
The Evolution of Pharmacological Activities Bouea macrophylla Griffith In Vivo and In Vitro Study: A Review


Abstract
:
1. Introduction
2. Taxonomical Classification
- Kingdom: Plantae
- Division: Magnoliophyta
- Class: Magnolipsida
- Order: Sapindales
- Family: Anacardiaceae
- Genus: Bouea
- Species: Bouea macrophylla (Griffith)
3. Nutrient Content of Bouea macrophylla Griffith
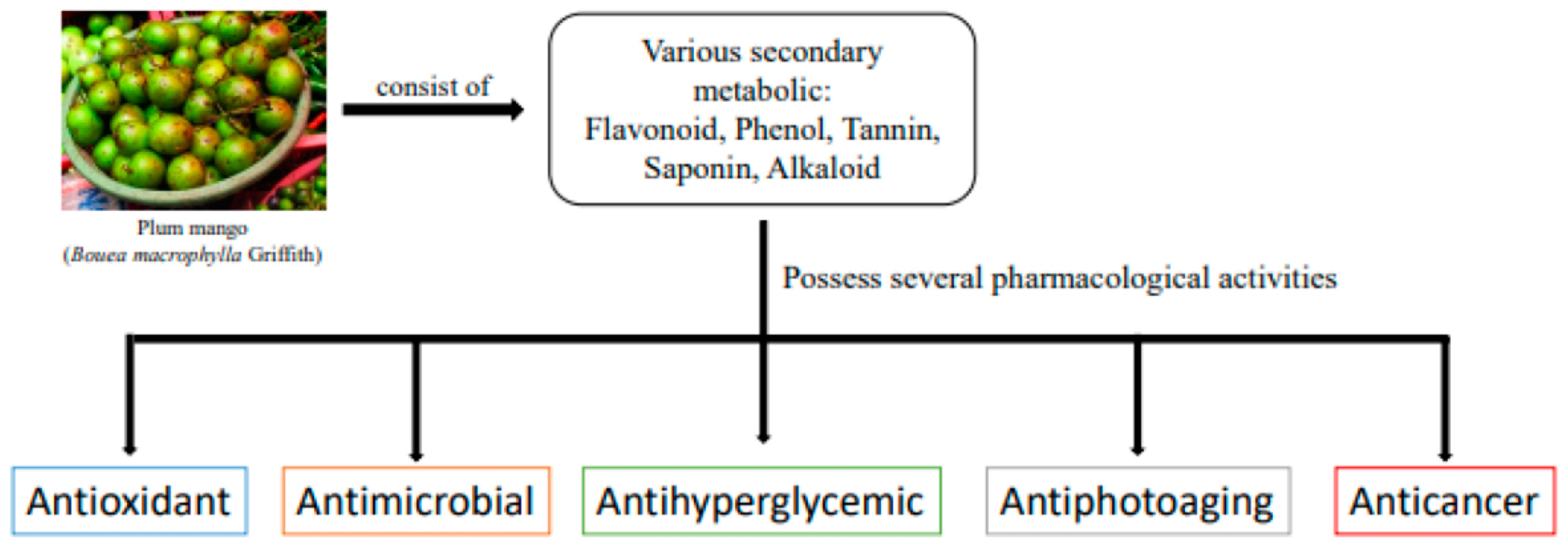
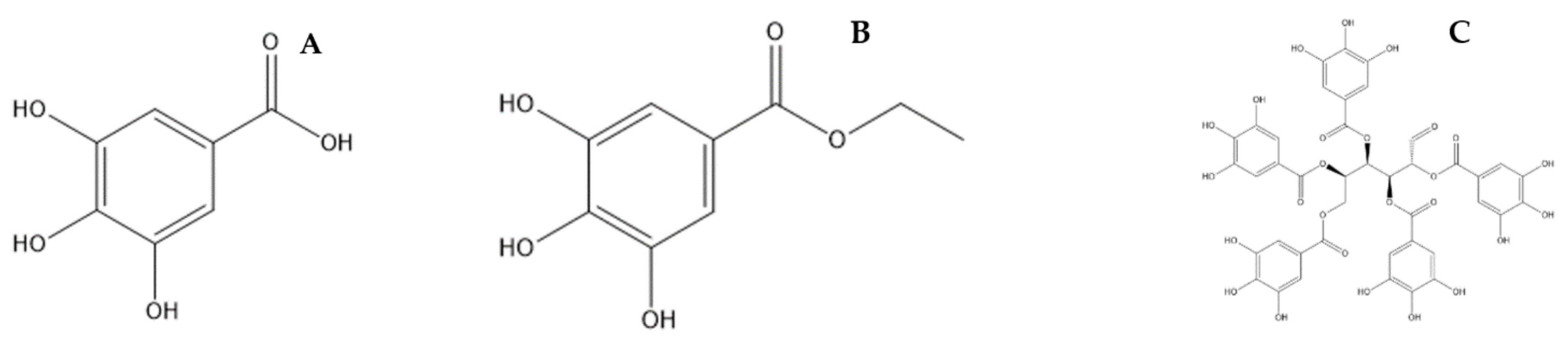
4. Phytochemical Constituents of Bouea macrophylla Griffith
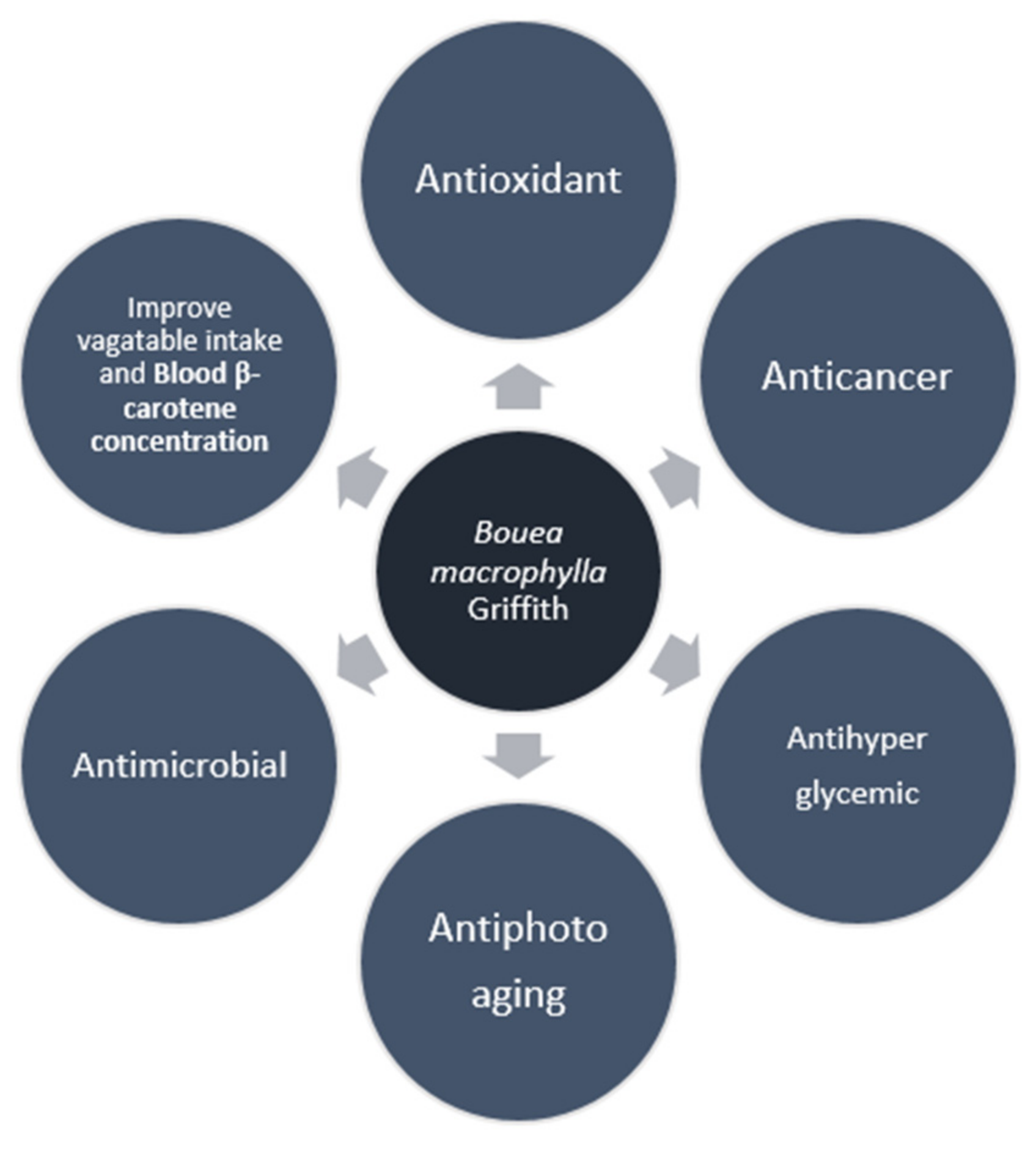
5. Pharmacological Activities of Bouea macrophylla Griffith
5.1. Antioxidant
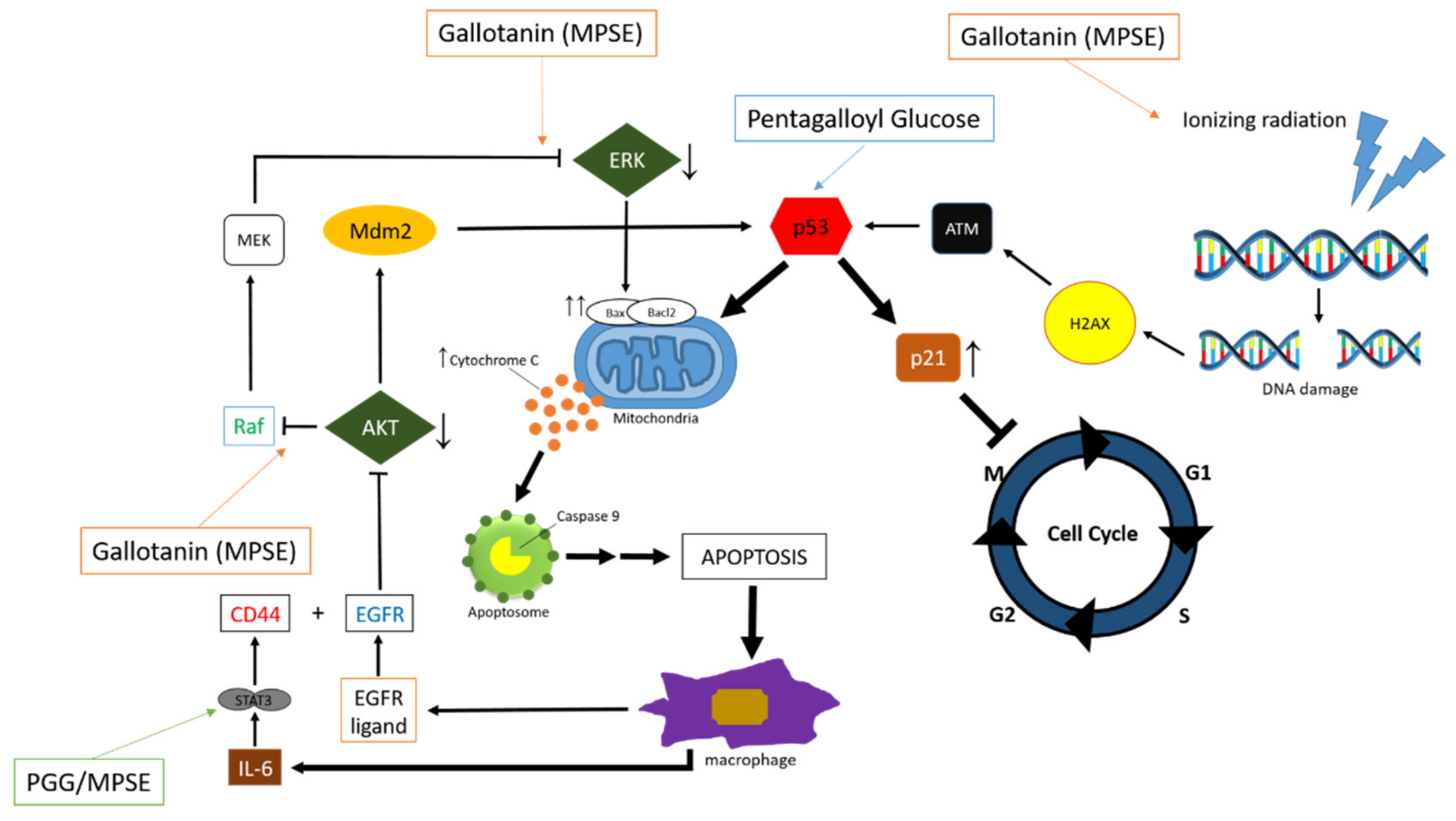
5.2. Anticancer
5.3. Antimicrobial
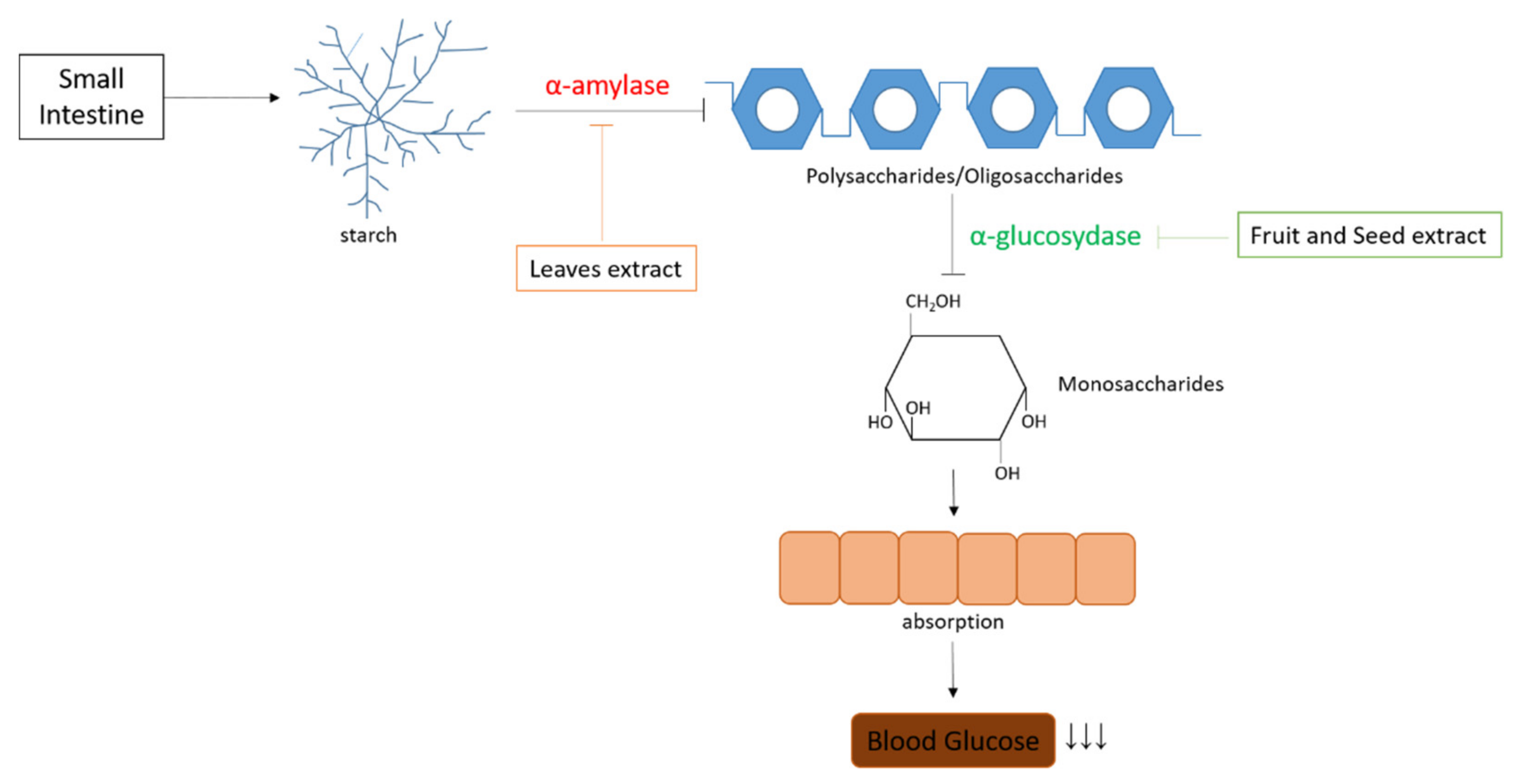
5.4. Antihyperglycemic
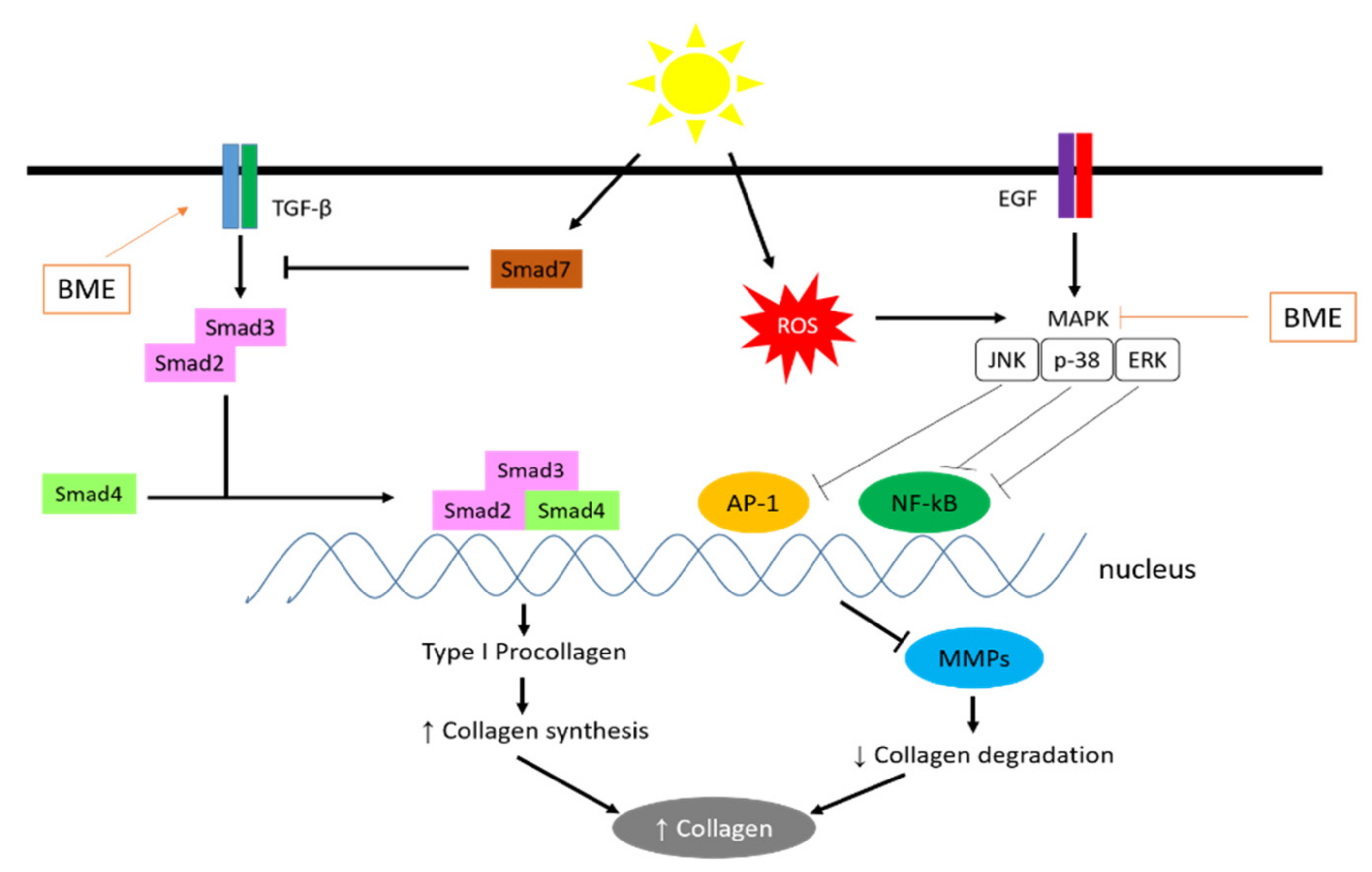
5.5. Antiphotoaging
6. Improve Vegetable Intake and Blood β-Carotene Concentration
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lawalata, V.N. The Physicochemical Characteristics of Gandaria (Bouea macrophylla) Leather with Sugar Concentration Treatment. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 883, p. 012086. [Google Scholar] [CrossRef]
- Shanmuga Rajan, N.; Bhat, R. Antioxidant Compounds and Antioxidant Activities in Unripe and Ripe Kundang Fruits (Bouea macrophylla Griffith). Fruits 2016, 71, 41–47. [Google Scholar] [CrossRef]
- Dechsupa, N.; Kantapan, J.; Tungjai, M.; Intorasoot, S. Maprang “Bouea macrophylla Griffith” Seeds: Proximate Composition, HPLC Fingerprint, and Antioxidation, Anticancer and Antimicrobial Properties of Ethanolic Seed Extracts. Heliyon 2019, 5, e02052. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.K. Edible Medicinal and Non-Medicinal Plants. Edible Med. Non-Med. Plants 2012, 1, 1–738. [Google Scholar] [CrossRef]
- Rajan, N.S.; Bhat, R. Bioactive Compounds of Plum Mango (Bouea macrophylla Griffith). In Bioactive Compounds in Underutilized Fruits and Nuts; Murthy, H.N., Bapat, V.A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 529–541. ISBN 978-3-030-30182-8. [Google Scholar]
- Rajan, N.S.; Bhat, R.; Karim, A. Preliminary Studies on the Evaluation of Nutritional Composition of Unripe and Ripe ‘Kundang’ Fruits (Bouea macrophylla Griffith). Int. Food Res. J. 2014, 21, 985–990. [Google Scholar]
- Nazifi, S.; Saeb, M.; Sepehrimanesh, M.; Poorgonabadi, S. The Effects of Wild Pistachio Oil on Serum Leptin, Thyroid Hormones, and Lipid Profile in Female Rats with Experimental Hypothyroidism. Comp. Clin. Path. 2012, 21, 851–857. [Google Scholar] [CrossRef]
- Lolaen, L.A.C.; Fatimawali, F.; Citraningtyas, G. Uji Aktivitas Antioksidan Kandungan Fitokimia Jus Buah Gandaria (Bouea Macrophylla Griffith). J. Ilm. Farm. 2013, 2, 1–8. [Google Scholar]
- Hanifa, D.; Susilawati, Y. Potensi Tanaman Gandaria (Bouea macrophylla Griff) Sebagai Obat Herbal Yang Beraktivitas Antioksidan. Farmaka 2017, 15, 134–142. [Google Scholar]
- Shah, K.; Patel, M.; Patel, R.; Parmar, P. Mangifera Indica (Mango). Pharmacogn. Rev. 2010, 4, 42–48. [Google Scholar] [CrossRef]
- Alsamri, H.; Athamneh, K.; Pintus, G.; Eid, A.H.; Iratni, R. Pharmacological and Antioxidant Activities of Rhus Coriaria L. (Sumac). Antioxidants 2021, 10, 73. [Google Scholar] [CrossRef]
- Fuadah, I.T.; Wilar, G.; Sumiwi, S.A. A Review of the Pharmacological Effects of Anacardiaceae Family on Controlling Lipid Profile (Dyslipidemia). J. Adv. Pharm. Educ. Res. 2021, 11, 50–58. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Baba, S.; Chan, H.T.; Kainuma, M.; Inoue, T.; Wong, S.K. Ulam Herbs: A Review on the Medicinal Properties of Anacardium Occidentale and Barringtonia Racemosa. J. Appl. Pharm. Sci. 2017, 7, 241–247. [Google Scholar] [CrossRef]
- de Sousa Leite, A.; Islam, M.T.; Júnior, A.L.; e Sousa, J.M.; de Alencar, M.V.; Paz, M.F.; Rolim, H.M.; de Medeiros, M.D.; de Carvalho Melo-Cavalcante, A.A.; Lopes, J.A. Pharmacological Properties of Cashew (Anacardium Occidentale). Afr. J. Biotechnol. 2016, 15, 1855–1863. [Google Scholar] [CrossRef]
- Suliaman, S.F.; Ooi, L.K. Antioxidant and A-glucosidase Inhibitory Activities of 40 Tropical Juices from Malaysia and Identification of Phenolics from the Bioactive Fruit Juices of Barringtonia Racemosa and Phyllanthus Acidus. Algriculture Food Chem. 2014, 62, 9576–9585. [Google Scholar] [CrossRef] [PubMed]
- Zainah, A.; Hassali, H.A.; Razali, R. Phytochemicals Content, Antioxidant and α-Glucosidase Inhibition Activity of Bouea macrophylla Griff Seed Extract. R&D Semin. 2016 Res. Dev. Semin. 2016, 48, 1–8. [Google Scholar]
- Andina, L.; Musfirah, Y. Total Phenolic Content of Cortex And Leaves of Ramania (Bouea macrophylla Griffith) and Antioxidant Activity Assay by DPPH Method. Rjpbcs 2017, 8, 134–140. [Google Scholar]
- Thummajitsakul, S.; Silprasit, K. Genetic Differentiation and Antioxidant Activities of Bouea macrophylla Griffith in Nakhon Nayok Province. J. Appl. Biol. Chem. 2017, 60, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Sukalingam, K. Preliminary Phytochemical Analysis and In Vitro Antioxidant Properties of Malaysian ‘Kundang’ (Bouea macrophylla Griffith). J. Food Biochem. 2018, 43, 261–266. [Google Scholar]
