
Experimental Studies Indicate That ST-2223, the Antagonist of Histamine H3 and Dopamine D2/D3 Receptors, Restores Social Deficits and Neurotransmission Dysregulation in Mouse Model of Autism

 , and
, and 
Abstract
:1. Introduction
2. Results
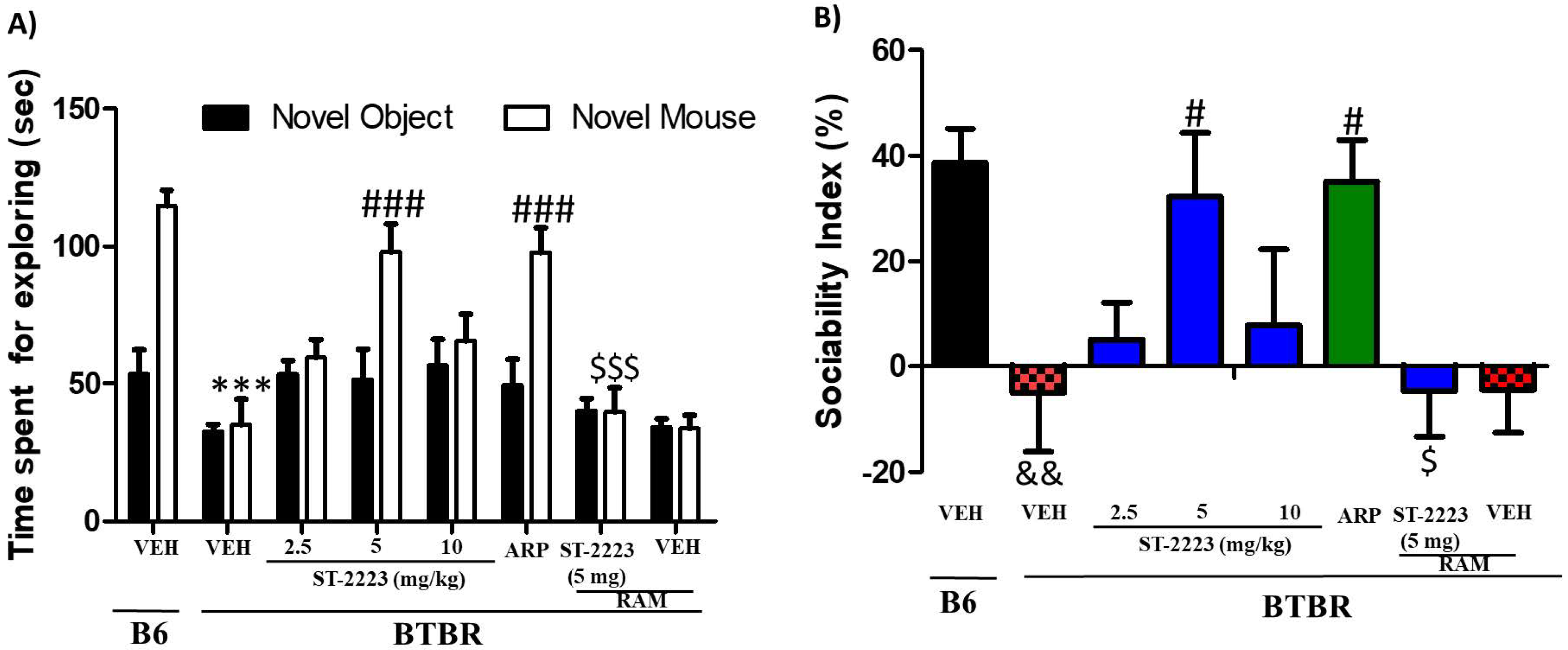
2.1. Effects of Systemic Administration of ST-2223 on Sociability Deficits in BTBR Mice
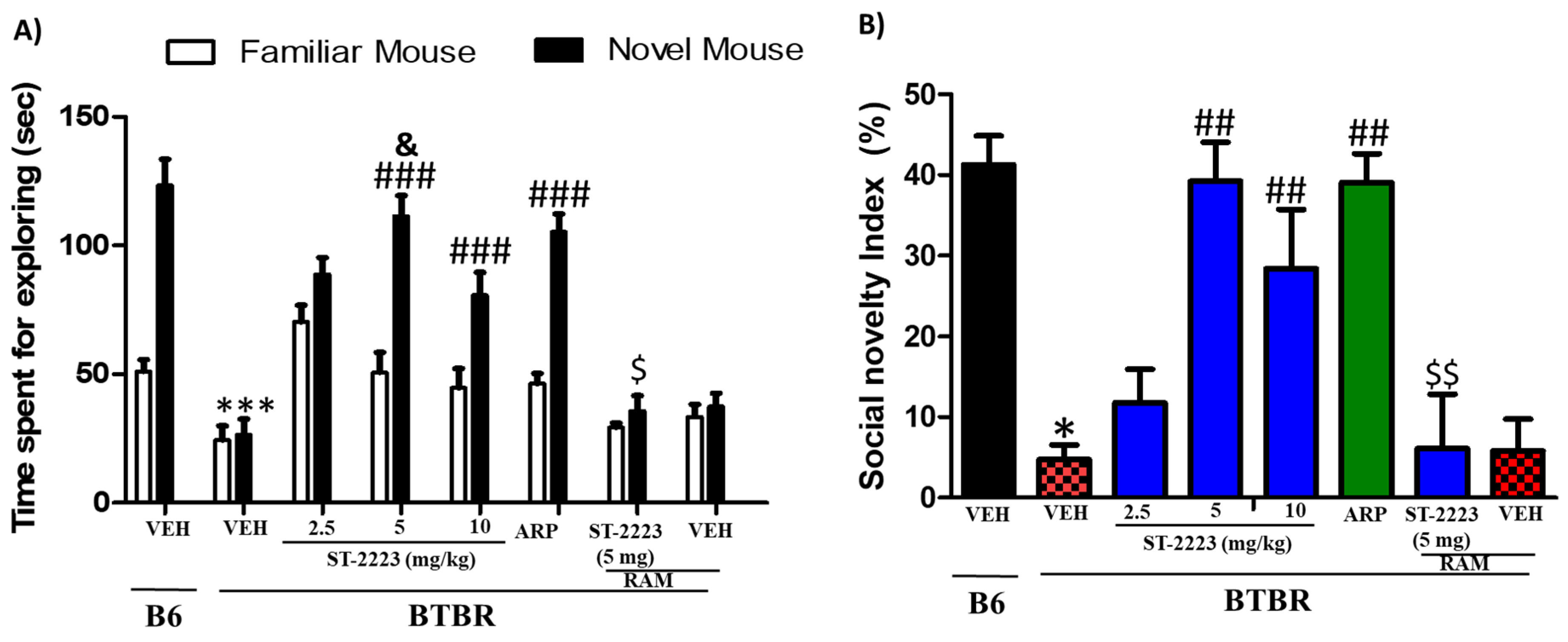
2.2. Effects of Systemic Administrtion of ST-2223 on Social Preference in BTBR Mice
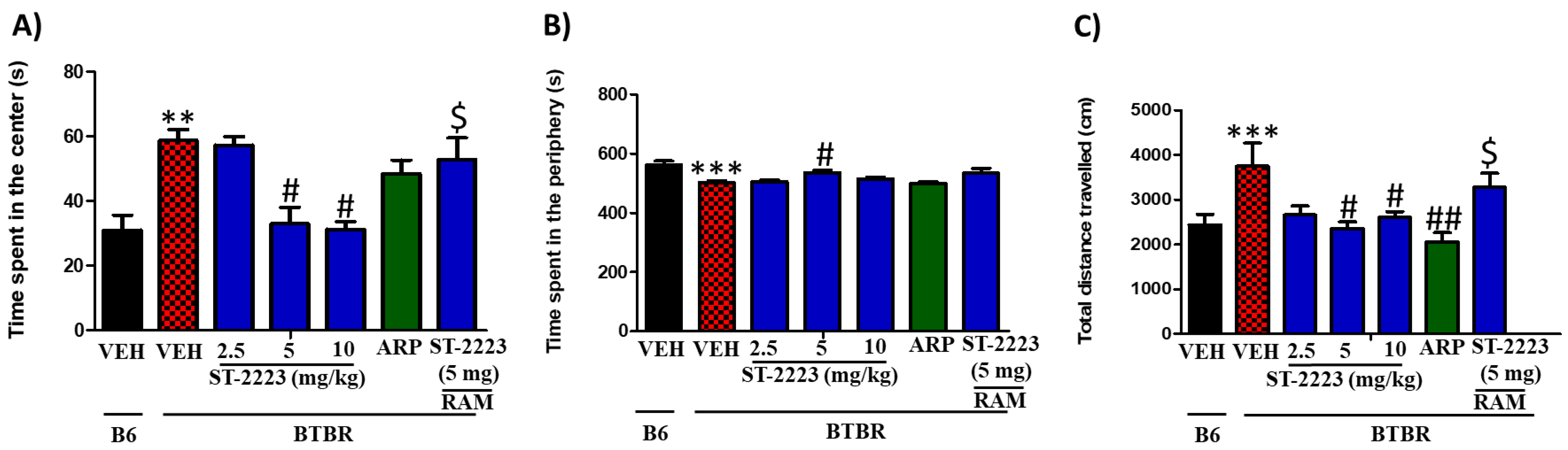
2.3. Effects of Systemic Administration of ST-2223 on Anxiety and Locomotor Activity in BTBR Mice in OFL Test
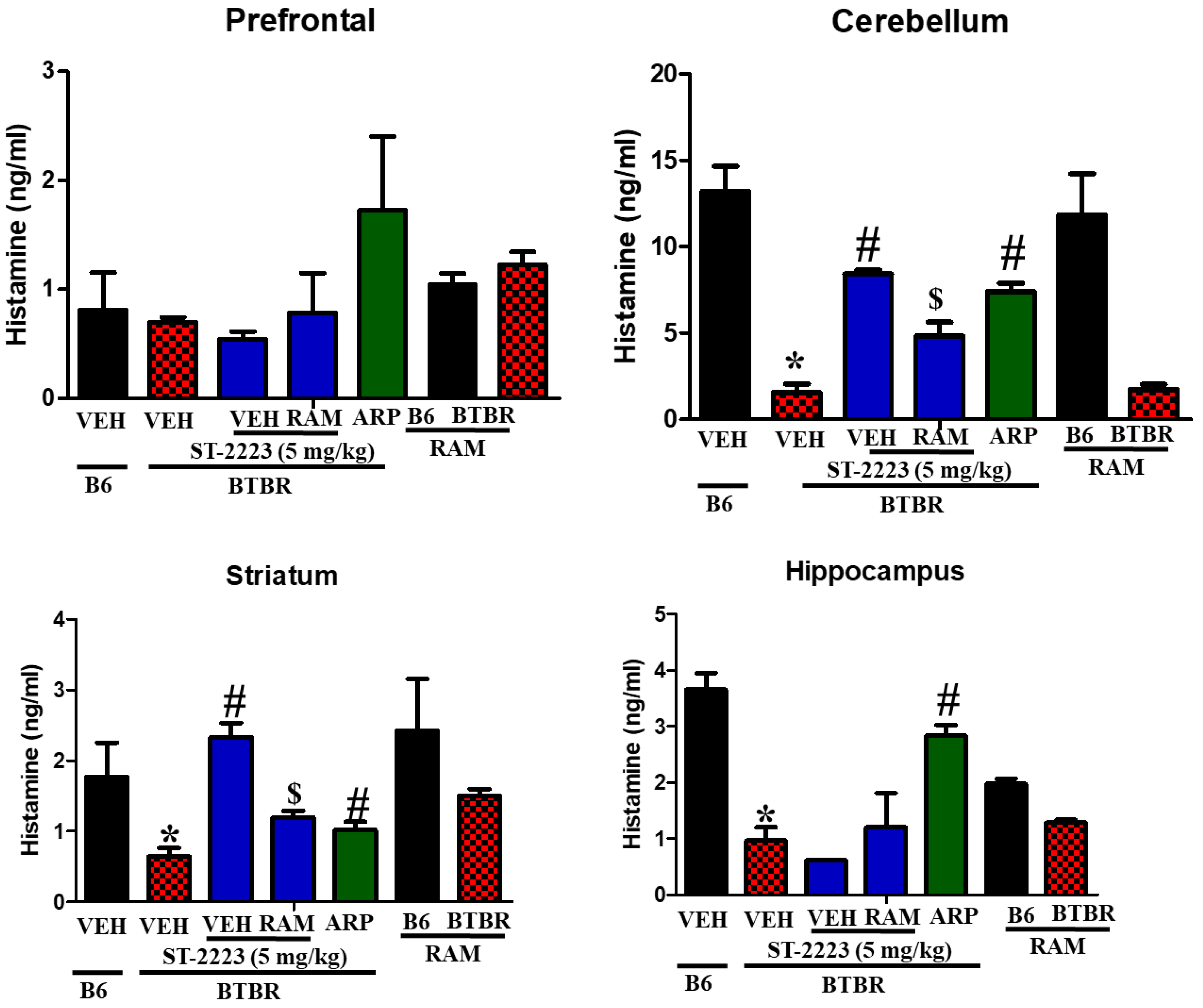
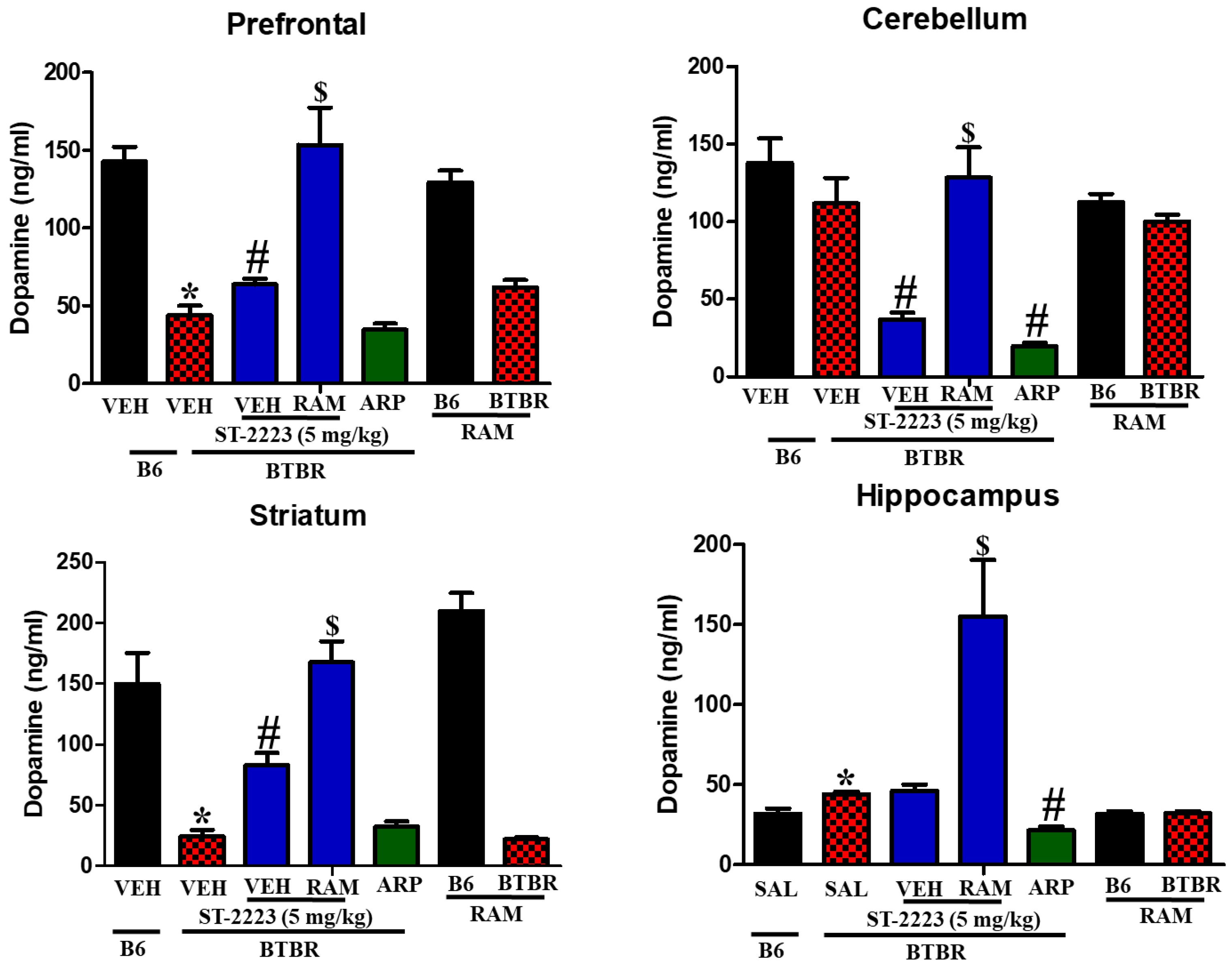
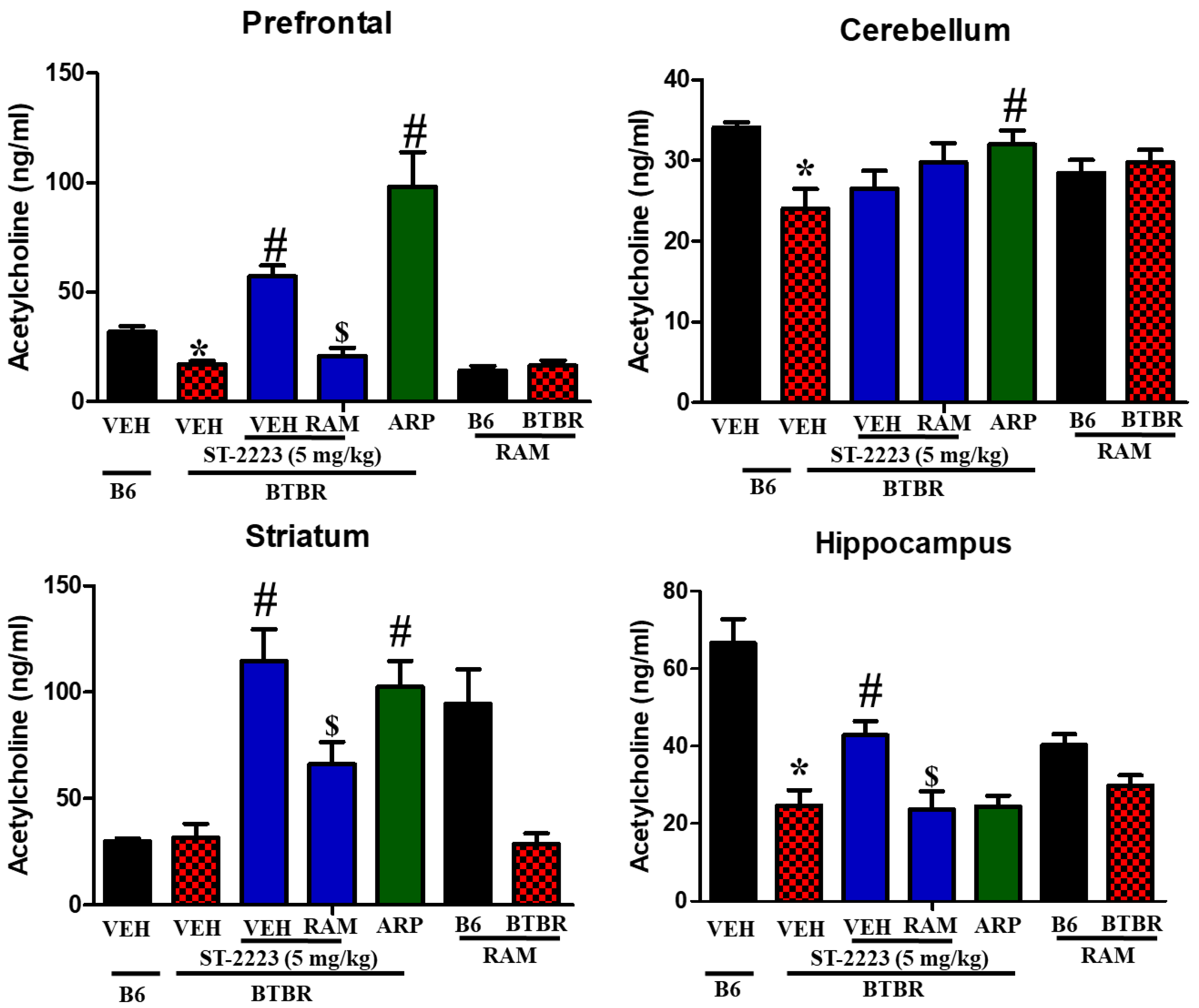
2.4. Effects of ST-2223 on the Brain Levels of Histamine, Dopamine, and Acetylcholine in Different Brain Parts of BTBR Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
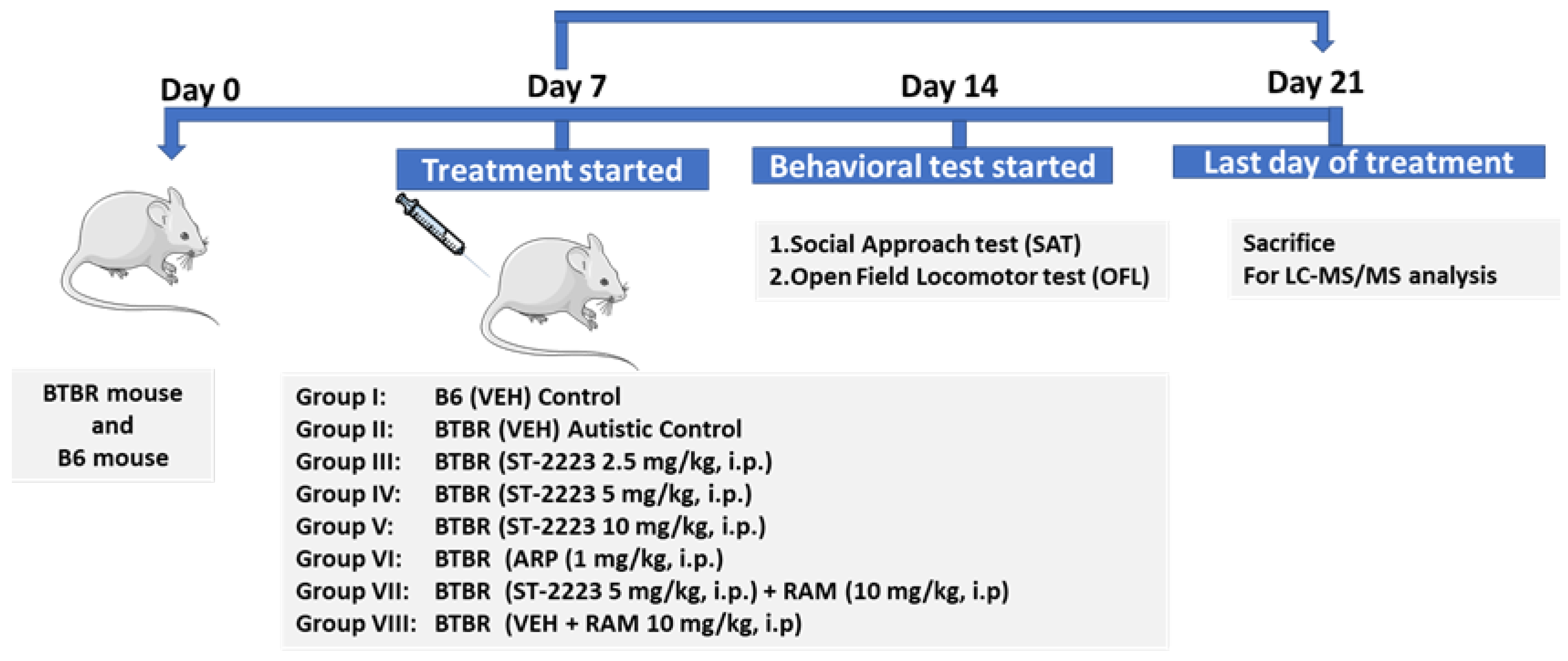
4.3. Study Design and Treatments
4.4. Behavioral Tests
4.4.1. Social Approach Test (SAT)
4.4.2. Open Field Locomotor Test (OFL)
4.5. Sample Preparation and LC-MS/MS Conditions
4.6. Statistics
5. Conclusions
6. Study Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choudhury, P.R.; Lahiri, S.; Rajamma, U. Glutamate mediated signaling in the pathophysiology of autism spectrum disorders. Pharmacol. Biochem. Behav. 2012, 100, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.H.; Chan, W.Y.; Lee, K.K.; Fan, M.; Yew, D.T. Neurotransmitters, neuropeptides and calcium binding proteins in developing human cerebellum: A review. Histochem. J. 2000, 32, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Marotta, R.; Risoleo, M.C.; Messina, G.; Parisi, L.; Carotenuto, M.; Vetri, L.; Roccella, M. The Neurochemistry of Autism. Brain Sci. 2020, 10, 163. [Google Scholar] [CrossRef] [Green Version]
- Hanks, C.E.; Lewin, A.B.; Mutch, P.J.; Storch, E.A.; Murphy, T.K. Social Deficits and Autism Spectrum Disorders in Tourette’s Syndrome. Curr. Dev. Disord. Rep. 2015, 2, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Eissa, N.; Al-Houqani, M.; Sadeq, A.; Ojha, S.K.; Sasse, A.; Sadek, B. Current Enlightenment About Etiology and Pharmacological Treatment of Autism Spectrum Disorder. Front. Neurosci. 2018, 12, 304. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, H.Y.; McGurk, S.R. The effects of clozapine, risperidone, and olanzapine on cognitive function in schizophrenia. Schizophr. Bull. 1999, 25, 233–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.V.; Goldman-Rakic, P.S. Modulation of memory fields by dopamine D1 receptors in prefrontal cortex. Nature 1995, 376, 572–575. [Google Scholar] [CrossRef]
- Çetin, F.; Tunca, H.; Guney, E.; Iseri, E. Neurotransmitter Systems in Autism Spectrum Disorder. In Autism Spectrum Disorder—Recent Advances; Fitzgerald, M., Ed.; InTech Open: London, UK, 2015; pp. 15–30. [Google Scholar] [CrossRef] [Green Version]
