
Anxiolytic-like Effects of the Positive GABAB Receptor Modulator GS39783 Correlate with Mice’s Individual Basal Anxiety and Stress Reactivity

 , ,
, , 
Abstract
:1. Introduction
2. Results
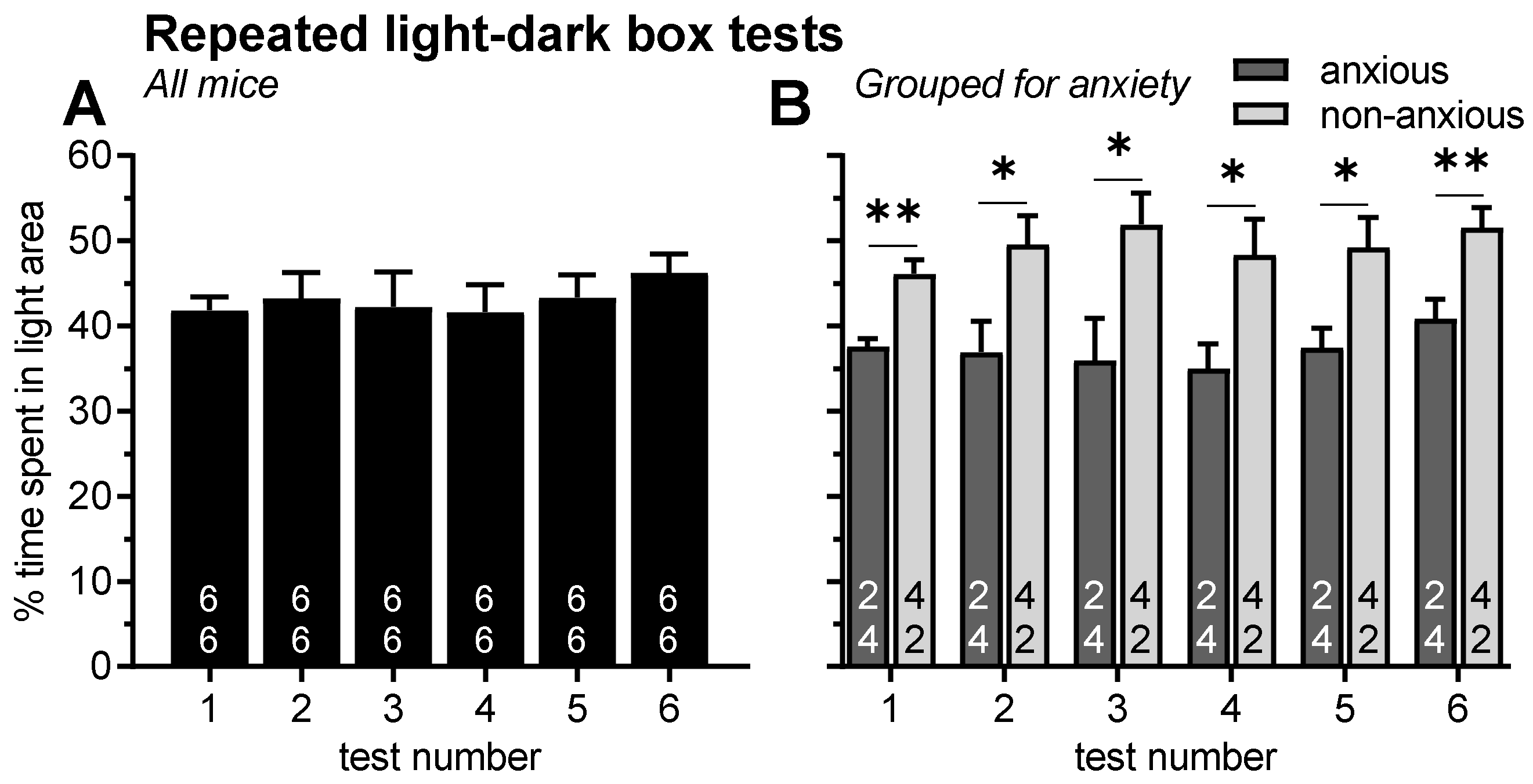
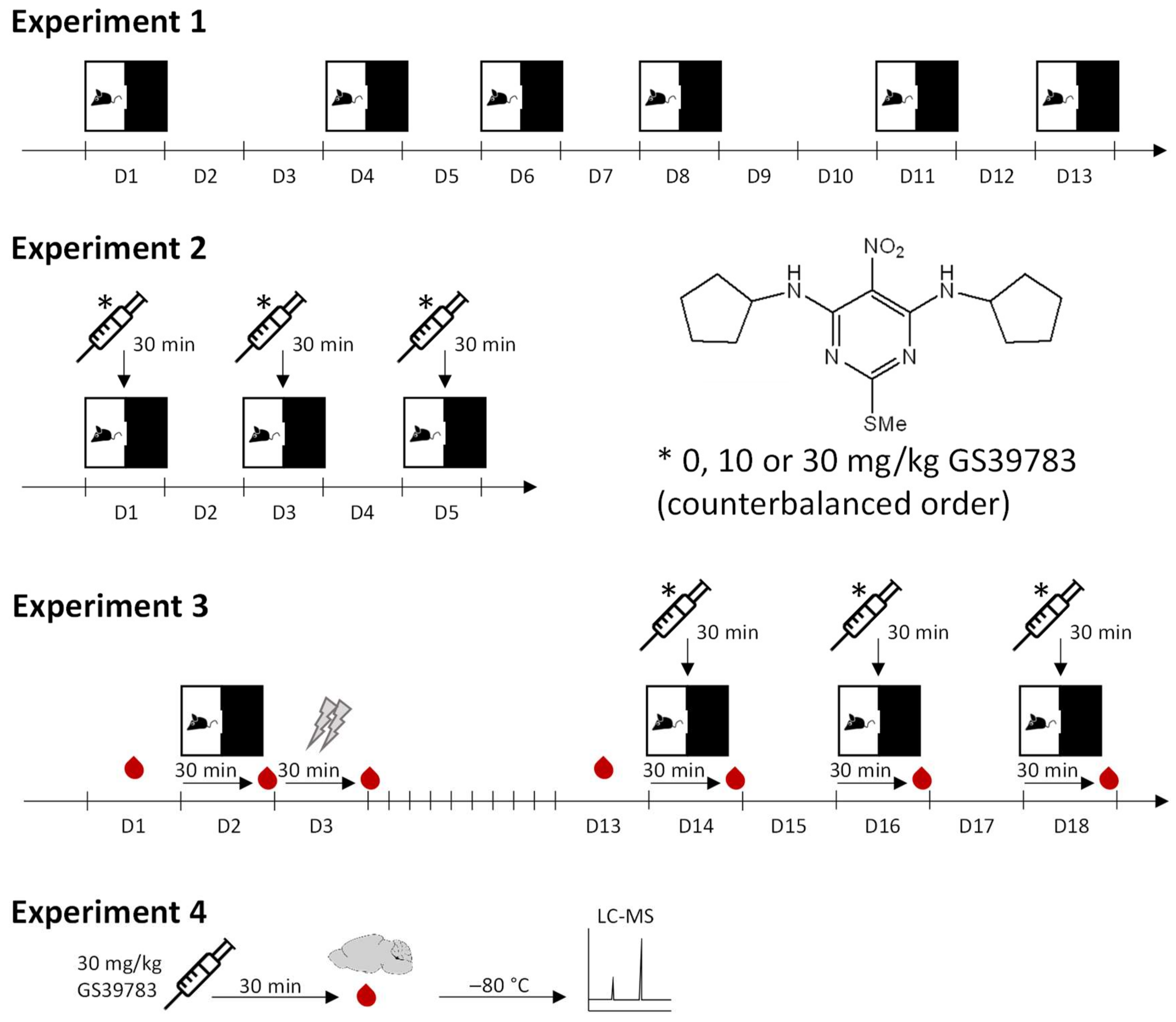
2.1. Experiment 1: Effects of Repeated Testing in the Light–Dark Box
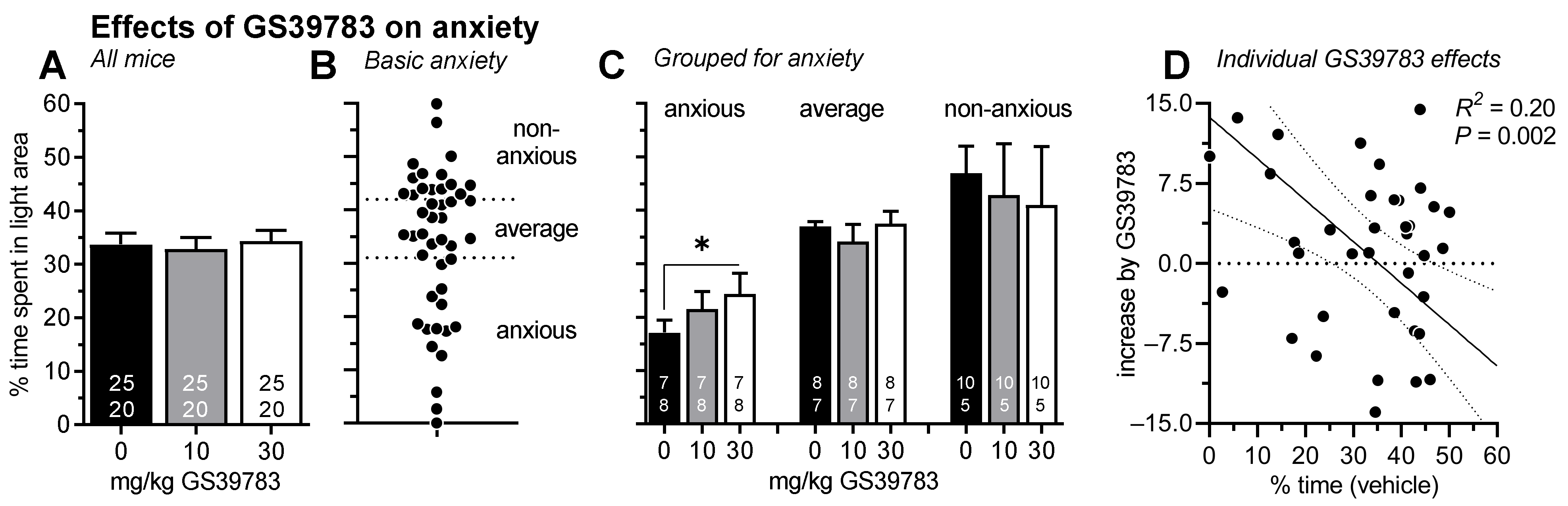
2.2. Experiment 2: Effects on GS39783 on Anxiety
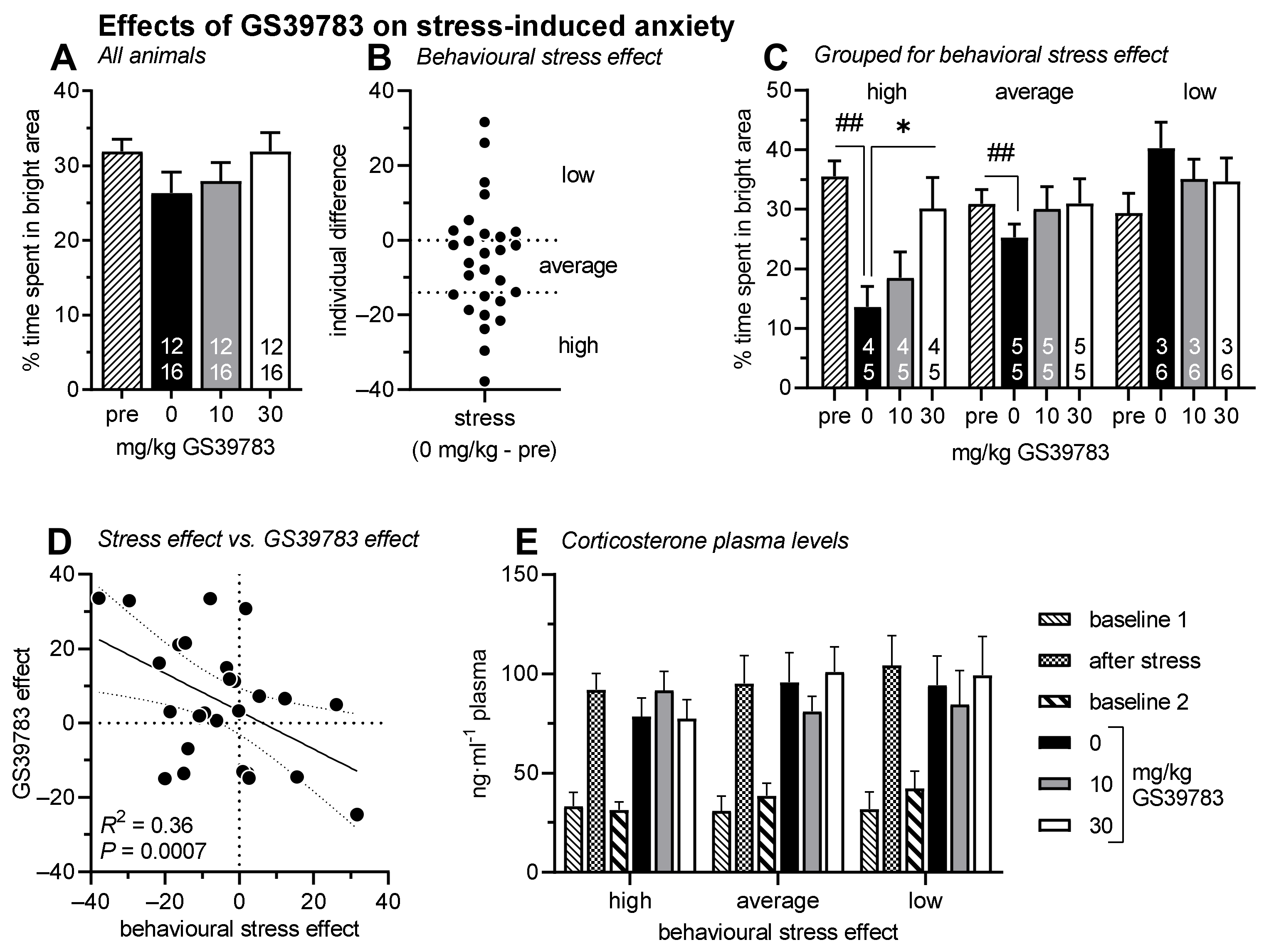
2.3. Stress-Induced Anxiety in the Light–Dark Box Test
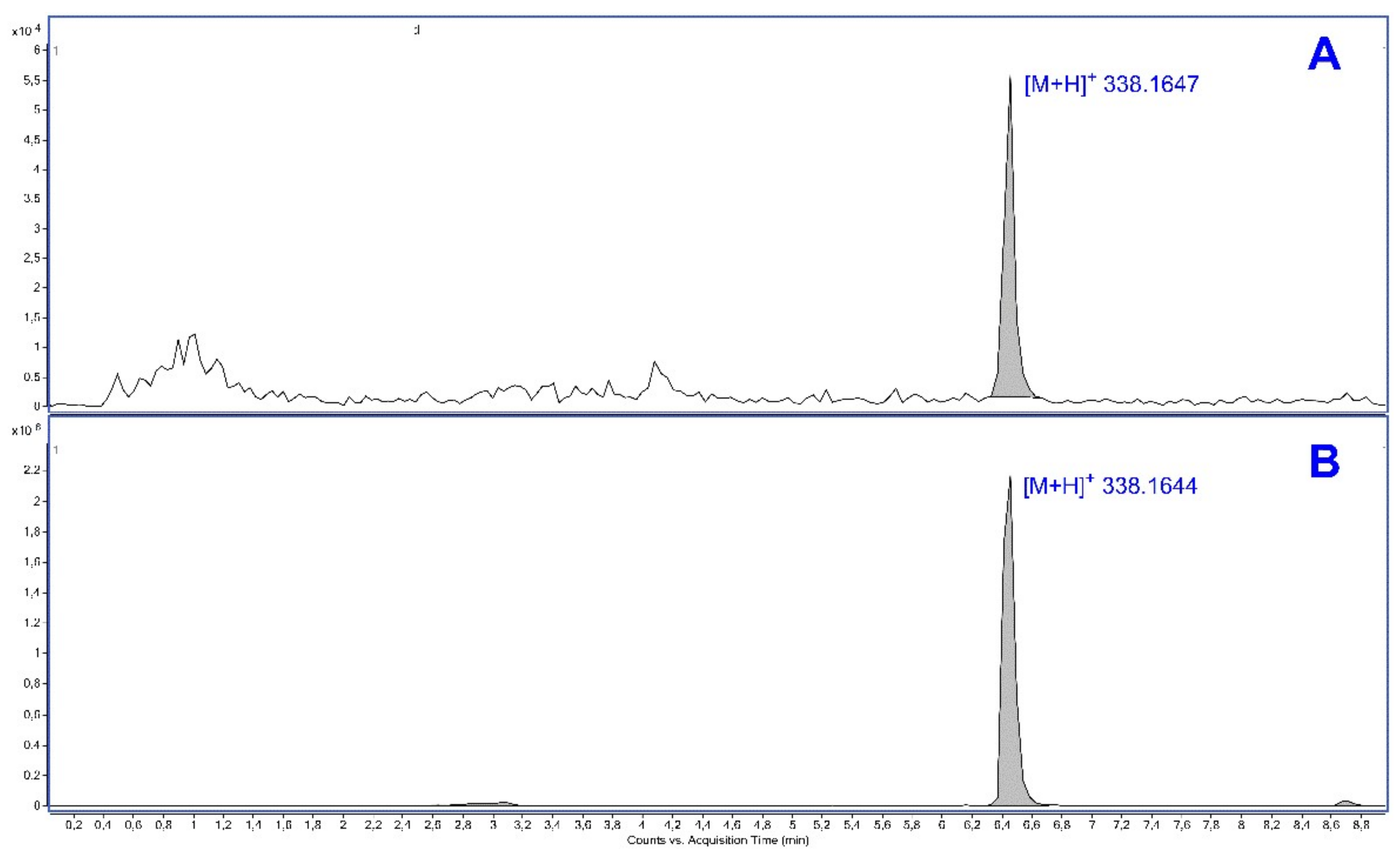
2.4. Plasma and Brain Levels of GS39783
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemical Reagents
4.3. Compound Formulation for Injection
4.4. Behavioral Experiments

4.5. Identification and Semi-Quantitative Analysis of GS39783 in Plasma and Brain (Experiment 4)
4.6. Descriptive and Analytical Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Millan, M.J. The neurobiology and control of anxious states. Prog. Neurobiol. 2003, 70, 83–244. [Google Scholar] [CrossRef]
- Brambilla, P.; Perez, J.; Barale, F.; Schettini, G.; Soares, J.C. GABAergic dysfunction in mood disorders. Mol. Psychiatry 2003, 8, 721–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenster, R.J.; Lebois, L.A.M.; Ressler, K.J.; Suh, J. Brain circuit dysfunction in post-traumatic stress disorder: From mouse to man. Nat. Rev. Neurosci. 2018, 19, 535–551. [Google Scholar] [CrossRef] [PubMed]
