
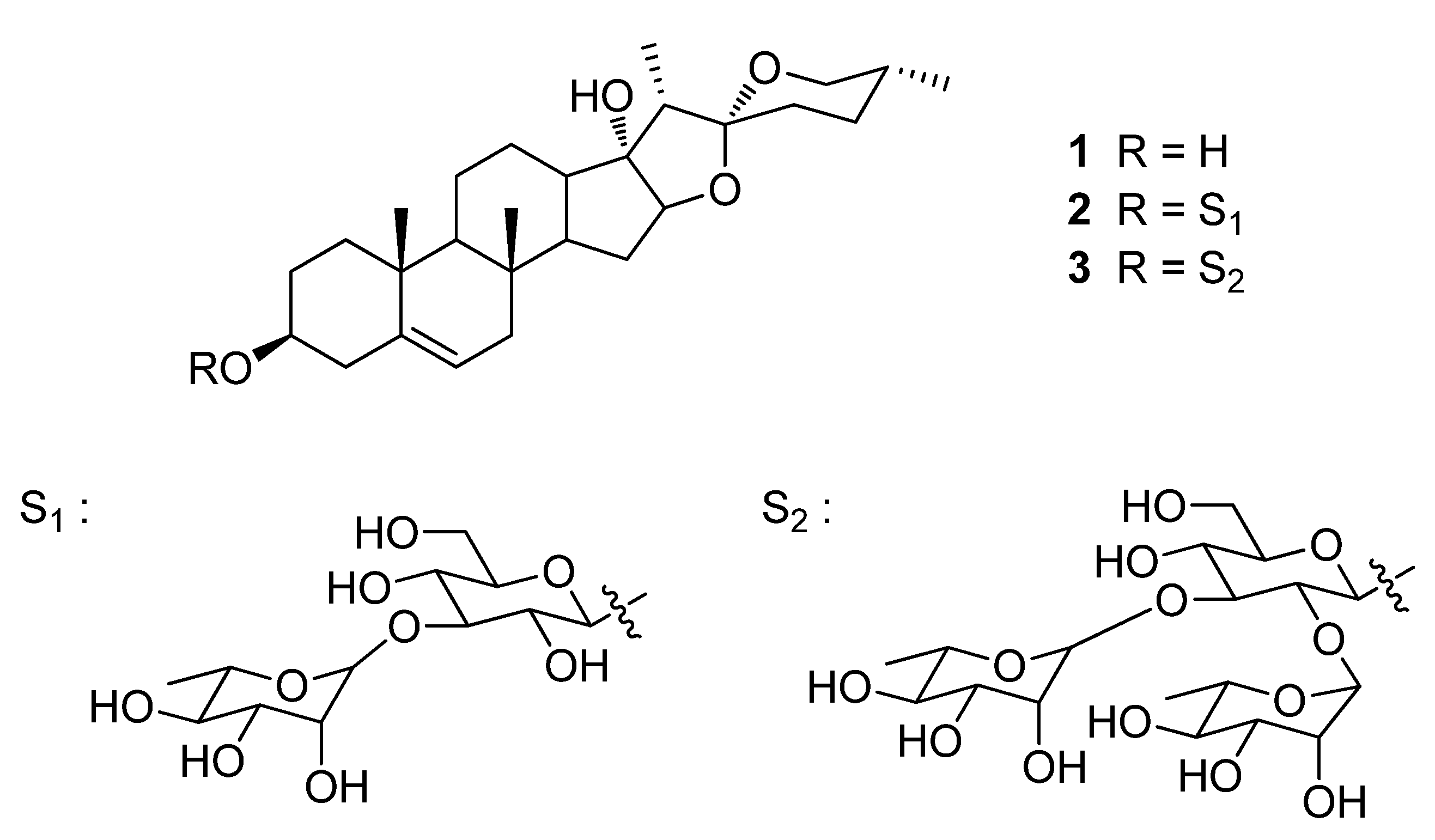
Pennogenin-3-O-α-L-Rhamnopyranosyl-(1→2)-[α-L-Rhamnopyranosyl-(1→3)]-β-D-Glucopyranoside (Spiroconazol A) Isolated from Dioscorea bulbifera L. var. sativa Induces Autophagic Cell Death by p38 MAPK Activation in NSCLC Cells

 and
and
Abstract
:1. Introduction
2. Results
2.1. Spiroconazol A Suppresses the Viability of Various Cancer Cells
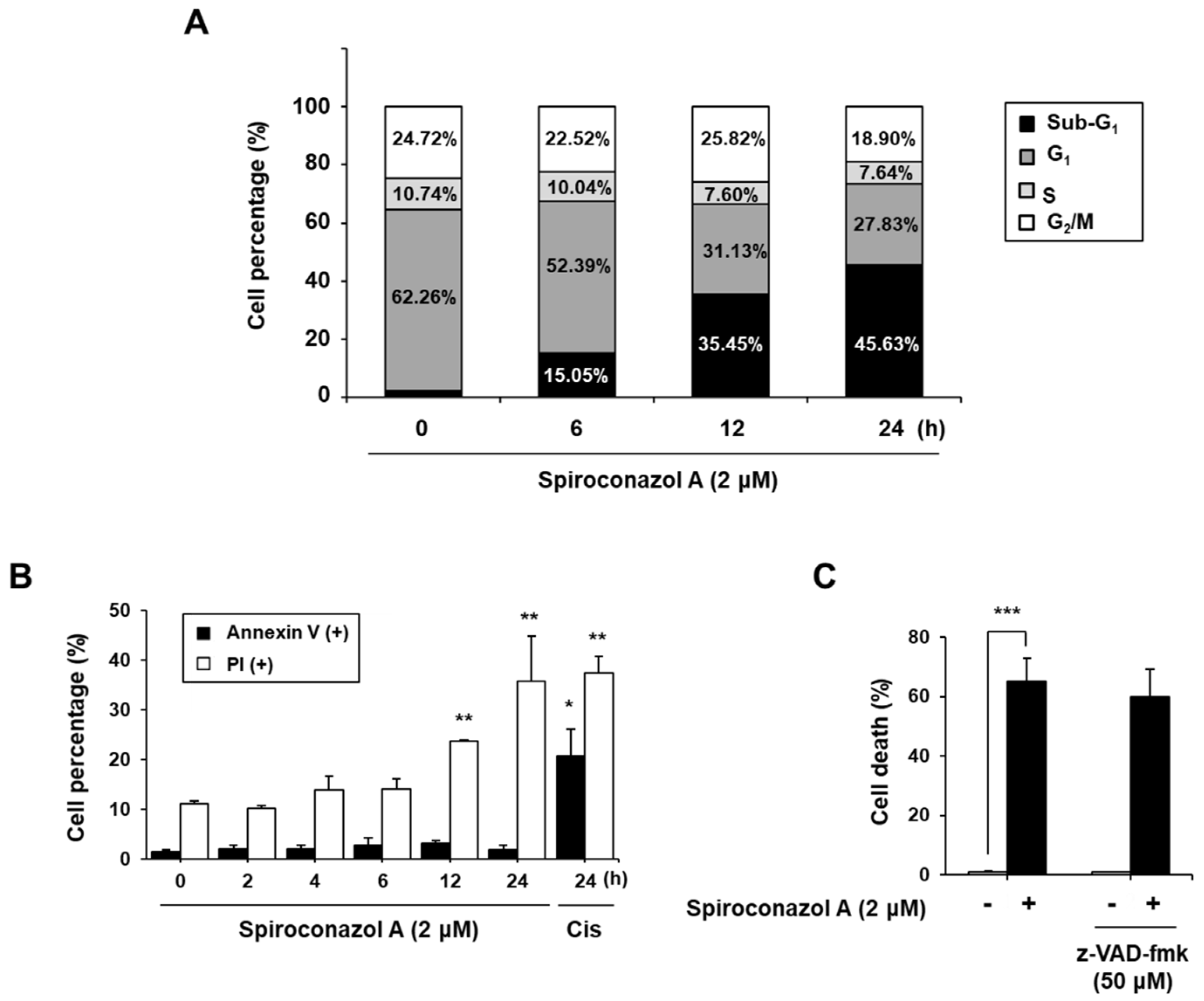
2.2. Spiroconazol A Induces Apoptosis-Independent Cell Death
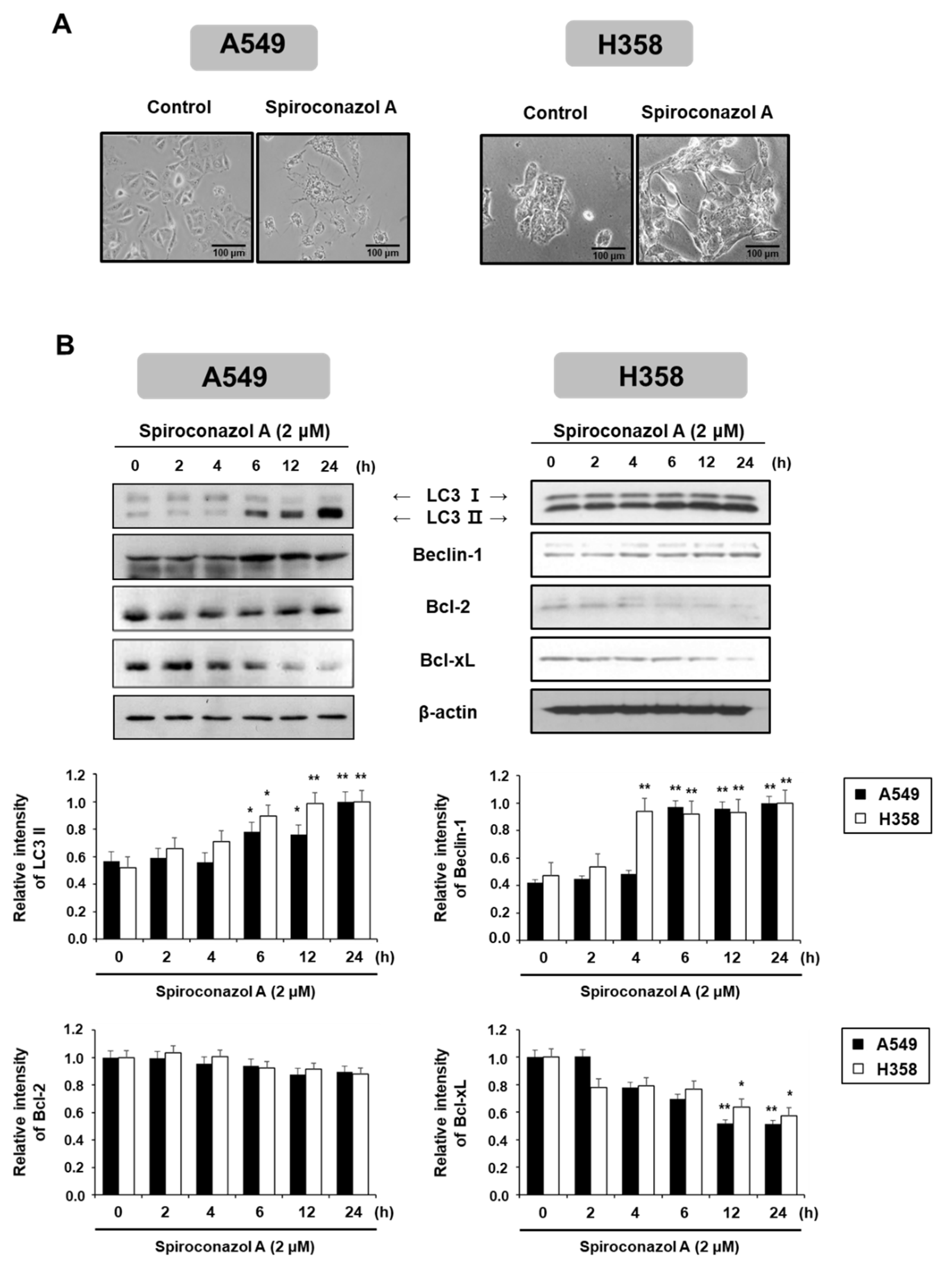
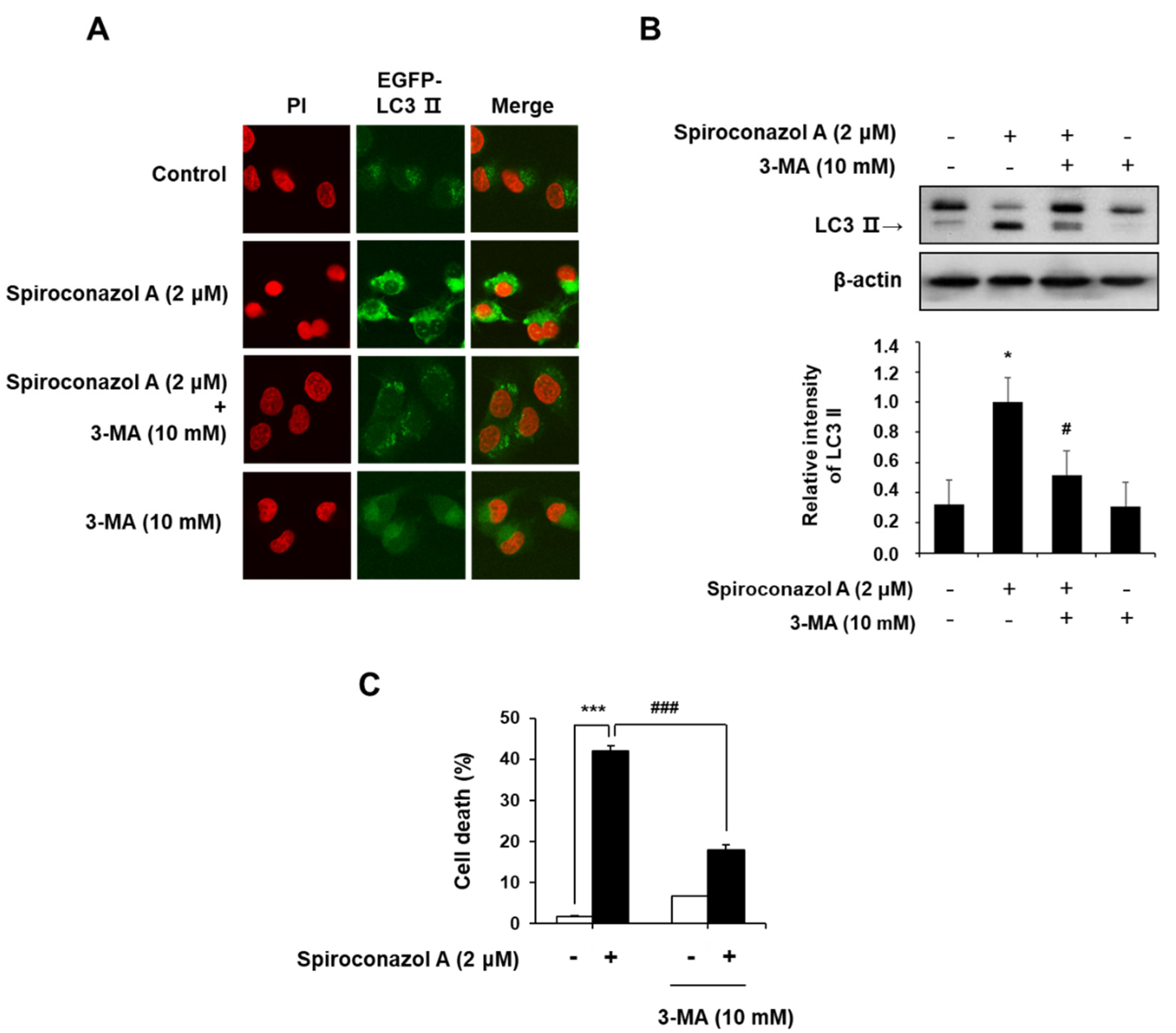
2.3. Spiroconazol A Induces Autophagy in A549 and NCI-H358 Cells
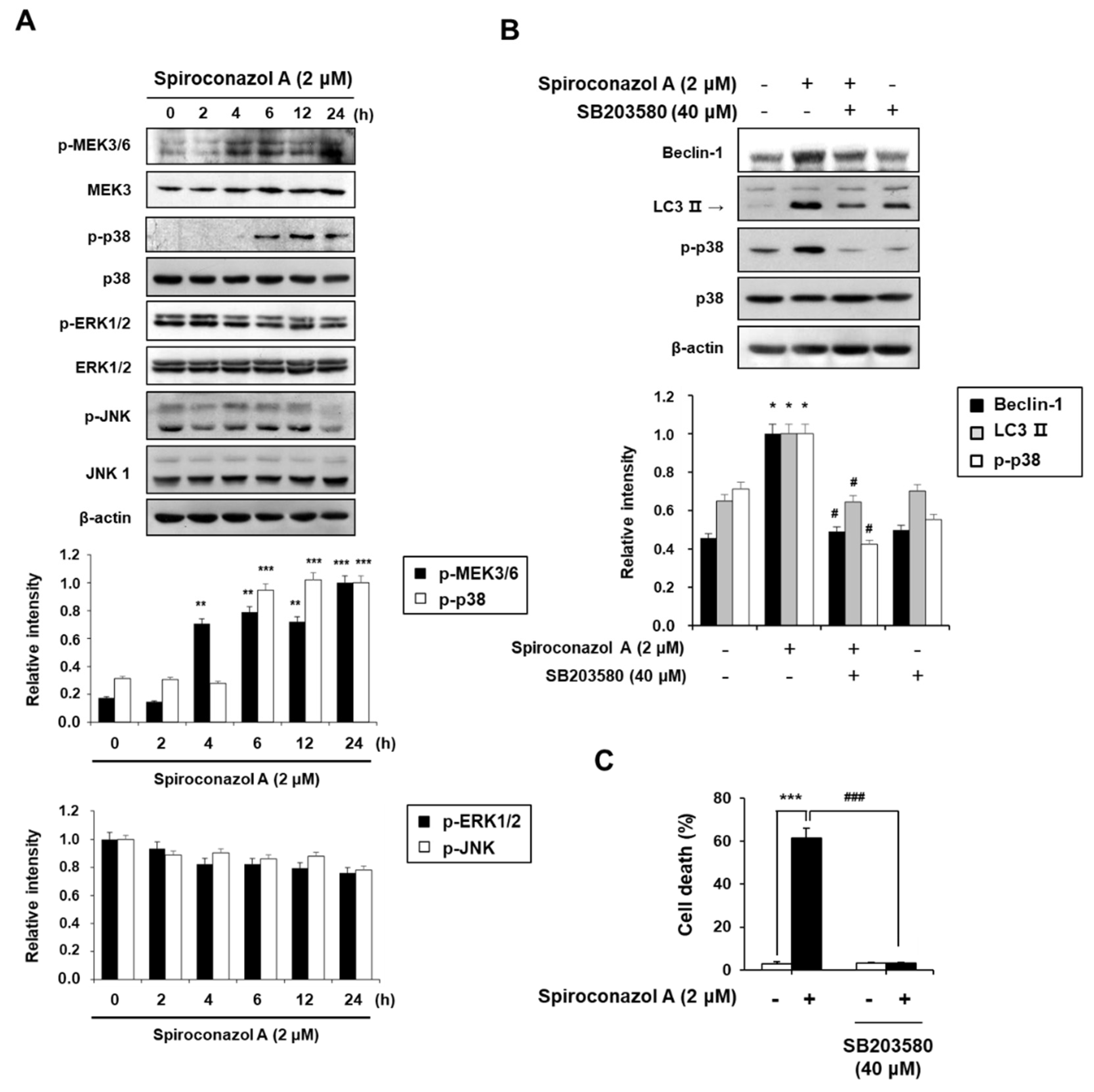
2.4. The p38 MAPK Activation Plays an Important Role in Spiroconazol A-Induced Autophagic Cell Death in A549 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. MTT Assay
4.4. PI Staining Analysis
4.5. Annexin V and PI Double Staining Assay
4.6. Western Blot Analysis
4.7. Construction of pEGFP-LC3-II and Observation with a Confocal Fluorescence Microscope
4.8. Detection of ROS Generation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, Q.; Qu, J.; Sheng, L.; Gao, Q.; Zhou, J. Case Report: Transformation From Non-Small Cell Lung Cancer to Small Cell Lung Cancer During Anti-PD-1 Therapy: A Report of Two Cases. Front. Oncol. 2021, 11, 619371. [Google Scholar] [CrossRef] [PubMed]
- Tungsukruthai, S.; Reamtong, O.; Roytrakul, S.; Sukrong, S.; Vinayanwattikun, C.; Chanvorachote, P. Targeting AKT/mTOR and Bcl-2 for Autophagic and Apoptosis Cell Death in Lung Cancer: Novel Activity of a Polyphenol Compound. Antioxidants 2021, 10, 534. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Chen, M.; Jiang, Z.; Zhao, F.; Xi, B.; Zhang, X.; Fu, H.; Zhou, K. Platycodin-D Induced Autophagy in Non-Small Cell Lung Cancer Cells via PI3K/Akt/mTOR and MAPK Signaling Pathways. J. Cancer 2015, 6, 623–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, M.B.; Chen, Y.; Henson, E.S.; Cizeau, J.; McMillan-Ward, E.; Israels, S.J.; Gibson, S.B. Hypoxia induces autophagic cell death in apoptosis-competent cells through a mechanism involving BNIP3. Autophagy 2008, 4, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanzawa, T.; Kondo, Y.; Ito, H.; Kondo, S.; Germano, I. Induction of autophagic cell death in malignant glioma cells by arsenic trioxide. Cancer Res. 2003, 63, 2103–2108. [Google Scholar] [PubMed]
- Fazi, B.; Bursch, W.; Fimia, G.M.; Nardacci, R.; Piacentini, M.; Di Sano, F.; Piredda, L. Fenretinide induces autophagic cell death in caspase-defective breast cancer cells. Autophagy 2008, 4, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y. Naturally occurring steroidal saponins as potential anticancer agents: Current developments and mechanisms of action. Curr. Top. Med. Chem. 2022; E-pub Ahead of Print. [Google Scholar] [CrossRef]
- He, H.; Sun, Y.-P.; Zheng, L.; Yue, Z.-G. Steroidal Saponins from Paris polyphylla Induce Apoptotic Cell Death and Autophagy in A549 Human Lung Cancer Cells. Asian Pac. J. Cancer Prev. 2015, 16, 1169–1173. [Google Scholar] [CrossRef] [Green Version]
- Yao, M.; Li, R.; Yang, Z.; Ding, Y.; Zhang, W.; Li, W.; Liu, M.; Zhao, C.; Wang, Y.; Tang, H.; et al. PP9, a steroidal saponin, induces G2/M arrest and apoptosis in human colorectal cancer cells by inhibiting the PI3K/Akt/GSK3β pathway. Chem. Interact. 2020, 331, 109246. [Google Scholar] [CrossRef]