- Yasui, D.H.; Scoles, H.A.; Horike, S.; Meguro-Horike, M.; Dunaway, K.W.; Schroeder, D.I.; Lasalle, J.M. 15q11.2-13.3 chromatin analysis reveals epigenetic regulation of CHRNA7 with deficiencies in Rett and autism brain. Hum. Mol. Genet. 2011, 20, 4311–4323. [Google Scholar] [CrossRef]
- Martin-Ruiz, C.M.; Lee, M.; Perry, R.H.; Baumann, M.; Court, J.A.; Perry, E.K. Molecular analysis of nicotinic receptor expression in autism. Brain Res. Mol. Brain Res. 2004, 123, 81–90. [Google Scholar] [CrossRef]
- Tan, C.C.; Yu, J.T.; Wang, H.F.; Tan, M.S.; Meng, X.F.; Wang, C.; Jiang, T.; Zhu, X.C.; Tan, L. Efficacy and safety of donepezil, galantamine, rivastigmine, and memantine for the treatment of Alzheimer’s disease: A systematic review and meta-analysis. J. Alzheimers Dis. 2014, 41, 615–631. [Google Scholar] [CrossRef]
- Potter, A.S.; Dunbar, G.; Mazzulla, E.; Hosford, D.; Newhouse, P.A. AZD3480, a novel nicotinic receptor agonist, for the treatment of attention-deficit/hyperactivity disorder in adults. Biol. Psychiatry 2014, 75, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Preskorn, S.H.; Gawryl, M.; Dgetluck, N.; Palfreyman, M.; Bauer, L.O.; Hilt, D.C. Normalizing effects of EVP-6124, an alpha-7 nicotinic partial agonist, on event-related potentials and cognition: A proof of concept, randomized trial in patients with schizophrenia. J. Psychiatr. Pract. 2014, 20, 12–24. [Google Scholar] [CrossRef] [PubMed]
- McTighe, S.M.; Neal, S.J.; Lin, Q.; Hughes, Z.A.; Smith, D.G. The BTBR mouse model of autism spectrum disorders has learning and attentional impairments and alterations in acetylcholine and kynurenic acid in prefrontal cortex. PLoS ONE 2013, 8, e62189. [Google Scholar] [CrossRef] [PubMed]
- Karvat, G.; Kimchi, T. Acetylcholine elevation relieves cognitive rigidity and social deficiency in a mouse model of autism. Neuropsychopharmacology 2014, 39, 831–840. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, G.E.; Aguilar, J.I.; Matthies, H.J.; Harrison, F.E.; Bundschuh, K.E.; West, A.; Hashemi, P.; Herborg, F.; Rickhag, M.; Chen, H.; et al. Autism-linked dopamine transporter mutation alters striatal dopamine neurotransmission and dopamine-dependent behaviors. J. Clin. Investig. 2019, 129, 3407–3419. [Google Scholar] [CrossRef] [PubMed]
- Squillace, M.; Dodero, L.; Federici, M.; Migliarini, S.; Errico, F.; Napolitano, F.; Krashia, P.; Di Maio, A.; Galbusera, A.; Bifone, A.; et al. Dysfunctional dopaminergic neurotransmission in asocial BTBR mice. Transl. Psychiatry 2014, 4, e427. [Google Scholar] [CrossRef] [Green Version]
- Dichter, G.S.; Felder, J.N.; Green, S.R.; Rittenberg, A.M.; Sasson, N.J.; Bodfish, J.W. Reward circuitry function in autism spectrum disorders. Soc. Cogn. Affect. Neurosci. 2012, 7, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Hagenow, S.; Affini, A.; Pioli, E.Y.; Hinz, S.; Zhao, Y.; Porras, G.; Namasivayam, V.; Müller, C.E.; Lin, J.-S.; Bezard, E.; et al. Adenosine A2AR/A1R Antagonists Enabling Additional H3R Antagonism for the Treatment of Parkinson’s Disease. J. Med. Chem. 2021, 64, 8246–8262. [Google Scholar] [CrossRef]