- Sartori, S.B.; Singewald, N. Novel pharmacological targets in drug development for the treatment of anxiety and anxiety-related disorders. Pharmacol. Ther. 2019, 204, 107402. [Google Scholar] [CrossRef] [PubMed]
- Castelli, M.P.; Gessa, G.L. Distribution and Localization of the GABAB Receptor. In GABAB Receptor; Colombo, G., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 75–92. [Google Scholar] [CrossRef]
- Mombereau, C.; Kaupmann, K.; van der Putten, H.; Cryan, J.F. Altered response to benzodiazepine anxiolytics in mice lacking GABA B(1) receptors. Eur. J. Pharmacol. 2004, 497, 119–120. [Google Scholar] [CrossRef] [PubMed]
- Spiering, M.J. The discovery of GABA in the brain. J. Biol. Chem. 2018, 293, 19159–19160. [Google Scholar] [CrossRef] [Green Version]
- Wick, J.Y. The history of benzodiazepines. Consult. Pharm. 2013, 28, 538–548. [Google Scholar] [CrossRef]
- Cryan, J.F.; Sweeney, F.F. The age of anxiety: Role of animal models of anxiolytic action in drug discovery. Br. J. Pharmacol. 2011, 164, 1129–1161. [Google Scholar] [CrossRef] [Green Version]
- Brohan, J.; Goudra, B.G. The Role of GABA receptor agonists in anesthesia and sedation. CNS Drugs 2017, 31, 845–856. [Google Scholar] [CrossRef]
- Atack, J.R. GABAA receptor subtype-selective modulators. I. α2/α3-selective agonists as non-sedating anxiolytics. Curr. Top. Med. Chem. 2011, 11, 1176–1202. [Google Scholar] [CrossRef]
- Nemeroff, C.B. The role of GABA in the pathophysiology and treatment of anxiety disorders. Psychopharmacol. Bull. 2003, 37, 133–146. [Google Scholar] [PubMed]