- Tapondjou, L.A.; Jenett-Siems, K.; Böttger, S.; Melzig, M.F. Steroidal saponins from the flowers of Dioscorea bulbifera var. sativa. Phytochem. 2013, 95, 341–350. [Google Scholar] [CrossRef]
- Tapondjou, L.A.; Ponou, K.B.; Teponno, R.B.; Mbiantcha, M.; Djoukeng, J.D.; Nguelefack, T.B.; Watcho, P.; Cadenas, A.G.; Park, H.-J. In vivo anti-inflammatory effect of a new steroidal saponin, mannioside A, and its derivatives isolated from Dracaena mannii. Arch. Pharmacal. Res. 2008, 31, 653–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.-J.; Lee, C.O.; Lee, K.-T.; Choi, J.; Park, H.-J. Structure-Activity Relationship of Oleanane Disaccharides Isolated from Akebia quinata versus Cytotoxicity against Cancer Cells and NO Inhibition. Biol. Pharm. Bull. 2004, 27, 744–747. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-J.; Kwon, S.-H.; Lee, J.-H.; Lee, K.-H.; Miyamoto, K.-I.; Lee, K.-T. Kalopanaxsaponin A is a Basic Saponin Structure for the Anti-Tumor Activity of Hederagenin Monodesmosides. Planta Med. 2001, 67, 118–121. [Google Scholar] [CrossRef]
- Prerna, K.; Dubey, V.K. Beclin1-mediated interplay between autophagy and apoptosis: New understanding. Int. J. Biol. Macromol. 2022, 204, 258–273. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, X.; Guo, H.; Zhang, B.; Zhang, X.-B.; Shi, Z.-J.; Yu, L. Synthesis and screening of 3-MA derivatives for autophagy inhibitors. Autophagy 2013, 9, 595–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, S.; Jain, K.; Basu, A. Regulation of Autophagy by Kinases. Cancers 2011, 3, 2630–2654. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Kong, N.; Ye, L.; Han, W.; Zhou, J.; Zhang, Q.; He, C.; Pan, H. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014, 344, 174–179. [Google Scholar] [CrossRef]