- Wahyuni, L.E.T.; Hardinsyah, H.; Setiawan, B. In-Vitro Alpha Amylase Inhibition and Antioxidant Activities of Leaves Extract of Sundanese Traditional Salad (Lalapan) from Indonesia. J. Gizi Pangan 2020, 15, 109–118. [Google Scholar] [CrossRef]
- Hardinsyah, H.; Windardi, I.P.; Aries, M.; Damayanthi, E. Total Phenolic Content, Quercetin, and Antioxidant Activity of Gandaria (Bouea macrophylla Griff.) Leaf Extract at Two Stages of Maturity. J. Gizi Pangan 2019, 14, 61–68. [Google Scholar] [CrossRef]
- Wardani, W.V.; Hardinsyah; Palupi, E.; Aries, M. Effect of Extraction Technique on Antioxidant Capacity, Vitamin C, Total Phenol, and Total Flavonoid of Bouea macrophylla Griff. Leaf. In IOP Conference Series: Materials Science and Engineering; IOP Publishing: Bristol, UK, 2019; Volume 546. [Google Scholar]
- Fitri, L.; Taufiqurrahman, I.; DH, I. Phytochemical and Cytotoxicity Testing of Ramania Leaves (Bouea macrophylla Griffith) Ethanol Extract toward Vero Cells Using MTT Assay Method (Preliminary Study of Adjuvant Therapy Materials to the Preparation of the Drug). J. Kedokt. Gigi 2018, III, 51–56. [Google Scholar]
- Paksee, S.; Kantapan, J.; Chawapun, P.; Sangthong, P.; Dechsupa, N. Maprang Seed Extracts Suppressed Chemoresistant Properties of Breast Cancer Cells Survived from Ionizing Radiation Treatment via the Regulation of ABCB1 Genes. J. Assoc. Med. Sci. 2019, 52, 185–192. [Google Scholar]
- Kantapan, J.; Paksee, S.; Chawapun, P.; Sangthong, P.; Dechsupa, N. Pentagalloyl Glucose- and Ethyl Gallate-Rich Extract from Maprang Seeds Induce Apoptosis in MCF-7 Breast Cancer Cells through Mitochondria-Mediated Pathway. Evid.-Based Complement. Altern. Med. 2020, 2020, 19. [Google Scholar] [CrossRef] [PubMed]
- Kantapan, J.; Dechsupa, N.; Tippanya, D.; Nobnop, W.; Chitapanarux, I. Gallotannin from Bouea macrophylla Seed Extract Suppresses Cancer Stem-like Cells and Radiosensitizes Head and Neck Cancer. Int. J. Mol. Sci. 2021, 22, 9253. [Google Scholar]
- Kantapan, J.; Paksee, S.; Duangya, A.; Sangthong, P.; Roytrakul, S.; Krobthong, S.; Suttana, W.; Dechsupa, N. A Radiosensitizer, Gallotannin-Rich Extract from Bouea macrophylla Seeds, Inhibits Radiation-Induced Epithelial-Mesenchymal Transition in Breast Cancer Cells. BMC Complement. Med. Ther. 2021, 21, 1–19. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Nguyen, T.T.; Ma, P.C.; Ta, Q.T.H.; Duong, T.H.; Vo, V.G. Potential Antimicrobial and Anticancer Activities of An Ethanol Extract from Bouea macrophylla. Molecules 2020, 25, 1996. [Google Scholar] [CrossRef]
- Leelapornpisid, W.; Poomanee, W. Effectiveness of Maprang “Bouea macrophylla Griffith” Seed Extract against Endodontopathogenic Microorganisms in a Multispecies Bacterial-Fungal Biofilm. Chiang Mai Dent. J. 2021, 42, 8. [Google Scholar]
- Cheong, Y.; Kim, C.; Kim, M.B.; Hwang, J.K. The Anti-Photoaging and Moisturizing Effects of Bouea macrophylla Extract in UVB-Irradiated Hairless Mice. Food Sci. Biotechnol. 2018, 27, 147–157. [Google Scholar] [CrossRef]
- Amrinanto, A.H.; Hardinsyah, H.; Palupi, E. The Eating Culture of the Sundanese: Does the Traditional Salad (Lalapan) Improve Vegetable Intake and Blood P-Carotene Concentration? Future Food J. Food Agric. Soc. 2019, 7, 1–10. [Google Scholar] [CrossRef]
- Cronquist, A. An Integrated System of Classification of Flowering Plants; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Michon, G.; Mary, F. Conversion of Traditional Village Gardens and New Economic Strategies of Rural Households in the Area of Bogor, Indonesia. Agrofor. Syst. 1994, 25, 31–58. [Google Scholar] [CrossRef]
- Rajan, N.S.; Bhat, R. Volatile Constituents of Unripe and Ripe Kundang Fruits (Bouea macrophylla Griffith). Int. J. Food Prop. 2017, 20, 1751–1760. [Google Scholar] [CrossRef]
- Xu, Q.; Zhong, T.; Li, H.L. Antioxidant and Free Radical Scavenging Activities of Chitosan Materials. Adv. Mater. Res. 2014, 1002, 81–90. [Google Scholar] [CrossRef]
- Gutowski, M.; Kowalczyk, S. A Study of Free Radical Chemistry: Their Role and Pathophysiological Significance. Acta Biochim. Pol. 2013, 60, 1–16. [Google Scholar]
- Gulcin, İ. Antioxidants and Antioxidant Methods: An Updated Overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruk, J.; Aboul-Enein, H.Y.; Kładna, A.; Bowser, J.E. Oxidative Stress in Biological Systems and Its Relation with Pathophysiological Functions: The Effect of Physical Activity on Cellular Redox Homeostasis. Free Radic. Res. 2019, 53, 497–521. [Google Scholar] [CrossRef]
- Yadav, P.; Malpathak, N. Estimation of Antioxidant Activity and Total Phenol, Flavonoid Content among Natural Populations of Caper (Capparis Moonii, Wight) from Western Ghats Region. Indian J. Pharm. Educ. Res. 2016, 50, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Adam, Z.; Razali, R.; Arapoc, D.J.; Aziz, A.H.A.; Marsiddi, N.A. DPPH Radical Scavenging and Folin-Ciocalteu Assays: Simple and Reliable Methods to Quantify Antioxidant Activity and Total Phenolic Content. In Proceedings of the Nuclear Technical Conventions, Bangi, Malaysia, 1 July 2021; p. 8. Available online: https://www.researchgate.net/figure/DPPH-radical-scavenging-activity-of-evaluated-plants_tbl1_353463146 (accessed on 18 January 2021).