- Baldwin, D.S.; Anderson, I.M.; Nutt, D.J.; Allgulander, C.; Bandelow, B.; den Boer, J.A.; Christmas, D.M.; Davies, S.; Fineberg, N.; Lidbetter, N.; et al. Evidence-based pharmacological treatment of anxiety disorders, post-traumatic stress disorder and obsessive-compulsive disorder: A revision of the 2005 guidelines from the British Association for Psychopharmacology. J. Psychopharmacol. 2014, 28, 403–439. [Google Scholar] [CrossRef] [PubMed]
- Bandelow, B.; Zohar, J.; Hollander, E.; Kasper, S.; Möller, H.J.; Zohar, J.; Hollander, E.; Kasper, S.; Möller, H.J.; Bandelow, B.; et al. World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for the pharmacological treatment of anxiety, obsessive-compulsive and post-traumatic stress disorders—first revision. World J. Biol. Psychiatry 2008, 9, 248–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penn, E.; Tracy, D.K. The drugs don’t work? antidepressants and the current and future pharmacological management of depression. Adv. Psychopharmacol. 2012, 2, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Mombereau, C.; Kaupmann, K.; Gassmann, M.; Bettler, B.; van der Putten, H.; Cryan, J.F. Altered anxiety and depression-related behaviour in mice lacking GABAB(2) receptor subunits. Neuroreport 2005, 16, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Mombereau, C.; Kaupmann, K.; Froestl, W.; Sansig, G.; van der Putten, H.; Cryan, J.F. Genetic and pharmacological evidence of a role for GABA(B) receptors in the modulation of anxiety- and antidepressant-like behavior. Neuropsychopharmacology 2004, 29, 1050–1062. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Kelly, P.H.; Chaperon, F.; Gentsch, C.; Mombereau, C.; Lingenhoehl, K.; Froestl, W.; Bettler, B.; Kaupmann, K.; Spooren, W.P. Behavioral characterization of the novel GABAB receptor-positive modulator GS39783 (N,N’-dicyclopentyl-2-methylsulfanyl-5-nitro-pyrimidine-4,6-diamine): Anxiolytic-like activity without side effects associated with baclofen or benzodiazepines. J. Pharm. Exp. 2004, 310, 952–963. [Google Scholar] [CrossRef] [Green Version]
- Kalinichev, M.; Girard, F.; Haddouk, H.; Rouillier, M.; Riguet, E.; Royer-Urios, I.; Mutel, V.; Lütjens, R.; Poli, S. The drug candidate, ADX71441, is a novel, potent and selective positive allosteric modulator of the GABA(B) receptor with a potential for treatment of anxiety, pain and spasticity. Neuropharmacology 2017, 114, 34–47. [Google Scholar] [CrossRef]
- Jacobson, L.H.; Cryan, J.F. Evaluation of the anxiolytic-like profile of the GABAB receptor positive modulator CGP7930 in rodents. Neuropharmacology 2008, 54, 854–862. [Google Scholar] [CrossRef]
- Kaupmann, K.; Cryan, J.F.; Wellendorph, P.; Mombereau, C.; Sansig, G.; Klebs, K.; Schmutz, M.; Froestl, W.; van der Putten, H.; Mosbacher, J.; et al. Specific gamma-hydroxybutyrate-binding sites but loss of pharmacological effects of gamma-hydroxybutyrate in GABA(B)(1)-deficient mice. Eur. J. Neurosci. 2003, 18, 2722–2730. [Google Scholar] [CrossRef]