- Efridlender, M.; Ekapulnik, Y.; Ekoltai, H. Plant derived substances with anti-cancer activity: From folklore to practice. Front. Plant Sci. 2015, 6, 799. [Google Scholar] [CrossRef]
- Zhu, G.-L.; Hao, Q.; Li, R.-T.; Li, H.-Z. Steroidal saponins from the roots of Asparagus cochinchinensis. Chin. J. Nat. Med. 2014, 12, 213–217. [Google Scholar] [CrossRef]
- Zheng, L.; Zhou, Y.; Zhang, J.-Y.; Song, M.; Yuan, Y.; Xiao, Y.-J.; Xiang, T. Two new steroidal saponins from the rhizomes of Dioscorea zingiberensis. Chin. J. Nat. Med. 2014, 12, 142–147. [Google Scholar] [CrossRef]
- Mirzayans, R.; Murray, D. Do TUNEL and Other Apoptosis Assays Detect Cell Death in Preclinical Studies? Int. J. Mol. Sci. 2020, 21, 9090. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhou, Y.; Cheng, X.; Fan, Y.; He, S.; Li, S.; Ye, H.; Xie, C.; Wu, W.; Li, C.; et al. Isogambogenic acid induces apoptosis-independent autophagic cell death in human non-small-cell lung carcinoma cells. Sci. Rep. 2015, 5, 7697. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Yao, H.; Li, S.; Jin, L.; Shi, P.; Li, Z.; Wang, G.; Lin, S.; Wu, Y.; Li, Y.; et al. Delicaflavone induces autophagic cell death in lung cancer via Akt/mTOR/p70S6K signaling pathway. J. Mol. Med. 2016, 95, 311–322. [Google Scholar] [CrossRef]