- Gülçin, İ. Antioxidant Properties of Resveratrol: A Structure—Activity Insight. Innov. Food Sci. Emerg. Technol. 2010, 11, 210–218. [Google Scholar] [CrossRef]
- Alisi, C.S.; Asiwe, E.S.; Ene, C.A.; Alisi, P.N. Antioxidant and Free Radical Scavenging Properties of Psidiumguajava Leaf. Futo J. Ser. 2018, 4, 222–234. [Google Scholar]
- Yuniarti, R.; Nadia, S.; Alamanda, A.; Zubir, M.; Syahputra, R.A.; Nizam, M. Characterization, Phytochemical Screenings and Antioxidant Activity Test of Kratom Leaf Ethanol Extract (Mitragyna Speciosa Korth) Using DPPH Method. J. Phys. Conf. Ser. 2020, 1462. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP Decolorization Assay of Antioxidant Capacity Reaction Pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [Green Version]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Afsar, T.; Razak, S.; Shabbir, M.; Khan, M.R. Antioxidant Activity of Polyphenolic Compounds Isolated from Ethyl-Acetate Fraction of Acacia Hydaspica R. Parker. Chem. Cent. J. 2018, 12, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielińska, E.; Karaś, M.; Jakubczyk, A. Antioxidant Activity of Predigested Protein Obtained from a Range of Farmed Edible Insects. Int. J. Food Sci. Technol. 2017, 52, 306–312. [Google Scholar] [CrossRef]
- Nunes, N.; Valente, S.; Ferraz, S.; Barreto, M.C.; de Carvalho, M.P. Nutraceutical Potential of Asparagopsis taxiformis (Delile) Trevisan Extracts and Assessment of a Downstream Purification Strategy. Heliyon 2018, 4, e00957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, A.; Jelodar, G.; Nazifi, S.; Sajedianfard, J. An Overview of the Characteristics and Function of Vitamin C in Various Tissues: Relying on Its Antioxidant Function. Zahedan J. Res. Med. Sci. 2016, In Press. [CrossRef] [Green Version]
- Townsend, N.; Nichols, M.; Scarborough, P.; Rayner, M. Cardiovascular Disease in Europe—Epidemiological Update 2015. Eur. Heart J. 2015, 36, 2696–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, F.; Zhao, L.; Zhou, A.; Zhang, B.; Li, A.; Wang, Z.; Han, J. The Advantages of Using Traditional Chinese Medicine as an Adjunctive Therapy in the Whole Course of Cancer Treatment Instead of Only Terminal Stage of Cancer. Biosci. Trends 2015, 9, 16–34. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, J.; Mengesha, A.; Hurisa, B.; Mulugeta, D.; Waldetensai, A.; Tesera, Y.; Gemechu, W.; Bizuneh, A.; Getachaw, D.; Desalegn, A. Sequential Adaptation of Vero Cell Lines in Serum Free Medium for Fixed Rabies Virus Propagation. Int. J. Appl. Sci.-Res. Rev. 2021, 8, 20. [Google Scholar]
- Kavitha, N.; Chen, Y.; Kanwar, J.R.; Sasidharan, S. In Situ Morphological Assessment of Apoptosis Induced by Phaleria macrocarpa (Boerl.) Fruit Ethyl Acetate Fraction (PMEAF) in MDA-MB-231 Cells by Microscopy Observation. Biomed. Pharmacother. 2017, 87, 609–620. [Google Scholar] [CrossRef]
- Vijayarathna, S.; Chen, Y.; Kanwar, J.R.; Sasidharan, S. Standardized Polyalthia Longifolia Leaf Extract (PLME) Inhibits Cell Proliferation and Promotes Apoptosis: The Anti-Cancer Study with Various Microscopy Methods. Biomed. Pharmacother. 2017, 91, 366–377. [Google Scholar] [CrossRef]
- Attene-Ramos, M.S.; Huang, R.; Michael, S.; Witt, K.L.; Richard, A.; Tice, R.R.; Simeonov, A.; Austin, C.P.; Xia, M. Profiling of the Tox21 Chemical Collection for Mitochondrial Function to Identify Compounds That Acutely Decrease Mitochondrial Membrane Potential. Environ. Health Perspect. 2015, 123, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Tarasov, A.I.; Griffiths, E.J.; Rutter, G.A. Regulation of ATP Production by Mitochondrial Ca2+. Cell Calcium 2012, 52, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murali, M.; Carvalho, M.S.; Shivanandappa, T. Oxidative Stress-Mediated Cytotoxicity of Endosulfan Is Causally Linked to the Inhibition of NADH Dehydrogenase and Na+, K+-ATPase in Ehrlich Ascites Tumor Cells. Mol. Cell. Biochem. 2020, 468, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Salehiniya, H.; Ghoncheh, M.; Pournamdar, Z. Incidence and Mortality and Epidemiology of Breast Cancer in the World. Asian Pacific J. Cancer Prev. 2016, 17, 43–46. [Google Scholar]
- McCormack, V.A.; Febvey-Combes, O.; Ginsburg, O.; dos-Santos-Silva, I. Breast Cancer in Women Living with HIV: A First Global Estimate. Int. J. Cancer 2018, 143, 2732–2740. [Google Scholar] [CrossRef] [Green Version]
- Parsons, H.A.; Rhoades, J.; Reed, S.C.; Gydush, G.; Ram, P.; Exman, P.; Xiong, K.; Lo, C.C.; Li, T.; Fleharty, M.; et al. Sensitive Detection of Minimal Residual Disease in Patients Treated for Early-Stage Breast Cancer. Clin. Cancer Res. 2020, 26, 2556–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, S.Y.; Moon, J.Y.; Unno, T.; Cho, S.K. Baicalein Suppresses Stem Cell-like Characteristics in Radio-and Chemoresistant MDA-MB-231 Human Breast Cancer Cells through up-Regulation of IFIT2. Nutrients 2019, 11, 624. [Google Scholar] [CrossRef] [Green Version]