- Proschak, E.; Stark, H.; Merk, D. Polypharmacology by Design: A Medicinal Chemist’s Perspective on Multitargeting Compounds. J. Med. Chem. 2019, 62, 420–444. [Google Scholar] [CrossRef]
- Khan, N.; Saad, A.; Nurulain, S.M.; Darras, F.H.; Decker, M.; Sadek, B. The dual-acting H3 receptor antagonist and AChE inhibitor UW-MD-71 dose-dependently enhances memory retrieval and reverses dizocilpine-induced memory impairment in rats. Behav. Brain Res. 2016, 297, 155–164. [Google Scholar] [CrossRef]
- Butini, S.; Nikolic, K.; Kassel, S.; Bruckmann, H.; Filipic, S.; Agbaba, D.; Gemma, S.; Brogi, S.; Brindisi, M.; Campiani, G.; et al. Polypharmacology of dopamine receptor ligands. Prog. Neurobiol. 2016, 142, 68–103. [Google Scholar] [CrossRef]
- Khanfar, M.A.; Affini, A.; Lutsenko, K.; Nikolic, K.; Butini, S.; Stark, H. Multiple Targeting Approaches on Histamine H3 Receptor Antagonists. Front. Neurosci. 2016, 10, 201. [Google Scholar] [CrossRef] [Green Version]
- Speranza, L.; di Porzio, U.; Viggiano, D.; de Donato, A.; Volpicelli, F. Dopamine: The Neuromodulator of Long-Term Synaptic Plasticity, Reward and Movement Control. Cells 2021, 10, 735. [Google Scholar] [CrossRef] [PubMed]
- Ghaeli, P.; Nikvarz, N.; Alaghband-Rad, J.; Alimadadi, A.; Tehrani-Doost, M. Effects of risperidone on core symptoms of autistic disorder based on childhood autism rating scale: An open label study. Indian J. Psychol. Med. 2014, 36, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougle, C.J.; Scahill, L.; Aman, M.G.; McCracken, J.T.; Tierney, E.; Davies, M.; Arnold, L.E.; Posey, D.J.; Martin, A.; Ghuman, J.K.; et al. Risperidone for the core symptom domains of autism: Results from the study by the autism network of the research units on pediatric psychopharmacology. Am. J. Psychiatry 2005, 162, 1142–1148. [Google Scholar] [CrossRef]
- McCracken, J.T.; McGough, J.; Shah, B.; Cronin, P.; Hong, D.; Aman, M.G.; Arnold, L.E.; Lindsay, R.; Nash, P.; Hollway, J.; et al. Risperidone in children with autism and serious behavioral problems. N. Engl. J. Med. 2002, 347, 314–321. [Google Scholar] [CrossRef]
- Scahill, L.; Hallett, V.; Aman, M.G.; McDougle, C.J.; Eugene, A.L.; McCracken, J.T.; Tierney, E.; Deng, Y.; Dziura, J.; Vitiello, B.; et al. Brief Report: Social disability in autism spectrum disorder: Results from Research Units on Pediatric Psychopharmacology (RUPP) Autism Network trials. J. Autism Dev. Disord. 2013, 43, 739–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavăl, D. A Dopamine Hypothesis of Autism Spectrum Disorder. Dev. Neurosci. 2017, 39, 355–360. [Google Scholar] [CrossRef]
- Langen, M.; Durston, S.; Kas, M.J.; van Engeland, H.; Staal, W.G. The neurobiology of repetitive behavior:...and men. Neurosci. Biobehav. Rev. 2011, 35, 356–365. [Google Scholar] [CrossRef]
- Rodriguiz, R.M.; Chu, R.; Caron, M.G.; Wetsel, W.C. Aberrant responses in social interaction of dopamine transporter knockout mice. Behav. Brain Res. 2004, 148, 185–198. [Google Scholar] [CrossRef]