- Wang, S.; Xu, X.; Hu, Y.; Lei, T.; Liu, T. Sotetsuflavone Induces Autophagy in Non-Small Cell Lung Cancer Through Blocking PI3K/Akt/mTOR Signaling Pathway in Vivo and in Vitro. Front. Pharmacol. 2019, 10, 1460. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Wang, Z.; Lu, D.; Huang, J.; Liu, J.; Hong, L. Paeonol induces cytoprotective autophagy via blocking the Akt/mTOR pathway in ovarian cancer cells. Cell Death Dis. 2019, 10, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Feng, Y.; Zhu, M.; Siu, F.-M.; Ng, K.-M.; Che, C.-M. A novel mechanism of XIAP degradation induced by timosaponin AIII in hepatocellular carcinoma. Biochim. Biophys. Acta 2013, 1833, 2890–2899. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.K.; Pyee, Y.; Chung, H.-J.; Park, H.J.; Hong, J.-Y.; Son, K.H.; Lee, S.K. Antitumor Activity of Spicatoside A by Modulation of Autophagy and Apoptosis in Human Colorectal Cancer Cells. J. Nat. Prod. 2016, 79, 1097–1104. [Google Scholar] [CrossRef]
- Chen, M.; DU, Y.; Qui, M.; Wang, M.; Chen, K.; Huang, Z.; Jiang, M.; Xiong, F.; Chen, J.; Zhou, J.; et al. Ophiopogonin B-induced autophagy in non-small cell lung cancer cells via inhibition of the PI3K/Akt signaling pathway. Oncol. Rep. 2013, 29, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Pattanaik, A.K.; Das, J.; Bhutia, S.K.; Behera, B.; Singh, P.; Maiti, T.K. Role of PI3K/Akt/mTOR and MEK/ERK pathway in Concanavalin A induced autophagy in HeLa cells. Chem. Interact. 2014, 210, 96–102. [Google Scholar] [CrossRef]
- Plotnikov, A.; Zehorai, E.; Procaccia, S.; Seger, R. The MAPK cascades: Signaling components, nuclear roles and mechanisms of nuclear translocation. Biochim. Biophys. Acta 2011, 1813, 1619–1633. [Google Scholar] [CrossRef] [Green Version]