- Spiro, A.S.; Wong, A.; Boucher, A.A.; Arnold, J.C. Enhanced Brain Disposition and Effects of Δ 9-Tetrahydrocannabinol in P-Glycoprotein and Breast Cancer Resistance Protein Knockout Mice. PLoS ONE 2012, 7, e35937. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.A.; Tzeng, D.T.W.; Huang, Y.P.; Lin, C.J.; Lo, U.G.; Wu, C.L.; Lin, H.; Hsieh, J.T.; Tang, C.H.; Lai, C.H. Antrocin Sensitizes Prostate Cancer Cells to Radiotherapy through Inhibiting PI3K/AKT and MAPK Signaling Pathways. Cancers 2019, 11, 34. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, Q.; Shanti, R.M.; Shi, S.; Chang, T.H.; Carrasco, L.; Alawi, F.; Le, A.D. Mesenchymal Stromal Cell-Derived Interleukin-6 Promotes Epithelial–Mesenchymal Transition and Acquisition of Epithelial Stem-Like Cell Properties in Ameloblastoma Epithelial Cells. Stem Cells 2017, 35, 2083–2094. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Graham, P.H.; Hao, J.; Bucci, J.; Cozzi, P.J.; Kearsley, J.H.; Li, Y. Emerging Roles of Radioresistance in Prostate Cancer Metastasis and Radiation Therapy. Cancer Metastasis Rev. 2014, 33, 469–496. [Google Scholar] [CrossRef]
- Aghajani, M.; Mansoori, B.; Mohammadi, A.; Asadzadeh, Z.; Baradaran, B. New Emerging Roles of CD133 in Cancer Stem Cell: Signaling Pathway and MiRNA Regulation. J. Cell. Physiol. 2019, 234, 21642–21661. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.M.; Zhang, J.G.; Zhang, X.; Li, Q. Targeting Cancer Stem Cells for Reversing Therapy Resistance: Mechanism, Signaling, and Prospective Agents. Signal Transduct. Target. Ther. 2021, 6, 1–7. [Google Scholar] [CrossRef]
- Chen, Y.W.; Chen, K.H.; Huang, P.I.; Chen, Y.C.; Chiou, G.Y.; Lo, W.L.; Tseng, L.M.; Hsu, H.S.; Chang, K.W.; Chiou, S.H. Cucurbitacin I Suppressed Stem-like Property and Enhanced Radiation-Induced Apoptosis in Head and Neck Squamous Carcinoma-Derived CD44+ALDH1+ Cells. Mol. Cancer Ther. 2010, 9, 2879–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, D.J.; Capela, J.P.; Feio-Azevedo, R.; Teixeira-Gomes, A.; Bastos, M.D.; Carvalho, F. Mitochondria: Key Players in the Neurotoxic Effects of Amphetamines. Arch. Toxicol. 2015, 89, 1695–1725. [Google Scholar] [CrossRef] [PubMed]
- Bertero, E.; Maack, C. Calcium Signaling and Reactive Oxygen Species in Mitochondria. Circ. Res. 2018, 122, 1460–1478. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ge, X.; Qu, H.; Wang, N.; Zhou, J.; Xu, W.; Xie, J.; Zhou, Y.; Shi, L.; Qin, Z.; et al. Glycyrrhizic Acid Inhibits Proliferation of Gastric Cancer Cells by Inducing Cell Cycle Arrest and Apoptosis. Cancer Manag. Res. 2020, 12, 2853–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Yuan, J.; Du, X.; Wang, M.; Yue, L.; Liu, J. Ethyl Gallate Suppresses Proliferation and Invasion in Human Breast Cancer Cells via Akt-NF-ΚB Signaling. Oncol. Rep. 2015, 33, 1284–1290. [Google Scholar] [CrossRef] [Green Version]
- Fazeela Mahaboob Begum, S.M.; Hemalatha, S. Phytoconstituents from Gelidiella acerosa Induce Apoptosis by Regulating Bax, Bcl2 Expression in A549 Cells. Biocatal. Agric. Biotechnol. 2020, 29, 101757. [Google Scholar] [CrossRef]
- Lee, H.L.; Lin, C.S.; Kao, S.H.; Chou, M.C. Gallic Acid Induces G1 Phase Arrest and Apoptosis of Triple-Negative Breast Cancer Cell MDA-MB-231 via P38 Mitogen-Activated Protein Kinase/P21/P27 Axis. Anticancer Drugs 2017, 28, 1150–1156. [Google Scholar] [CrossRef]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [Green Version]
- Adwas, A.A.; Elsayed, A.; Azab, A.E.; Quwaydir, F.A. Oxidative Stress and Antioxidant Mechanisms in Human Body. J. Appl. Biotechnol. Bioeng. 2019, 6, 43–47. [Google Scholar] [CrossRef]
- Reinhardt, H.C.; Schumacher, B. The P53 Network: Cellular and Systemic DNA Damage Responses in Aging and Cancer. Trends Genet. 2012, 28, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Cao, L.; Wang, Z.R.; Li, Z.; Ma, J. Anti-Cancer Effect of Toosendanin and Its Underlying Mechanisms. J. Asian Nat. Prod. Res. 2019, 21, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Sola, M.; Magrin, C.; Pedrioli, G.; Pinton, S.; Salvadè, A.; Papin, S.; Paganetti, P. Tau Affects P53 Function and Cell Fate during the DNA Damage Response. Commun. Biol. 2020, 3, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Bouarab Chibane, L.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant Antimicrobial Polyphenols as Potential Natural Food Preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef] [Green Version]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Arshad, M.I.; Khurshid, M.; Rasool, M.H.; Nisar, M.A.; Aslam, M.A.; Qamar, M.U. Antibiotic Resistance: A Rundown of a Global Crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar]
- Atolani, O.; Olabiyi, E.T.; Issa, A.A.; Azeez, H.T.; Onoja, E.G.; Ibrahim, S.O.; Zubair, M.F.; Oguntoye, O.S.; Olatunji, G.A. Green Synthesis and Characterisation of Natural Antiseptic Soaps from the Oils of Underutilised Tropical Seed. Sustain. Chem. Pharm. 2016, 4, 32–39. [Google Scholar] [CrossRef]
- Szabo, K.; Cătoi, A.F.; Vodnar, D.C. Bioactive Compounds Extracted from Tomato Processing By-Products as a Source of Valuable Nutrients. Plant Foods Hum. Nutr. 2018, 73, 268–277. [Google Scholar] [CrossRef]
- Soong, Y.Y.; Barlow, P.J. Antioxidant Activity and Phenolic Content of Selected Fruit Seeds. Food Chem. 2004, 88, 411–417. [Google Scholar] [CrossRef]
