
Non-Apoptotic Programmed Cell Death in Thyroid Diseases

Thyroid Surgery, The First Affiliated Hospital of Zhengzhou University, Zhengzhou 450052, China
*
Author to whom correspondence should be addressed.
Pharmaceuticals 2022, 15(12), 1565; https://doi.org/10.3390/ph15121565
Submission received: 30 October 2022
/
Revised: 11 December 2022
/
Accepted: 13 December 2022
/
Published: 15 December 2022
(This article belongs to the Special Issue Targeting Thyroid Cancer: From Biology to Therapeutic Strategies)
Abstract
:Thyroid disorders are among the most common endocrinological conditions. As the prevalence of thyroid diseases increases annually, the exploration of thyroid disease mechanisms and the development of treatments are also gradually improving. With the gradual advancement of therapies, non-apoptotic programmed cell death (NAPCD) has immense potential in inflammatory and neoplastic diseases. Autophagy, pyroptosis, ferroptosis, and immunogenic cell death are all classical NAPCD. In this paper, we have compiled the recent mechanistic investigations of thyroid diseases and established the considerable progress by NAPCD in thyroid diseases. Furthermore, we have elucidated the role of various types of NAPCD in different thyroid disorders. This will help us to better understand the pathophysiology of thyroid-related disorders and identify new targets and mechanisms of drug resistance, which may facilitate the development of novel diagnostic and therapeutic strategies for patients with thyroid diseases. Here, we have reviewed the advances in the role of NAPCD in the occurrence, progression, and prognosis of thyroid diseases, and highlighted future research prospects in this area.
1. Introduction
1.1. Thyroid Disease
The thyroid gland is an endocrine organ, similar in shape to a butterfly, that secretes thyroid hormones [1]. Thyroid disorders are a general term for disorders in which the function, size, and structure of the thyroid gland are altered. Thyroid disorders are common in endocrinology and include goiter, thyroiditis, thyroid nodules, thyroid cancer (TC), hypothyroidism, and hyperthyroidism.
As the most common endocrine malignancy [2], TC incidence has increased rapidly in the last few decades [3,4]. TC can be classified as papillary thyroid cancer (PTC), follicular thyroid cancer (FTC), medullary thyroid cancer (MTC), and anaplastic thyroid cancer (ATC). TC develops from two cell types: PTC, FTC and ATC originate from follicular cells that line the colloidal follicles and are responsible for thyroid hormone biosynthesis and iodine uptake [5,6]. Parafollicular cells (also called C cells), the origin of MTC, are another cell type of the thyroid gland responsible for synthesizing and secreting calcitonin hormones [7,8,9]. Among these, PTC and FTC, which account for 90% of all cases, have a survival rate of 90% and an overall good prognosis [9]. However, a small proportion of patients, such as those with invasive, or metastatic types of cancer, have a poor prognosis [10,11]. In addition, as one of the most aggressive human malignancies, the post-diagnosis median survival of patients with ATC is approximately 3–5 months [12]. In addition to conventional surgery, several therapies exist, such as radioactive iodine (RAI, 131I) [13] and tyrosine kinase inhibitors (TKIs) [14]. However, these treatments have limitations, including drug and radioactivity resistance and adverse side effects [10]. Therefore, exploring new treatments for TC remains a current research priority in the discipline.
As a typical organ-specific autoimmune disease, autoimmune thyroiditis (AIT) is the leading cause of hypothyroidism, with a population prevalence of approximately 1–5% [15,16]. Graves’ disease (GD) and Hashimoto’s thyroiditis (HT) are the two most common autoimmune diseases [15,17]. AIT pathogenesis involves complex interactions between genes and the environment. Although HT and GD have opposite clinical manifestations, they share a common etiology of a decreased tolerance to thyroid autoantigens [18]. As a non-neoplastic disease of the thyroid, AIT has also been the focus of academic research. The mechanistic exploration of AIT has led to a greater understanding of thyroid autoimmunity, which in turn has allowed us to identify new therapeutic targets and facilitate the exploration of new treatment options [19,20].
However, the factors affecting thyroid disease remain unclear. Research on the factors influencing its occurrence is of great significance for preventing and controlling thyroid disease.
1.2. Non-Apoptotic Programmed Cell Death (NAPCD)
The Nomenclature Committee on Cell Death (NCCD) classifies cell death into programmed cell death (PCD) and accidental cell death (ACD), defining and explaining cell death from morphological, biochemical, and functional perspectives [21,22]. ACD refers to catastrophic cell death under natural conditions [23]. As an autonomous cell death process, PCD involves intracellular suicide pathways controlled by strict genetic mechanisms that maintain a stable internal environment, critical in response to inflammation, infection, and injury [24]. Based on its different mechanisms [25], PCD has two major categories: apoptosis and non-apoptotic programmed cell death (NAPCD). NAPCD includes autophagy, ferroptosis, pyroptosis, immunogenic cell death, mitochondrial catastrophe, necroptosis, and anoikis [26]. As research on NAPCD continues to advance, we have found that NAPCD has excellent potential in the pathogenesis and diagnosis of various diseases, such as various malignant tumors and even in benign diseases such as hearing impairment [22,27,28,29,30].