- Olivier, B.; Zethof, T.; Pattij, T.; van Boogaert, M.; van Oorschot, R.; Leahy, C.; Oosting, R.; Bouwknecht, A.; Veening, J.; van der Gugten, G.; et al. Stress-induced hyperthermia and anxiety: Pharmacological validation. Eur. J. Pharm. 2003, 463, 117–132. [Google Scholar] [CrossRef]
- Jacobson, L.H.; Vlachou, S.; Slattery, D.A.; Li, X.; Cryan, J.F. The gamma-aminobutyric acid B receptor in depression and reward. Biol. Psychiatry 2018, 83, 963–976. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Risbrough, V.B.; Cates-Gatto, C.; Kaczanowska, K.; Finn, M.G.; Roberts, A.J.; Markou, A. Comparison of the effects of the GABAB receptor positive modulator BHF177 and the GABAB receptor agonist baclofen on anxiety-like behavior, learning, and memory in mice. Neuropharmacology 2013, 70, 156–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mareš, P.; Tichá, K.; Mikulecká, A. Anticonvulsant and behavioral effects of GABA(B) receptor positive modulator CGP7930 in immature rats. Epilepsy Behav. 2013, 28, 113–120. [Google Scholar] [CrossRef]
- Sweeney, F.F.; O’Leary, O.F.; Cryan, J.F. GABAB receptor ligands do not modify conditioned fear responses in BALB/c mice. Behav. Brain Res. 2013, 256, 151–156. [Google Scholar] [CrossRef]
- Li, X.; Kaczanowska, K.; Finn, M.G.; Markou, A.; Risbrough, V.B. The GABA(B) receptor positive modulator BHF177 attenuated anxiety, but not conditioned fear, in rats. Neuropharmacology 2015, 97, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Bourin, M.; Hascoet, M. The mouse light/dark box test. Eur. J. Pharm. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Urwyler, S.; Pozza, M.F.; Lingenhoehl, K.; Mosbacher, J.; Lampert, C.; Froestl, W.; Koller, M.; Kaupmann, K. N,N’-Dicyclopentyl-2-methylsulfanyl-5-nitro-pyrimidine-4,6-diamine (GS39783) and structurally related compounds: Novel allosteric enhancers of gamma-aminobutyric acidB receptor function. J Pharm. Exp. 2003, 307, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Gjoni, T.; Desrayaud, S.; Imobersteg, S.; Urwyler, S. The positive allosteric modulator GS39783 enhances GABA(B) receptor-mediated inhibition of cyclic AMP formation in rat striatum in vivo. J. Neurochem. 2006, 69, 1416–1422. [Google Scholar] [CrossRef]
- Blumstein, L.K.; Crawley, J.N. Further characterization of a simple, automated exploratory model for the anxiolytic effects of benzodiazepines. Pharm. Biochem Behav. 1983, 18, 37–40. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Martin, B.R. Neuropharmacological and physiological validation of a computer-controlled two-compartment black and white box for the assessment of anxiety. Prog. Neuropsychopharmacol. Biol. Psychiatry 1989, 13, 963–976. [Google Scholar] [CrossRef]
- Cryan, J.F.; Kaupmann, K. Don’t worry ‘B’ happy!: A role for GABA(B) receptors in anxiety and depression. Trends Pharm. Sci. 2005, 26, 36–43. [Google Scholar] [CrossRef]
- Jacobson, L.H.; Cryan, J.F. Genetic approaches to modeling anxiety in animals. Curr. Top. Behav. Neurosci. 2010, 2, 161–201. [Google Scholar] [CrossRef] [PubMed]