- Sudjaroen, Y.; Haubner, R.; Würtele, G.; Hull, W.E.; Erben, G.; Spiegelhalder, B.; Changbumrung, S.; Bartsch, H.; Owen, R.W. Isolation and Structure Elucidation of Phenolic Antioxidants from Tamarind (Tamarindus indica L.) Seeds and Pericarp. Food Chem. Toxicol. 2005, 43, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Rangkadilok, N.; Tongchusak, S.; Boonhok, R.; Chaiyaroj, S.C.; Junyaprasert, V.B.; Buajeeb, W.; Akanimanee, J.; Raksasuk, T.; Suddhasthira, T.; Satayavivad, J. In Vitro Antifungal Activities of Longan (Dimocarpus Longan Lour.) Seed Extract. Fitoterapia 2012, 83, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.E.; Vargas, A.; Pérez-Sánchez, T.; Encío, I.J.; Cabello-Olmo, M.; Barajas, M. Human Microbiota Network: Unveiling Potential Crosstalk between the Different Microbiota Ecosystems and Their Role in Health and Disease. Nutrients 2021, 13, 2905. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.; Liken, H.; Odackal, N.J. Diagnosis, Risk Factors, Outcomes, and Evaluation of Invasive Candida Infections; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Qin, Q.; Shen, K. ling Community-Acquired Pneumonia and Its Complications. Indian J. Pediatr. 2015, 82, 745–751. [Google Scholar] [CrossRef]
- Pfleger, A.; Eber, E. Assessment and Causes of Stridor. Paediatr. Respir. Rev. 2016, 18, 64–72. [Google Scholar] [CrossRef]
- Schnupf, P.; Sansonetti, P.J. Shigella Pathogenesis: New Insights through Advanced Methodologies. Bact. Intracell. 2020, 15–39. [Google Scholar] [CrossRef]
- Mattock, E.; Blocker, A.J. How Do the Virulence Factors of Shigella Work Together to Cause Disease? Front. Cell. Infect. Microbiol. 2017, 7, 1–24. [Google Scholar] [CrossRef]
- Griffith, R.W.; Carlson, S.A.; Krull, A.C. Salmonellosis Etiology Epidemiology. In Diseases of Swine, 11th Ed. ed; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 912–925. [Google Scholar]
- Todd, E.C.D.; Notermans, S. Surveillance of Listeriosis and Its Causative Pathogen, Listeria monocytogenes. Food Control 2011, 22, 1484–1490. [Google Scholar] [CrossRef]
- Foxman, B. Urinary Tract Infection Syndromes. Occurrence, Recurrence, Bacteriology, Risk Factors, and Disease Burden. Infect. Dis. Clin. N. Am. 2014, 28, 1–13. [Google Scholar] [CrossRef]
- Henry Oladeinde, B.; Omoregie, R.; Olley, M.; Anunibe, J.A. Urinary Tract Infection in a Rural Community of Nigeria. N. Am. J. Med. Sci. 2011, 3, 75–77. [Google Scholar] [CrossRef]
- Chen, H.; Liu, Y. Teeth; Elsevier Inc.: Amsterdam, The Netherlands, 2014; ISBN 9780123946195. [Google Scholar]
- Swimberghe, R.C.D.; Coenye, T.; De Moor, R.J.G.; Meire, M.A. Biofilm Model Systems for Root Canal Disinfection: A Literature Review. Int. Endod. J. 2019, 52, 604–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.S.; De Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-Resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Getachew, Y.; Hassan, L.; Zakaria, Z.; Abdul Aziz, S. Genetic Variability of Vancomycin-Resistant Enterococcus faecium and Enterococcus faecalis Isolates from Humans, Chickens, and Pigs in Malaysia. Appl. Environ. Microbiol. 2013, 79, 4528–4533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodford, N.; Turton, J.F.; Livermore, D.M. Multiresistant Gram-Negative Bacteria: The Role of High-Risk Clones in the Dissemination of Antibiotic Resistance. FEMS Microbiol. Rev. 2011, 35, 736–755. [Google Scholar] [CrossRef] [Green Version]
- Kabuki, T.; Nakajima, H.; Arai, M.; Ueda, S.; Kuwabara, Y.; Dosako, S. Characterization of Novel Antimicrobial Compounds from Mango (Mangifera indica L.) Kernel Seeds. Food Chem. 2000, 71, 61–66. [Google Scholar] [CrossRef]
- Engels, C.; Knödler, M.; Zhao, Y.Y.; Carle, R.; Gänzle, M.G.; Schieber, A. Antimicrobial Activity of Gallotannins Isolated from Mango (Mangifera indica L.) Kernels. J. Agric. Food Chem. 2009, 57, 7712–7718. [Google Scholar] [CrossRef]
- Jiamboonsri, P.; Pithayanukul, P.; Bavovada, R.; Chomnawang, M.T. The Inhibitory Potential of Thai Mango Seed Kernel Extract against Methicillin-Resistant Staphylococcus aureus. Molecules 2011, 16, 6255–6270. [Google Scholar] [CrossRef] [Green Version]
- Mutua, J.K.; Imathiu, S.; Owino, W. Evaluation of the Proximate Composition, Antioxidant Potential, and Antimicrobial Activity of Mango Seed Kernel Extracts. Food Sci. Nutr. 2017, 5, 349–357. [Google Scholar] [CrossRef]
- Guo, F.; Zi, T.; Liu, L.; Feng, R.; Sun, C. A 1H-NMR Based Metabolomics Study of the Intervention Effect of Mangiferin on Hyperlipidemia Hamsters Induced by a High-Fat Diet. Food Funct. 2017, 8, 2455–2464. [Google Scholar] [CrossRef]
- Hintz, T.; Matthews, K.K.; Di, R. The Use of Plant Antimicrobial Compounds for Food Preservation. Biomed Res. Int. 2015, 2015, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wolf, E.; Abdullah, M.I.; Jones, S.M.; Menezes, K.; Moss, D.M.; Drijfhout, F.P.; Hart, S.R.; Hoskins, C.; Stronach, E.A.; Richardson, A. Dietary Geranylgeraniol Can Limit the Activity of Pitavastatin as a Potential Treatment for Drug-Resistant Ovarian Cancer. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Hiyoshi, T.; Fujiwara, M.; Yao, Z. Postprandial Hyperglycemia and Postprandial Hypertriglyceridemia in Type 2 Diabetes. J. Biomed. Res. 2019, 33, 1–16. [Google Scholar] [CrossRef]