As the most common form of NAPCD [31], autophagy refers to cellular self-digestion mediated by lysosomal hydrolases to maintain normal intracellular and tissue homeostasis when cells lack nutrients or are affected by inflammation [32]. Ferroptosis is also a form of NAPCD, characterized by lipid peroxidation damage to the cell membrane and the production of iron ion-dependent lipid reactive oxygen species [33]. Pyroptosis is a recently proposed NAPCD-dependent activation of cellular inflammation through inflammatory forms of regulated cell death [34]. It can lead to PCD under various conditions [35]. Immunogenic cell death (ICD) is a new type of NAPCD [36] that activates adaptive immune responses in an immunoreactive environment [22]. ICD is caused by certain chemotherapeutic agents, lytic virus, physical chemotherapy, photodynamic therapy, and radiotherapy, which induce cell death by activating the immune system of an immunocompetent host [37]. Other types of NAPCD, such as mitotic catastrophe, necroptosis, and anoikis, have been less reported in thyroid-related diseases [38,39].
In conclusion, NAPCD provides a new frontier in the pathogenesis and treatment of the disease. In this paper, we have reviewed the progress of research on NAPCD in benign and malignant thyroid diseases, including efficacy prediction, drug resistance, therapeutic targets, and the relationship between different NAPCDs for clinical diagnosis and treatment.
2. Autophagy and Thyroid Disease
Autophagy is the most common type of NAPCD, and as a highly conserved physiological process, it removes damaged organelles and abnormal proteins through lysosomal degradation [40]. Autophagy is implicated in various pathological and physiological processes, including neurodegenerative diseases [41], the maintenance of intracellular homeostasis [42], inflammation [43], and cancer [44]. Autophagy is widely believed to have a dual effect on cells. On the one hand, it supports cell survival by adjusting the physiologically relevant mechanisms required to support cell proliferation and survival and maintains a stable internal environment [45,46]. On the other hand, it regulates cell death by regulating cellular autophagy-related mechanisms [47,48,49].
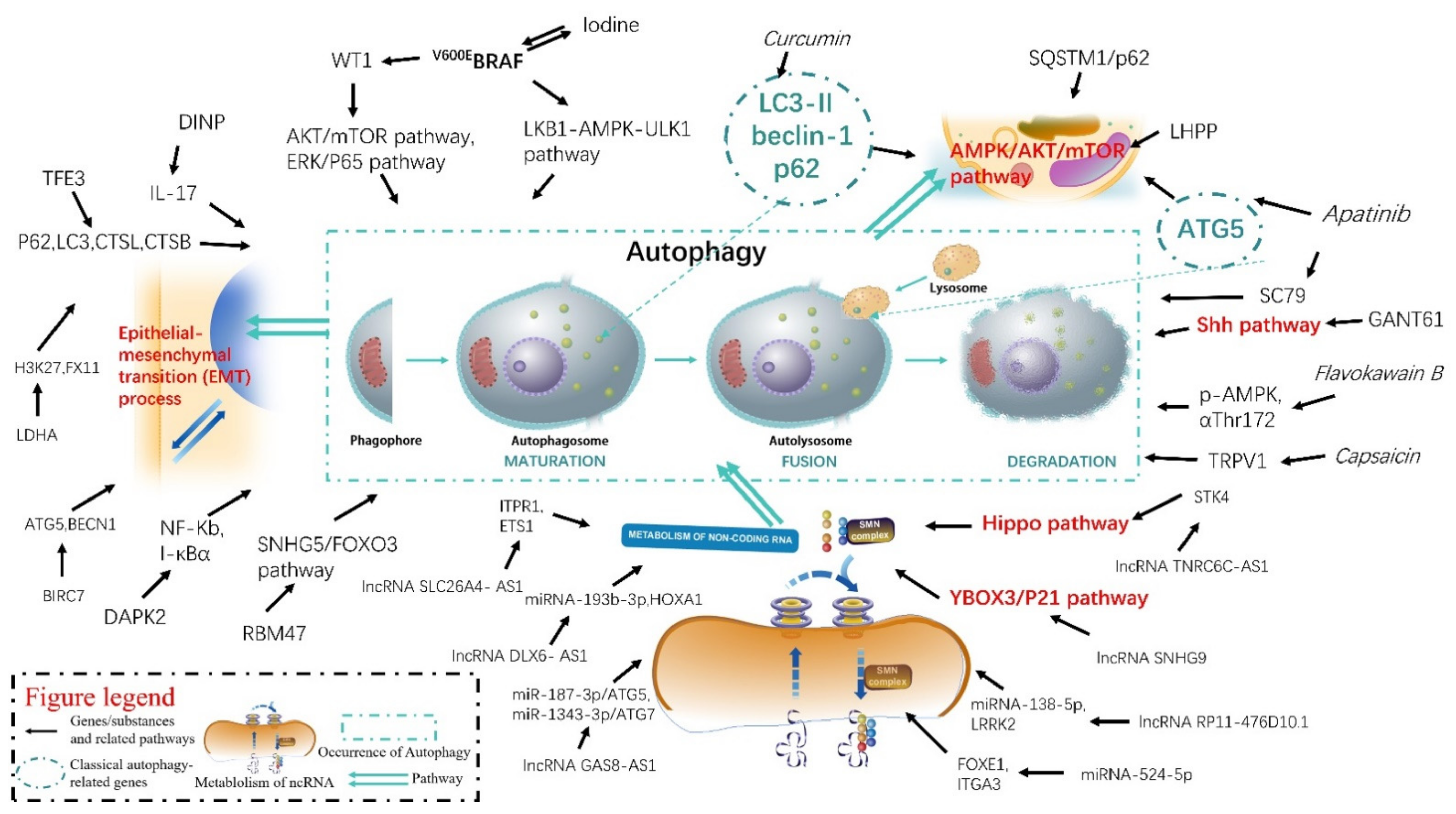
A comprehensive understanding of the mechanisms of autophagy in the pathogenesis and progression of thyroid diseases will help determine the appropriate timing, effective therapeutic targets, and provide innovative ideas for diagnosing and treating thyroid-related diseases. We attempted to elucidate the changes in autophagy-related gene or RNA expression, therapeutic targets, and natural substances or their extracts in TC, which help us to better understand the autophagy mechanism in TC occurrence and its development comprehensively (Figure 1).