- Santangelo, A.; Passani, M.B.; Casarrubea, M. Brain histamine and behavioral neuroscience. Oncotarget 2017, 8, 16107–16108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Kaminga, A.C.; Wen, S.W.; Wu, X.; Acheampong, K.; Liu, A. Dopamine and Dopamine Receptors in Alzheimer’s Disease: A Systematic Review and Network Meta-Analysis. Front. Aging Neurosci. 2019, 11, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, C.; Shin, J.H.; Rajpurohit, A.; Deep-Soboslay, A.; Collado-Torres, L.; Brandon, N.J.; Hyde, T.M.; Kleinman, J.E.; Jaffe, A.E.; Cross, A.J.; et al. Altered expression of histamine signaling genes in autism spectrum disorder. Transl. Psychiatry 2017, 7, e1126. [Google Scholar] [CrossRef] [Green Version]
- von Coburg, Y.; Kottke, T.; Weizel, L.; Ligneau, X.; Stark, H. Potential utility of histamine H3 receptor antagonist pharmacophore in antipsychotics. Bioorg. Med. Chem. Lett. 2009, 19, 538–542. [Google Scholar] [CrossRef]
- Sander, K.; Kottke, T.; Weizel, L.; Stark, H. Kojic acid derivatives as histamine H(3) receptor ligands. Chem. Pharm. Bull. 2010, 58, 1353–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Baronio, D.; Castro, K.; Gonchoroski, T.; de Melo, G.M.; Nunes, G.D.; Bambini-Junior, V.; Gottfried, C.; Riesgo, R. Effects of an H3R antagonist on the animal model of autism induced by prenatal exposure to valproic acid. PLoS ONE 2015, 10, e0116363. [Google Scholar] [CrossRef] [Green Version]
- Fox, G.B.; Esbenshade, T.A.; Pan, J.B.; Radek, R.J.; Krueger, K.M.; Yao, B.B.; Browman, K.E.; Buckley, M.J.; Ballard, M.E.; Komater, V.A.; et al. Pharmacological properties of ABT-239 [4-(2-{2-[(2R)-2-Methylpyrrolidinyl]ethyl}-benzofuran-5-yl)benzonitrile]: II. Neurophysiological characterization and broad preclinical efficacy in cognition and schizophrenia of a potent and selective histamine H3 receptor antagonist. J. Pharmacol. Exp. Ther. 2005, 313, 176–190. [Google Scholar] [CrossRef] [Green Version]
- Fox, G.B.; Pan, J.B.; Esbenshade, T.A.; Bennani, Y.L.; Black, L.A.; Faghih, R.; Hancock, A.A.; Decker, M.W. Effects of histamine H(3) receptor ligands GT-2331 and ciproxifan in a repeated acquisition avoidance response in the spontaneously hypertensive rat pup. Behav. Brain Res. 2002, 131, 151–161. [Google Scholar] [CrossRef]
- Fox, G.B.; Pan, J.B.; Esbenshade, T.A.; Bitner, R.S.; Nikkel, A.L.; Miller, T.; Kang, C.H.; Bennani, Y.L.; Black, L.A.; Faghih, R.; et al. Differential in vivo effects of H3 receptor ligands in a new mouse dipsogenia model. Pharm. Biochem. Behav. 2002, 72, 741–750. [Google Scholar] [CrossRef]
- Fox, G.B.; Pan, J.B.; Radek, R.J.; Lewis, A.M.; Bitner, R.S.; Esbenshade, T.A.; Faghih, R.; Bennani, Y.L.; Williams, M.; Yao, B.B.; et al. Two novel and selective nonimidazole H3 receptor antagonists A-304121 and A-317920: II. In vivo behavioral and neurophysiological characterization. J. Pharmacol. Exp. Ther. 2003, 305, 897–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, A.; Kiss, D.J.; Keseru, G.M.; Stark, H. Binding kinetics of cariprazine and aripiprazole at the dopamine D3 receptor. Sci. Rep. 2018, 8, 12509. [Google Scholar] [CrossRef]