- Vieira, R.; Souto, S.B.; Sánchez-López, E.; Machado, A.L.; Severino, P.; Jose, S.; Santini, A.; Silva, A.M.; Fortuna, A.; García, M.L.; et al. Sugar-Lowering Drugs for Type 2 Diabetes Mellitus and Metabolic Syndrome—Strategies for in Vivo Administration: Part-II. J. Clin. Med. 2019, 8, 1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubczyk, A.; Kara, M.; Urszula, Z.; Szymanowska, U. Identification of Potential Inhibitory Peptides of Enzymes Involved in the Metabolic Syndrome Obtained by Simulated Gastrointestinal Digestion of Fermented Bean (Phaseolus vulgaris L.) Seeds. Food Res. Int. 2017, 100, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Proença, C.; Freitas, M.; Ribeiro, D.; Tomé, S.M.; Oliveira, E.F.T.; Viegas, M.F.; Araújo, A.N.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.A.; et al. Evaluation of a Flavonoids Library for Inhibition of Pancreatic α-Amylase towards a Structure–Activity Relationship. J. Enzyme Inhib. Med. Chem. 2019, 34, 577–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sultana, R.; Alashi, A.M.; Islam, K.; Saifullah, M.; Haque, C.E.; Aluko, R.E. Inhibitory Activities of Polyphenolic Extracts of Bangladeshi Vegetables against α-Amylase. Foods 2020, 9, 844. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Das, S.; De, B. In Vitro Inhibition of Key Enzymes Related to Diabetes by the Aqueous Extracts of Some Fruits of West Bengal, India. Curr. Nutr. Food Sci. 2012, 8, 19–24. [Google Scholar] [CrossRef]
- Oboh, G.; Ademosun, A.O.; Odubanjo, O.V.; Akinbola, I.A. Antioxidative Properties and Inhibition of Key Enzymes Relevant to Type-2 Diabetes and Hypertension by Essential Oils from Black Pepper. Adv. Pharmacol. Sci. 2013, 2013, 6. [Google Scholar] [CrossRef]
- Gangoiti, J.; Corwin, S.F.; Lamothe, L.M.; Vafiadi, C.; Hamaker, B.R.; Dijkhuizen, L. Synthesis of Novel α-Glucans with Potential Health Benefits through Controlled Glucose Release in the Human Gastrointestinal Tract. Crit. Rev. Food Sci. Nutr. 2020, 60, 123–146. [Google Scholar] [CrossRef]
- Ayua, E.O.; Nkhata, S.G.; Namaumbo, S.J.; Kamau, E.H.; Ngoma, T.N.; Aduol, K.O. Polyphenolic Inhibition of Enterocytic Starch Digestion Enzymes and Glucose Transporters for Managing Type 2 Diabetes May Be Reduced in Food Systems. Heliyon 2021, 7, e06245. [Google Scholar] [CrossRef]
- Pires, T.C.; Caleja, C.; Santos-Buelga, C.; Barros, L.; Ferreira, I.C. Vaccinium Myrtillus L. Fruits as a Novel Source of Phenolic Compounds with Health Benefits and Industrial Applications—A Review. Curr. Pharm. Des. 2020, 26, 1917–1928. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Menichini, F. Natural Products as α-Amylase and α-Glucosidase Inhibitors and Their Hypoglycaemic Potential in the Treatment of Diabetes: An Update. Mini-Rev. Med. Chem. 2010, 10, 315–331. [Google Scholar] [CrossRef]
- Samudra, A.; Nugroho, A.; Husni, A. Aktivitas Inhibisi A-Amilase Ekstrak Karagenan Dan Senyawa Polifenol Dari Eucheuma Denticulatum. Media Farm. 2015, 12, 83–92. [Google Scholar] [CrossRef]
- Gu, C.; Zhang, H.; Putri, C.Y.; Ng, K. Evaluation of α-Amylase and α-Glucosidase Inhibitory Activity of Flavonoids. Int. J. Food Nutr. Sci. 2016, 2, 1–6. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-Glucosidase and α-Amylase by Flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, Y.; Miao, M. Inhibition of α-Amylase by Polyphenolic Compounds: Substrate Digestion, Binding Interactions and Nutritional Intervention. Trends Food Sci. Technol. 2020, 104, 190–207. [Google Scholar] [CrossRef]
- Dawadi, S.; Katuwal, S.; Gupta, A.; Lamichhane, U.; Thapa, R.; Jaisi, S.; Lamichhane, G.; Bhattarai, D.P.; Parajuli, N. Current Research on Silver Nanoparticles: Synthesis, Characterization, and Applications. J. Nanomater. 2021, 2021, 23. [Google Scholar] [CrossRef]
- Yuan, E.; Liu, B.; Wei, Q.; Yang, J.; Chen, L.; Li, Q. Structure Activity Relationships of Flavonoids as Potent α-Amylase Inhibitors. Nat. Prod. Commun. 2014, 9, 1173–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adisakwattana, S.; Chanathong, B. Alpha-Glucosidase Inhibitory Activity and Lipid-Lowering Mechanisms of Moringa Oleifera Leaf Extract. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 803–808. [Google Scholar] [PubMed]
- Liu, K.; Luo, M.; Wei, S. The Bioprotective Effects of Polyphenols on Metabolic Syndrome against Oxidative Stress: Evidences and Perspectives. Oxid. Med. Cell. Longev. 2019, 2019, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Manson, J.E.; Buring, J.E.; Sesso, H.D.; Liu, S. Associations of Dietary Flavonoids with Risk of Type 2 Diabetes, and Markers of Insulin Resistance and Systemic Inflammation in Women: A Prospective Study and Cross-Sectional Analysis. J. Am. Coll. Nutr. 2005, 24, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.T.; Lin-Shiau, S.Y.; Shyur, L.F.; Lin, J.K. Pu-Erh Tea Polysaccharides Decrease Blood Sugar by Inhibition of α-Glucosidase Activity in Vitro and in Mice. Food Funct. 2015, 6, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Feng, D.; Wang, T.; Ren, Y.; Liu, Y.; Wang, J. Inhibitors of α-Amylase and α-Glucosidase: Potential Linkage for Whole Cereal Foods on Prevention of Hyperglycemia. Food Sci. Nutr. 2020, 8, 6320–6337. [Google Scholar] [CrossRef]