- Cagnol, S.; Chambard, J.C. ERK and cell death: Mechanisms of ERK-induced cell death—Apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.F.; Nebreda, Á.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Liang, H.; Qin, Z.-H.; Liu, C.-Y. Crotoxin induces apoptosis and autophagy in human lung carcinoma cells in vitro via activation of the p38MAPK signaling pathway. Acta Pharmacol. Sin. 2014, 35, 1323–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akpinar, H.A.; Kahraman, H.; Yaman, I. Ochratoxin A Sequentially Activates Autophagy and the Ubiquitin-Proteasome System. Toxins 2019, 11, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Giordano, S.; Zhang, J. Autophagy, mitochondria and oxidative stress: Cross-talk and redox signalling. Biochem. J. 2012, 441, 523–540. [Google Scholar] [CrossRef] [Green Version]
- Bazhin, A.V.; Philippov, P.P.; Karakhanova, S. Reactive Oxygen Species in Cancer Biology and Anticancer Therapy. Oxid. Med. Cell. Longev. 2016, 2016, 4197815. [Google Scholar] [CrossRef]
- Fan, J.; Ren, D.; Wang, J.; Liu, X.; Zhang, H.; Wu, M.; Yang, G. Bruceine D induces lung cancer cell apoptosis and autophagy via the ROS/MAPK signaling pathway in vitro and in vivo. Cell Death Dis. 2020, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Sun, X.; Wu, Z.; Yuan, H.; Han, H.; Huang, H.; Shu, Y.; Xu, M.; Gao, R.; Li, S.; et al. A Novel Benzofuran Derivative Moracin N Induces Autophagy and Apoptosis Through ROS Generation in Lung Cancer. Front. Pharmacol. 2020, 11, 391. [Google Scholar] [CrossRef]