- Eissa, N.; Jayaprakash, P.; Azimullah, S.; Ojha, S.K.; Al-Houqani, M.; Jalal, F.Y.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. The histamine H3R antagonist DL77 attenuates autistic behaviors in a prenatal valproic acid-induced mouse model of autism. Sci. Rep. 2018, 8, 13077. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; et al. The Dual-Active Histamine H3 Receptor Antagonist and Acetylcholine Esterase Inhibitor E100 Alleviates Autistic-Like Behaviors and Oxidative Stress in Valproic Acid Induced Autism in Mice. Int. J. Mol. Sci. 2020, 21, 3996. [Google Scholar] [CrossRef]
- Eissa, N.; Jayaprakash, P.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. Simultaneous Blockade of Histamine H3 Receptors and Inhibition of Acetylcholine Esterase Alleviate Autistic-Like Behaviors in BTBR T+ tf/J Mouse Model of Autism. Biomolecules 2020, 10, 1251. [Google Scholar] [CrossRef]
- Brioni, J.D.; Esbenshade, T.A.; Garrison, T.R.; Bitner, S.R.; Cowart, M.D. Discovery of histamine H3 antagonists for the treatment of cognitive disorders and Alzheimer’s disease. J. Pharmacol. Exp. Ther. 2011, 336, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Stahl, S.M. Dopamine system stabilizers, aripiprazole, and the next generation of antipsychotics, part 2: Illustrating their mechanism of action. J. Clin. Psychiatry 2001, 62, 923–924. [Google Scholar] [CrossRef] [Green Version]
- Stahl, S.M. Dopamine system stabilizers, aripiprazole, and the next generation of antipsychotics, part 1, "Goldilocks" actions at dopamine receptors. J. Clin. Psychiatry 2001, 62, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Ligneau, X.; Perrin, D.; Landais, L.; Camelin, J.C.; Calmels, T.P.; Berrebi-Bertrand, I.; Lecomte, J.M.; Parmentier, R.; Anaclet, C.; Lin, J.S.; et al. BF2.649 [1-{3-[3-(4-Chlorophenyl)propoxy]propyl}piperidine, hydrochloride], a nonimidazole inverse agonist/antagonist at the human histamine H3 receptor: Preclinical pharmacology. J. Pharmacol. Exp. Ther. 2007, 320, 365–375. [Google Scholar] [CrossRef]
- Medhurst, A.D.; Atkins, A.R.; Beresford, I.J.; Brackenborough, K.; Briggs, M.A.; Calver, A.R.; Cilia, J.; Cluderay, J.E.; Crook, B.; Davis, J.B.; et al. GSK189254, a novel H3 receptor antagonist that binds to histamine H3 receptors in Alzheimer’s disease brain and improves cognitive performance in preclinical models. J. Pharmacol. Exp. Ther. 2007, 321, 1032–1045. [Google Scholar] [CrossRef] [Green Version]
- Chao, O.Y.; Pathak, S.S.; Zhang, H.; Dunaway, N.; Li, J.-S.; Mattern, C.; Nikolaus, S.; Huston, J.P.; Yang, Y.-M. Altered dopaminergic pathways and therapeutic effects of intranasal dopamine in two distinct mouse models of autism. Mol. Brain 2020, 13, 111. [Google Scholar] [CrossRef]
