | EAE induces apoptosis | [70] |
| Trichoderma Fusarium | Bacopa monnieri | Methanol | - | HCT-116 DMSO MTT | B1, B20, BX1: O and A (>100 μg/mL); T. aureoviride: O (11 μg/mL), A (>100 μg/mL); Fusarium sp. 6241: 5 μg/mL; F. oxysporum: O (22 μg/mL), A (98.68 μg/mL) | - | [66] |
| Genus | Isolated from | Extraction | Isolated Compounds | Cell Line/ /Administration Cytotoxicity Assay | Compound and IC50 or Cell Death (%) | Reference |
|---|---|---|---|---|---|---|
| Alternaria | Miquelia dentata | Methanol Ethyl acetate (EtOAc) | - | SW-480 and HCT-116 - Hoechst 33342 | Methanol extract: HCT-116 (5.39 μg/mL), SW-480 (12.37 μg/mL); EAE: HCT-116 (6.59 μg/mL), SW-480 (7.2 μg/mL) | [77] |
| Alternaria | Erythrophleum fordii | Ethanol | (6aR, 6bS, 7S)-3, 6a, 7, 10-tetra-hydroxy-4, 9-dioxo-4, 6a, 6b, 7, 8, 9-hexahydroperylene (1) | HCT-8 DMSO MTT | (1): 1.78 μmol/L | [78] |
| Bipolaris | Soil | EtOAc | - | HCT-8 and HCT-116 DMSO CellTiter-Glo assay | Extract: HCT-8 (202.5 μg/mL), HCT-116 (18.97 μg/mL) | [67] |
| Drechslera | - | Ethanol | di-2-ethylhexyl phthalate | HCT-116 DMSO MTT | Drechslera extract: 104.0; di-2-ethylhexyl phthalate: 9.5 | [23] |
| Paradendryphiella | Pomacea canaliculata | EtOAc | (3R, 6R) hyalodendrin | SW-48, DLD-1, LS513, LOVO, RKO, LS174T, SW-480 HT-29 and HCT-116 DMSO MTT | (3R, 6R) hyalodendrin: SW-48 (149.0 nM), DLD-1 (40.0 nM), HT-29 (58.0 nM), HT-29 5FU (146.8 nM), HT-29 oxa (141.8 nM), HT-29 SN-38 (93.8 nM), HCT-116 (48.0 nM), HCT-116 5FU (72.0 nM), HCT-116 oxa (25.7 nM), HCT-116 SN-38 (43.8 nM), LS513 (78.0 nM), LOVO (73.4 nM), RKO (74.3 nM), LS174T (158.0 nM), SW-480 (163.7 nM) | [80] |
| Phoma Curvularia Pleosporales sp. Alternaria | Catharanthus roseus Euphorbia prostrata Calotropis procera | EtOAc | - | HT-29 and HCT-116 - MTT | C. aeria extract: HT29 (74.5 μg/mL), HCT-116 (53.9 μg/mL); Pleosporales sp. extract: HT29 (69.4 μg/mL), HCT-116 (36.7 μg/mL); P. multirostrata extract: HT29 and HCT-116 (>100 μg/mL); C. australiensis extracts 1: HT29 (54.3 μg/mL), HCT-116 (25.6 μg/mL); 2: HT29 (>100 μg/mL), HCT-116 (59.7 μg/mL); A. alternata extract: HT29 (>100 μg/mL), HCT-116 (52.5 μg/mL); Alternaria sp. extract: HT29 (28.4 μg/mL), HCT-116 (29.1 μg/mL) | [39] |
| Pleosporales sp. | Bacopa monnieri | Dichloromethane | - | HCT-116 DMSO MTT | CK01: O (12 μg/mL), A (>100 μg/mL) | [66] |
| Setophoma | Leaf litter | Methanol/ Chloroform (1:1) | Secalonic acid A (1), E (2) and G (3) Penicillixanthone A (4) and B (5) Blennolide J (6) Hypothemycin (7) | SW-620 DMSO CellTiter 96 AQueous One Solution Cell Proliferation Assay | (1): 0.41 μM; (2): 19.12 μM; (3): 3.67 μM; (4): 0.21 μM); (5): 5.55 μM; (6): 6.14 μM; (7): 2.14 μM | [79] |
| Genus | Isolated from | Extraction | Isolated Compounds | Cell Line/ /Administration Cytotoxicity Assay | Compound and IC50 or Cell Death (%) | Mechanism of Action | Reference |
|---|---|---|---|---|---|---|---|