- Christiansen, S.C.; Fougner, A.L.; Stavdahl, Ø.; Kölle, K.; Ellingsen, R.; Carlsen, S.M. A Review of the Current Challenges Associated with the Development of an Artificial Pancreas by a Double Subcutaneous Approach. Diabetes Ther. 2017, 8, 489–506. [Google Scholar] [CrossRef]
- Karasov, W.H.; Caviedes-Vidal, E. Adaptation of Intestinal Epithelial Hydrolysis and Absorption of Dietary Carbohydrate and Protein in Mammals and Birds. Comp. Biochem. Physiol.-Part A Mol. Integr. Physiol. 2021, 253, 110860. [Google Scholar] [CrossRef]
- Kawee-Ai, A.; Kim, A.T.; Kim, S.M. Inhibitory Activities of Microalgal Fucoxanthin against α-Amylase, α-Glucosidase, and Glucose Oxidase in 3T3-L1 Cells Linked to Type 2 Diabetes. J. Oceanol. Limnol. 2019, 37, 928–937. [Google Scholar] [CrossRef]
- Van Doan, H.; Riyajan, S.; Iyara, R.; Chudapongse, N. Antidiabetic Activity, Glucose Uptake Stimulation and α-Glucosidase Inhibitory Effect of Chrysophyllum cainito L. Stem Bark Extract. BMC Complement. Altern. Med. 2018, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Coniff, R.; Krol, A. Acarbose: A Review of US Clinical Experience. Clin. Ther. 1997, 19, 16–26. [Google Scholar] [CrossRef]
- Lee, S.Y.; Mediani, A.; Nur Ashikin, A.H.; Azliana, A.B.S.; Abas, F. Antioxidant and α-Glucosidase Inhibitory Activities of the Leaf and Stem of Selected Traditional Medicinal Plants. Int. Food Res. J. 2014, 21, 379–386. [Google Scholar]
- Kuhn, A.; Zahn, S.; Patsinakidis, N.; Landmann, A.; Graef, M.; Sauerland, C.; Surber, C.; Wenzel, J. Resistance to Water and Abrasion of a Broad-Spectrum Sunscreen: A Prospective, Open-Label Study. Exp. Dermatol. 2016, 25, 151–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, N.; Ryu, J.J.; Choi, E.H.; Kaushik, N.K. Generation and Role of Reactive Oxygen and Nitrogen Species Induced by Plasma, Lasers, Chemical Agents, and Other Systems in Dentistry. Oxid. Med. Cell. Longev. 2017, 2017, 7542540. [Google Scholar] [CrossRef] [PubMed]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’Nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, J.Y.; Wang, X.; Liu, Y.; Zhang, Z.; Wang, L.; Chen, J.; Huang, S.; Zeng, H.; Lai, X. UV-Induced Skin Photoaging through Inhibiting Oxidative Stress and Inflammation. Mediat. Inflamm. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K. Garlic Supplementation Ameliorates UV-Induced Photoaging in Hairless Mice by Regulating Antioxidative Activity and MMPs Expression. Molecules 2016, 21, 70. [Google Scholar] [CrossRef] [Green Version]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet Radiation and Skin Aging: Roles of Reactive Oxygen Species, Inflammation and Protease Activation, and Strategies for Prevention of Inflammation-Induced Matrix Degradation—A Review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Tanaka, Y.T.; Tanaka, K.; Kojima, H.; Hamada, T.; Masutani, T.; Tsuboi, M.; Akao, Y. Cynaropicrin from Cynara Scolymus L. Suppresses Photoaging of Skin by Inhibiting the Transcription Activity of Nuclear Factor-Kappa B. Bioorg. Med. Chem. Lett. 2013, 23, 518–523. [Google Scholar] [CrossRef]
- Sharma, S.; Dwivedi, S.; Chandra, S.; Srivastava, A.; Vijay, P. Collagen: A Brief Analysis. Oral Maxillofac. Pathol. J. 2019, 10, 10–17. [Google Scholar]
- Breitkreutz, D.; Koxholt, I.; Thiemann, K.; Nischt, R. Skin Basement Membrane: The Foundation of Epidermal Integrity. Hindawi 2013, 2013, 16. [Google Scholar]
- Chen, T.; Hou, H.; Lu, J.; Zhang, K.; Li, B. Protective Effect of Gelatin and Gelatin Hydrolysate from Salmon Skin on UV Irradiation-Induced Photoaging of Mice Skin. J. Ocean Univ. China 2016, 15, 711–718. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix Metalloproteinases and the Regulation of Tissue Remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Denton, C.P.; Zheng, B.; Evans, L.A.; Shi-Wen, X.; Ong, V.H.; Fisher, I.; Lazaridis, K.; Abraham, D.J.; Black, C.M.; De Crombrugghe, B. Fibroblast-Specific Expression of a Kinase-Deficient Type II Transforming Growth Factor β (TGFβ) Receptor Leads to Paradoxical Activation of TGFβ Signaling Pathways with Fibrosis in Transgenic Mice. J. Biol. Chem. 2003, 278, 25109–25119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Y.; Gudey, S.K.; Landström, M. Non-Smad Signaling Pathways. Cell Tissue Res. 2012, 347, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, K.W.; Kim, H.Y.; Oh, M.H.; Byun, S.; Lim, S.H.; Heo, Y.S.; Kang, N.J.; Bode, A.M.; Dong, Z.; et al. Myricetin Suppresses UVB-Induced Wrinkle Formation and MMP-9 Expression by Inhibiting Raf. Biochem. Pharmacol. 2010, 79, 1455–1461. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Jin, M.; Yang, F.; Zhu, J.; Xiao, Q.; Zhang, L. Matrix Metalloproteinases: Inflammatory Regulators of Cell Behaviors in Vascular Formation and Remodeling. Mediat. Inflamm. 2013, 2013, 14. [Google Scholar] [CrossRef] [Green Version]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.C.; Cheng, C.W.; Lee, H.J.; Chu, H.C. Apple Polyphenol Suppresses Indomethacin-Induced Gastric Damage in Experimental Animals by Lowering Oxidative Stress Status and Modulating the MAPK Signaling Pathway. J. Med. Food 2017, 20, 1113–1120. [Google Scholar] [CrossRef]