- Zocchi, A.; Fabbri, D.; Heidbreder, C.A. Aripiprazole increases dopamine but not noradrenaline and serotonin levels in the mouse prefrontal cortex. Neurosci. Lett. 2005, 387, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.; Koprivica, V.; Dunn, R.; Tottori, K.; Kikuchi, T.; Altar, C.A. In vivo effects of aripiprazole on cortical and striatal dopaminergic and serotonergic function. Eur. J. Pharmacol. 2004, 483, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Noland, J.S.; Steven, R.J.; Stone, W.L.; Walden, T.; Sheridan, E.H. Better working memory for non-social targets in infant siblings of children with Autism Spectrum Disorder. Dev. Sci. 2010, 13, 244–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedel, G.; Kang, S.H.; Choi, D.Y.; Platt, B. Scopolamine-induced deficits in social memory in mice: Reversal by donepezil. Behav. Brain Res. 2009, 204, 217–225. [Google Scholar] [CrossRef]
- Kohls, G.; Schulte-Ruther, M.; Nehrkorn, B.; Muller, K.; Fink, G.R.; Kamp-Becker, I.; Herpertz-Dahlmann, B.; Schultz, R.T.; Konrad, K. Reward system dysfunction in autism spectrum disorders. Soc. Cogn. Affect. Neurosci. 2013, 8, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Scott-Van Zeeland, A.A.; Dapretto, M.; Ghahremani, D.G.; Poldrack, R.A.; Bookheimer, S.Y. Reward processing in autism. Autism Res. 2010, 3, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Pandina, G.J.; Bossie, C.A.; Youssef, E.; Zhu, Y.; Dunbar, F. Risperidone improves behavioral symptoms in children with autism in a randomized, double-blind, placebo-controlled trial. J. Autism Dev. Disord. 2007, 37, 367–373. [Google Scholar] [CrossRef]
- Owen, R.; Sikich, L.; Marcus, R.N.; Corey-Lisle, P.; Manos, G.; McQuade, R.D.; Carson, W.H.; Findling, R.L. Aripiprazole in the treatment of irritability in children and adolescents with autistic disorder. Pediatrics 2009, 124, 1533–1540. [Google Scholar] [CrossRef] [Green Version]
- Cervenka, S.; Hedman, E.; Ikoma, Y.; Djurfeldt, D.R.; Rück, C.; Halldin, C.; Lindefors, N. Changes in dopamine D2-receptor binding are associated to symptom reduction after psychotherapy in social anxiety disorder. Transl. Psychiatry 2012, 2, e120. [Google Scholar] [CrossRef] [Green Version]
- Schneier, F.R.; Martinez, D.; Abi-Dargham, A.; Zea-Ponce, Y.; Simpson, H.B.; Liebowitz, M.R.; Laruelle, M. Striatal dopamine D(2) receptor availability in OCD with and without comorbid social anxiety disorder: Preliminary findings. Depress. Anxiety 2008, 25, 1–7. [Google Scholar] [CrossRef]
- Sadek, B.; Saad, A.; Subramanian, D.; Shafiullah, M.; Lazewska, D.; Kiec-Kononowiczc, K. Anticonvulsant and procognitive properties of the non-imidazole histamine H3 receptor antagonist DL77 in male adult rats. Neuropharmacology 2016, 106, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [Green Version]
- Sadek, B.; Saad, A.; Sadeq, A.; Jalal, F.; Stark, H. Histamine H3 receptor as a potential target for cognitive symptoms in neuropsychiatric diseases. Behav. Brain Res. 2016, 312, 415–430. [Google Scholar] [CrossRef]
- Bahi, A.; Schwed, J.S.; Walter, M.; Stark, H.; Sadek, B. Anxiolytic and antidepressant-like activities of the novel and potent non-imidazole histamine H(3) receptor antagonist ST-1283. Drug Des. Dev. Ther. 2014, 8, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Gaikwad, U.; Parle, M. Combination of aripiprazole and ethanol attenuates marble-burying behavior in mice. Acta Pol. Pharm. 2011, 68, 435–440. [Google Scholar] [PubMed]