- Tucker, L.B.; McCabe, J.T. Behavior of male and female C57BL/6J mice is more consistent with repeated trials in the elevated zero maze than in the elevated plus maze. Front. Behav. Neurosci. 2017, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrader, A.J.; Taylor, R.M.; Lowery-Gionta, E.G.; Moore, N.L.T. Repeated elevated plus maze trials as a measure for tracking within-subjects behavioral performance in rats (Rattus norvegicus). PLoS ONE 2018, 13, e0207804. [Google Scholar] [CrossRef] [Green Version]
- Ilse, A.; Prameswari, V.; Kahl, E.; Fendt, M. The role of trait anxiety in associative learning during and after an aversive event. Learn. Mem. 2019, 26, 56–59. [Google Scholar] [CrossRef]
- Crawley, J.N.; Belknap, J.K.; Collins, A.; Crabbe, J.C.; Frankel, W.; Henderson, N.; Hitzemann, R.J.; Maxson, S.C.; Miner, L.L.; Silva, A.J.; et al. Behavioral phenotypes of inbred mouse strains: Implications and recommendations for molecular studies. Psychopharmacology 1997, 132, 107–124. [Google Scholar] [CrossRef]
- Liebsch, G.; Montkowski, A.; Holsboer, F.; Landgraf, R. Behavioral profiles of two Wistar rat lines selectivly bred for high or low anxiety-related behaviour. Behav. Brain Res. 1998, 94, 301–310. [Google Scholar] [CrossRef]
- Garcia, A.M.; Cardenas, F.P.; Morato, S. Effect of different illumination levels on rat behavior in the elevated plus-maze. Physiol. Behav. 2005, 85, 265–270. [Google Scholar] [CrossRef]