- Lee, C.W.; Na, Y.; Park, N.H.; Kim, H.S.; Ahn, S.M.; Kim, J.W.; Kim, H.K.; Jang, Y.P. Amentoflavone Inhibits UVB-Induced Matrix Metalloproteinase-1 Expression through the Modulation of AP-1 Components in Normal Human Fibroblasts. Appl. Biochem. Biotechnol. 2012, 166, 1137–1147. [Google Scholar] [CrossRef]
- Oh, Y.; Lim, H.W.; Kim, K.; Lim, C.J. Ginsenoside Re Improves Skin Barrier Function in HaCaT Keratinocytes under Normal Growth Conditions. Biosci. Biotechnol. Biochem. 2016, 80, 2165–2167. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lim, Y.J.; Park, J.H.; Cho, Y. Dietary Silk Protein, Sericin, Improves Epidermal Hydration with Increased Levels of Filaggrins and Free Amino Acids in NC/Nga Mice. Br. J. Nutr. 2012, 108, 1726–1735. [Google Scholar] [CrossRef] [Green Version]
- Souyoul, S.A.; Saussy, K.P.; Lupo, M.P. Nutraceuticals: A Review. Dermatol. Ther. 2018, 8, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Zouboulis, C.C.; Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Elewa, R.; Makrantonaki, E. Aesthetic Aspects of Skin Aging, Prevention, and Local Treatment. Clin. Dermatol. 2019, 37, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Photoprotection by Dietary Carotenoids: Concept, Mechanisms, Evidence and Future Development. Mol. Nutr. Food Res. 2012, 56, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Schagen, S.K.; Zampeli, V.A.; Makrantonaki, E.; Zouboulis, C.C. Discovering the Link between Nutrition and Skin Aging. Dermatoendocrinol 2012, 4, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Meinke, M.C.; Darvin, M.E.; Vollert, H.; Lademann, J. Bioavailability of Natural Carotenoids in Human Skin Compared to Blood. Eur. J. Pharm. Biopharm. 2010, 76, 269–274. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Part | Extraction Technique | Total Phenolic Content | Antioxidant Assay | Antioxidant Activity | Reference | |
|---|---|---|---|---|---|---|
| Seed (unripe and ripe) | Ethanol maceration | 0.77 mg GAE/mg (unripe) | FRAP | 114.98 μg Fe2+E/μg | [3] | |
| TEAC | 2.21 μg TE/μg | |||||
| DPPH | 20.87 µg/mL | |||||
| 0.47 mg GAE/mg (ripe) | FRAP | 94.82 μg Fe2+E/μg | ||||
| TEAC | 1.72 μg TE/μg | |||||
| DPPH | 31.14 µg/mL | |||||
| Seed | Chloroform maceration | 686.04 mg GAE/g | DPPH | 4.34 µg/mL | [40] | |
| Leaves | Ethanol maceration | 530.85 mg GAE/g | ABTS | 1.37 µg/mL | [18] | |
| FIC | 1.70 µg/mL | |||||
| Leaves | Ethanol maceration | 68.53 mg GAE/g | DPPH | 55.83 µg/mL | [17] | |
| Leaves | Vacuum Evaporator extraction | 117.836 mg GAE/g | DPPH | 26 µg/mL | [22] | |
| Leaves | Ethanol maceration | 20 mg GAE/g | FRAP | 5.62 ± 0.38 mg FeSO4 equivalent/g | [21] | |
| Hexane maceration | 30.84 mg GAE/g | 4.5 mg FeSO4 equivalent/g | ||||
| Leaves | Water maceration | 364.56 mg GAE/g | DPPH | 35 µg/mL | [20] | |
| Fruit | Ripe | Aqueous maceration | - | DPPH | 83% | [19] |
| Unripe | 82% | |||||
| Leaves | 76% | |||||
| Fruit | Water maceration | 372.35 µg GAE/g | FRAP | 133.31 μg TEAC/g | [15] | |
| DPPH | 258.17 μg VCEAC/g | |||||
| Fruit | Maceration (methanol, Ethanol, and distilled water) | - | FRAP | 16,290.91 µM Fe(II)/100 g | [2] | |
| DPPH | 77.69% | |||||
| ABTS | 99.76% | |||||
| Plants Part | Extraction Techniques | Anticancer Assay | Cell Lines | IC50 | Reference |
|---|---|---|---|---|---|
| Leaves | Ethanol maceration | MTT assay | Vero Cell | 35.808 µg/mL | [23] |
| Seed | Ethanol maceration | MTT assay | Doxorubicin-sensitive and resistant leukemic(K562, K562/ADR) and lung cancer (GLC4 and GLC4/ADR) cells | 4–16 µg/mL | [3] |
| Seed | Ethanol maceration | MTT assay | MCF-7/IR6 cells | 215.42 nM | [24] |
| Seed | Ethanol maceration | MTT assay | MCF cells | 6.94 µg/mL | [25] |
| Seed | Ethanol maceration | Mammosphere formation assay | Breast cancer stem cells (CSCs) | [27] | |
| Seed | Ethanol maceration | Tumorsphere formation assay, colony formation assay, and apoptosis assay | Head and neck squamous cell carcinoma (HNSCC) | 14.52 µg/mL | [26] |
| Plants Part | Extraction Techniques | Antihyperglycemic Assay | Dose/IC50 | Mechanisms of Action | Reference |
|---|---|---|---|---|---|
| Seed | Decoction | α-Glucosidase Inhibitory Assay | 0.55 mg/mL | Delayed glucose absorption in the small intestine | [40] |
| Leaves | Water maceration | α-Amylase Inhibitory Assay | 60 µg/mL | Inhibit α-Amylase by forming a complex | [20] |
| Fruit | Water maceration | α-Glucosidase Inhibitory Assay | 83.44% | Delayed glucose absorption in the small intestine | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu’adah, I.T.; Sumiwi, S.A.; Wilar, G. The Evolution of Pharmacological Activities Bouea macrophylla Griffith In Vivo and In Vitro Study: A Review. Pharmaceuticals 2022, 15, 238. https://doi.org/10.3390/ph15020238
Fu’adah IT, Sumiwi SA, Wilar G. The Evolution of Pharmacological Activities Bouea macrophylla Griffith In Vivo and In Vitro Study: A Review. Pharmaceuticals. 2022; 15(2):238. https://doi.org/10.3390/ph15020238
Chicago/Turabian StyleFu’adah, Intan Tsamrotul, Sri Adi Sumiwi, and Gofarana Wilar. 2022. "The Evolution of Pharmacological Activities Bouea macrophylla Griffith In Vivo and In Vitro Study: A Review" Pharmaceuticals 15, no. 2: 238. https://doi.org/10.3390/ph15020238