- Burda, K.; Czubak, A.; Kus, K.; Nowakowska, E.; Ratajczak, P.; Zin, J. Influence of aripiprazole on the antidepressant, anxiolytic and cognitive functions of rats. Pharmacol. Rep. 2011, 63, 898–907. [Google Scholar] [CrossRef]
- Russo, E.; Citraro, R.; Davoli, A.; Gallelli, L.; Di Paola, E.D.; De Sarro, G. Ameliorating effects of aripiprazole on cognitive functions and depressive-like behavior in a genetic rat model of absence epilepsy and mild-depression comorbidity. Neuropharmacology 2013, 64, 371–379. [Google Scholar] [CrossRef]
- Besson, M.; Belin, D.; McNamara, R.; Theobald, D.E.H.; Castel, A.; Beckett, V.L.; Crittenden, B.M.; Newman, A.H.; Everitt, B.J.; Robbins, T.W.; et al. Dissociable Control of Impulsivity in Rats by Dopamine D2/3 Receptors in the Core and Shell Subregions of the Nucleus Accumbens. Neuropsychopharmacology 2010, 35, 560–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLeon, A.; Patel, N.C.; Crismon, M.L. Aripiprazole: A comprehensive review of its pharmacology, clinical efficacy, and tolerability. Clin. Ther. 2004, 26, 649–666. [Google Scholar] [CrossRef]
- Eissa, N.; Venkatachalam, K.; Jayaprakash, P.; Falkenstein, M.; Dubiel, M.; Frank, A.; Reiner-Link, D.; Stark, H.; Sadek, B. The Multi-Targeting Ligand ST-2223 with Histamine H3 Receptor and Dopamine D2/D3 Receptor Antagonist Properties Mitigates Autism-Like Repetitive Behaviors and Brain Oxidative Stress in Mice. Int. J. Mol. Sci. 2021, 22, 1947. [Google Scholar] [CrossRef] [PubMed]
- Bastaki, S.M.; Abdulrazzaq, Y.M.; Shafiullah, M.; Wiecek, M.; Kiec-Kononowicz, K.; Sadek, B. Anticonvulsant and reproductive toxicological studies of the imidazole-based histamine H3R antagonist 2-18 in mice. Drug Des. Dev. Ther. 2018, 12, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Hackling, A.; Ghosh, R.; Perachon, S.; Mann, A.; Höltje, H.D.; Wermuth, C.G.; Schwartz, J.C.; Sippl, W.; Sokoloff, P.; Stark, H. N-(omega-(4-(2-methoxyphenyl)piperazin-1-yl)alkyl)carboxamides as dopamine D2 and D3 receptor ligands. J. Med. Chem. 2003, 46, 3883–3899. [Google Scholar] [CrossRef]
- Derieux, C.; Leaute, A.; Brugoux, A.; Jaccaz, D.; Terrier, C.; Pin, J.P.; Kniazeff, J.; Le Merrer, J.; Becker, J.A.J. Chronic sodium bromide treatment relieves autistic-like behavioral deficits in three mouse models of autism. Neuropsychopharmacology 2022, 47, 1680–1692. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 ameliorates stereotyped repetitive behavior and neuroinflammmation in sodium valproate induced autism in mice. Chem. Biol. Interact. 2019, 312, 108775. [Google Scholar] [CrossRef]
- Silverman, J.L.; Yang, M.; Lord, C.; Crawley, J.N. Behavioural phenotyping assays for mouse models of autism. Nat. Rev. Neurosci. 2010, 11, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bambini-Junior, V.; Zanatta, G.; Nunes, G.D.F.; de Melo, G.M.; Michels, M.; Fontes-Dutra, M.; Freire, V.N.; Riesgo, R.; Gottfried, C. Resveratrol prevents social deficits in animal model of autism induced by valproic acid. Neurosci. Lett. 2014, 583, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Valvassori, S.S.; Varela, R.B.; Quevedo, J. Chapter 38—Animal Models of Mood Disorders: Focus on Bipolar Disorder and Depression. In Animal Models for the Study of Human Disease, 2nd ed.; Conn, P.M., Ed.; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar] [CrossRef]