- Bluett, R.J.; Gamble-George, J.C.; Hermanson, D.J.; Hartley, N.D.; Marnett, L.J.; Patel, S. Central anandamide deficiency predicts stress-induced anxiety: Behavioral reversal through endocannabinoid augmentation. Transl. Psychiatry 2014, 4, e408. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, L.H.; Bettler, B.; Kaupmann, K.; Cryan, J.F. Behavioral evaluation of mice deficient in GABA(B(1)) receptor isoforms in tests of unconditioned anxiety. Psychopharmacology 2007, 190, 541–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse | Plasma (µg/mL) | Brain (µg/g) | Brain/Plasma Ratio |
|---|---|---|---|
| 1 | 219.8 | 71.0 | 0.32 |
| 2 | 222.3 | 74.2 | 0.33 |
| 3 | 209.8 | 26.1 | 0.12 |
| Mean ± SEM | 217.3 ± 3.8 | 57.1 ± 15.5 | 0.26 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bicakci, A.O.; Sarkar, M.; Chang, Y.-H.; Kahl, E.; Ragazzi, L.; Moldes-Anaya, A.; Fendt, M. Anxiolytic-like Effects of the Positive GABAB Receptor Modulator GS39783 Correlate with Mice’s Individual Basal Anxiety and Stress Reactivity. Pharmaceuticals 2022, 15, 233. https://doi.org/10.3390/ph15020233
Bicakci AO, Sarkar M, Chang Y-H, Kahl E, Ragazzi L, Moldes-Anaya A, Fendt M. Anxiolytic-like Effects of the Positive GABAB Receptor Modulator GS39783 Correlate with Mice’s Individual Basal Anxiety and Stress Reactivity. Pharmaceuticals. 2022; 15(2):233. https://doi.org/10.3390/ph15020233
Chicago/Turabian StyleBicakci, Ahmet Oguzhan, Mousumi Sarkar, Yu-Hsin Chang, Evelyn Kahl, Lorenzo Ragazzi, Angel Moldes-Anaya, and Markus Fendt. 2022. "Anxiolytic-like Effects of the Positive GABAB Receptor Modulator GS39783 Correlate with Mice’s Individual Basal Anxiety and Stress Reactivity" Pharmaceuticals 15, no. 2: 233. https://doi.org/10.3390/ph15020233