- Tang, Z.-H.; Cao, W.-X.; Su, M.-X.; Chen, X.; Lu, J.-J. Osimertinib induces autophagy and apoptosis via reactive oxygen species generation in non-small cell lung cancer cells. Toxicol. Appl. Pharmacol. 2017, 321, 18–26. [Google Scholar] [CrossRef]
- Gutiérrez-Uzquiza, Á.; Arechederra, M.; Bragado, P.; Aguirre-Ghiso, J.A.; Porras, A. p38α Mediates Cell Survival in Response to Oxidative Stress via Induction of Antioxidant Genes: Effect on the p70S6K pathway. J. Biol. Chem. 2012, 287, 2632–2642. [Google Scholar] [CrossRef] [Green Version]
- Gil, H.-S.; Lee, J.-H.; Farag, A.K.; Hassan, A.H.E.; Chung, K.-S.; Choi, J.-H.; Roh, E.-J.; Lee, K.-T. AKF-D52, a Synthetic Phenoxypyrimidine-Urea Derivative, Triggers Extrinsic/Intrinsic Apoptosis and Cytoprotective Autophagy in Human Non-Small Cell Lung Cancer Cells. Cancers 2021, 13, 5849. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Chung, K.-S.; Lee, H.-H.; Ko, D.; Kang, M.; Yoo, H.; Ahn, J.; Lee, J.Y.; Lee, K.-T. Improved tumor-suppressive effect of OZ-001 combined with cisplatin mediated by mTOR/p70S6K and STAT3 inactivation in A549 human lung cancer cells. Biomed. Pharmacother. 2021, 142, 111961. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. Point mutations close to the AUG initiator codon affect the efficiency of translation of rat preproinsulin in vivo. Nature 1984, 308, 241–246. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Origin | IC50 (μM) (a) | ||
|---|---|---|---|---|
| Pennogenin | Mannioside A | Spiroconazol A | ||
| A549 | Human lung adenocarcinoma | <100 | 45.77 ± 7.59 | 2.07 ± 0.18 |
| NCI-H358 | Human bronchioalveolar carcinoma | <100 | 52.39 ± 5.30 | 2.17 ± 0.08 |
| HeLa | Human cervix adenocarcinoma | <100 | 69.09 ± 4.92 | 8.21 ± 2.46 |