- Lucchina, L.; Depino, A.M. Altered Peripheral and Central Inflammatory Responses in a Mouse Model of Autism. Autism Res. 2014, 7, 273–289. [Google Scholar] [CrossRef]
- Kim, J.W.; Seung, H.; Kwon, K.J.; Ko, M.J.; Lee, E.J.; Oh, H.A.; Choi, C.S.; Kim, K.C.; Gonzales, E.L.; You, J.S.; et al. Subchronic treatment of donepezil rescues impaired social, hyperactive, and stereotypic behavior in valproic acid-induced animal model of autism. PLoS ONE 2014, 9, e104927. [Google Scholar] [CrossRef] [Green Version]
- Prut, L.; Belzung, C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: A review. Eur. J. Pharmacol. 2003, 463, 3–33. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ST-2223 | |
|---|---|
| Target Tested | Ki (nM) |
| hH3R a | 4.6 |
| hH1R b | 85.2 |
| hD2R c | 19.8 |
| hD3R c | 2.0 |
| hD1R d | 564 |
| hD5R d | 5064 |
| EeAChE e | 1000 (<60% inhibition) |
| EqBChE f | 1000 (<30% inhibition) |
| BTBR Mice | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| of BTBR vs. B6 | Histamine | Dopamine | Acetylcholine | ||||||||||
| VEH | ST-2223 (5 mg) | ST-2223 (5 mg) + RAM | ARP (1 mg/kg) | VEH | ST-2223 (5 mg) | ST-2223 (5 mg)+ RAM | ARP (1 mg) | VEH | ST-2223 (5 mg) | ST-2223 (5 mg)+ RAM | ARP (1 mg) | ||
| Prefrontal cortex | ↔ | ↔ | ↔ | ↔ | ↓ a | ↑ b | ↑↑ c | ↔ | ↓ a | ↑ b | ↓ c | ↑ b | |
| Cerebellum | ↓ a | ↑ b | ↓ c | ↑ b | ↔ | ↓ b | ↑↑ c | ↓b | ↓ a | ↔ | ↔ | ↑ b | |
| Striatum | ↓ a | ↑ b | ↓ c | ↔ | ↓ a | ↑ b | ↑↑ c | ↔ | ↔ | ↑ b | ↓ c | ↑ b | |
| Hippocampus | ↓ a | ↔ | ↔ | ↑ b | ↑ a | ↔ | ↑↑ c | ↓b | ↓ a | ↑ b | ↓ c | ↔ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eissa, N.; Venkatachalam, K.; Jayaprakash, P.; Yuvaraju, P.; Falkenstein, M.; Stark, H.; Sadek, B. Experimental Studies Indicate That ST-2223, the Antagonist of Histamine H3 and Dopamine D2/D3 Receptors, Restores Social Deficits and Neurotransmission Dysregulation in Mouse Model of Autism. Pharmaceuticals 2022, 15, 929. https://doi.org/10.3390/ph15080929
Eissa N, Venkatachalam K, Jayaprakash P, Yuvaraju P, Falkenstein M, Stark H, Sadek B. Experimental Studies Indicate That ST-2223, the Antagonist of Histamine H3 and Dopamine D2/D3 Receptors, Restores Social Deficits and Neurotransmission Dysregulation in Mouse Model of Autism. Pharmaceuticals. 2022; 15(8):929. https://doi.org/10.3390/ph15080929
Chicago/Turabian StyleEissa, Nermin, Karthikkumar Venkatachalam, Petrilla Jayaprakash, Priya Yuvaraju, Markus Falkenstein, Holger Stark, and Bassem Sadek. 2022. "Experimental Studies Indicate That ST-2223, the Antagonist of Histamine H3 and Dopamine D2/D3 Receptors, Restores Social Deficits and Neurotransmission Dysregulation in Mouse Model of Autism" Pharmaceuticals 15, no. 8: 929. https://doi.org/10.3390/ph15080929