| Chaetomium | - | Liquid medium: Ethyl acetate (EtOAc) Mycelia: Methanol | Chaetocochin (Ch.) J | RKO, SW-480 and HCT-116 - CCK8 | Ch. J: RKO (0.56 μM), SW-480 (0.61 μM), HCT-116 (0.65 μM) | Ch. J. induces apoptosis, autophagy and activates AMPK and PI3K/AKT/mTOR signaling pathway | [83] |
| Chaetomium | - | EtOAc | - | HCT-116 DMSO SRB | C. globosum extract: 1.2 μg/ml | - | [49] |
| Chaetomium | Soil | EtOAc | - | HCT-8 and HCT-116 DMSO CellTiter-Glo assay | Ethyl acetate extract (EAE): HCT-8 (8.744 μg/mL), HCT-116 (152.8 μg/mL) | - | [67] |
| Chaetomium | Cymbidium goeringii | Liquid medium: EtOAc Mycelia: Methanol | Ch. A and C | SW-480 Alone MTT | Ch. A: 15.21 μM Ch. C: 0.63 μM | Ch. C. induces G2/M arrest, apoptosis, activation of the caspase 3 and PARP degradation, increased Bax and decreased Bcl-2 level | [82] |
| Chaetomium | Trigonella foenum-graecum | EtOAc | - | HT-29 and HCT-116 - MTT | C. globosum extract: HT-29 (>100 μg/mL), HCT-116 (75.2 μg/mL) | - | [39] |
| Chaetomium | Ginkgo biloba | Methanol | Chaetoglobosin A (1), G (2), V (3), Vb (4), E (5), F (6), Fex (7), Fa (8) 20-dihydrochaetoglobosin A (9) | HCT-116 Alone SRB | (1): 3.15 μM; (2): 65.6 μM; (3): 29.5 μM; (4): 18.4 μM; (5): >100 μM; (6): 17.8 μM; (7): 17.8 μM; (8): 5.85 μM; (9): 8.44 μM | - | [81] |
| Trichlocladium | Soil | EtOAc | Trichocladinols E (1), F (2) and G (3) | SW-480 and HCT-116 DMSO MTS | (1): SW-480 (54.9 μM), HCT-116 (48.8 μM); (2): SW-480 (51.9 μM), HCT-116 (56.6 μM); (3): SW-480 (43.6 μM), HCT-116 (41.7 μM) | - | [84] |
| Scytalidium | - | Methanol/Chloroform (1:1) | [5’-formyl-2’-hydroxyl-4’-methoxy-(E,E)-sorbophenone (1) Scalbucillin B (2) 1-(2’-hydroxy-4’-methoxy-5’-methylphenyl)-2,4-E,E-hexadien-1-one (3) 5′-formyl-2′-hydroxy-4′-methoxy-(E)-4-hexenophenone (4) | SW-620 DMSO CellTiter 96® AQueous One Solution Cell Proliferation Assay | (1): 0.5 μM; (2): 16 μM; (3): 15.1 μM; (4): 2.5 μM | - | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luque, C.; Cepero, A.; Perazzoli, G.; Mesas, C.; Quiñonero, F.; Cabeza, L.; Prados, J.; Melguizo, C. In Vitro Efficacy of Extracts and Isolated Bioactive Compounds from Ascomycota Fungi in the Treatment of Colorectal Cancer: A Systematic Review. Pharmaceuticals 2023, 16, 22. https://doi.org/10.3390/ph16010022
Luque C, Cepero A, Perazzoli G, Mesas C, Quiñonero F, Cabeza L, Prados J, Melguizo C. In Vitro Efficacy of Extracts and Isolated Bioactive Compounds from Ascomycota Fungi in the Treatment of Colorectal Cancer: A Systematic Review. Pharmaceuticals. 2023; 16(1):22. https://doi.org/10.3390/ph16010022
Chicago/Turabian StyleLuque, Cristina, Ana Cepero, Gloria Perazzoli, Cristina Mesas, Francisco Quiñonero, Laura Cabeza, Jose Prados, and Consolación Melguizo. 2023. "In Vitro Efficacy of Extracts and Isolated Bioactive Compounds from Ascomycota Fungi in the Treatment of Colorectal Cancer: A Systematic Review" Pharmaceuticals 16, no. 1: 22. https://doi.org/10.3390/ph16010022