| Caski | Human cervix epidermoid carcinoma | <100 | 54.91 ± 12.92 | 10.95 ± 0.62 |
| HT-29 | Human colorectal adenocarcinoma | <100 | 34.24 ± 2.73 | 4.94 ± 0.73 |
| HCT-116 | Human colorectal carcinoma | <100 | 68.33 ± 3.94 | 2.55 ± 0.05 |
| ASPC-1 | Human pancreatic adenocarcinoma | <100 | 55.59 ± 7.85 | 3.54 ± 0.75 |
| MiaPaCa-2 | Pancreatic ductal adenocarcinoma | <100 | 60.09 ± 7.77 | 2.18 ± 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ki, Y.S.; Chung, K.-S.; Lee, H.-W.; Choi, J.-H.; Tapondjou, L.A.; Jang, E.; Lee, K.-T. Pennogenin-3-O-α-L-Rhamnopyranosyl-(1→2)-[α-L-Rhamnopyranosyl-(1→3)]-β-D-Glucopyranoside (Spiroconazol A) Isolated from Dioscorea bulbifera L. var. sativa Induces Autophagic Cell Death by p38 MAPK Activation in NSCLC Cells. Pharmaceuticals 2022, 15, 893. https://doi.org/10.3390/ph15070893
Ki YS, Chung K-S, Lee H-W, Choi J-H, Tapondjou LA, Jang E, Lee K-T. Pennogenin-3-O-α-L-Rhamnopyranosyl-(1→2)-[α-L-Rhamnopyranosyl-(1→3)]-β-D-Glucopyranoside (Spiroconazol A) Isolated from Dioscorea bulbifera L. var. sativa Induces Autophagic Cell Death by p38 MAPK Activation in NSCLC Cells. Pharmaceuticals. 2022; 15(7):893. https://doi.org/10.3390/ph15070893
Chicago/Turabian StyleKi, Yo Sook, Kyung-Sook Chung, Heon-Woo Lee, Jung-Hye Choi, Léon Azefack Tapondjou, Eungyeong Jang, and Kyung-Tae Lee. 2022. "Pennogenin-3-O-α-L-Rhamnopyranosyl-(1→2)-[α-L-Rhamnopyranosyl-(1→3)]-β-D-Glucopyranoside (Spiroconazol A) Isolated from Dioscorea bulbifera L. var. sativa Induces Autophagic Cell Death by p38 MAPK Activation in NSCLC Cells" Pharmaceuticals 15, no. 7: 893. https://doi.org/10.3390/ph15070893