2.1. The Role of Genes and Autophagy in Thyroid Disease
Certain genes can significantly affect autophagy by triggering changes at the protein or RNA level, which may promote or inhibit the development of thyroid-related diseases. By elucidating the genes that affect the occurrence and development of thyroid-related diseases, we have a deeper understanding of the pathogenesis of thyroid diseases, and also laid the foundation for us to subsequently explore target molecules that can help in the early diagnosis of diseases and the development of targeted drugs.
BRAF mutations are the most common genetic lesions in thyroid tumors, with an incidence of 45% in PTC and 25% in ATC [50]. Currently, V600EBRAF mutations are considered highly-specific diagnostic genetic markers for PTC, and V600EBRAF is closely related to the development and metastasis of PTC [51]. V600EBRAF mutants activate marker pathways and promote cancer progression in PTC [52] and Wilms’s tumor 1 (WT1), encoded as a transcription factor located on chromosome 11p13 [53]. Targeting the V600EBRAF mutant is an effective treatment for PTC, and BRAF activation of WT1 promotes the growth of PTC and regulates autophagy and apoptosis [54].
Adenosine monophosphate-activated protein kinase (AMPK) regulates cellular metabolism as an energy sensor by mediating the insulin pathway [55,56]. Autophagy can be modulated through the AKT/AMPK/mTOR pathway [57,58]. Two studies have identified the role of autophagy and the AKT/AMPK/mTOR pathway in thyroid tumors. Sequestosome 1 (SQSTM1), also known as p62, is a vital gene in autophagy that regulates intracellular protein degradation [59]. SQSTM1 has been suggested to regulate autophagy via the AKT/AMPK/mTOR signaling pathway to trigger autophagy and promote the growth of papillary thyroid cancer cells [60]. Some researchers have suggested that SIRT6 can inhibit the glucose transporter protein 1 (GLUT1) through autophagy-mediated degradation, thereby suppressing the Warburg effect that affects tumor growth and development [60]. Furthermore, as a histidine phosphatase, phospholysine phosphohistidine inorganic pyrophosphate phosphatase (LHPP) is an antitumor factor [61,62]. LHPP similarly inhibits papillary TC cell growth and migration by regulating the AKT/AMPK/mTOR signaling pathway and triggering autophagy [57]. In addition, the sonic hedgehog (Shh) pathway has been implicated in autophagy in TC. The Shh pathway is highly activated in various malignancies and plays an essential role in tumor development [63]. Inhibition of the hedgehog pathway has been suggested to activate TGF-β-activated kinase (TAK1), which inhibits the apoptosis of thyroid tumor cells by inducing autophagy onset [64].
Epithelial-mesenchymal transition (EMT) appears early in the tumor metastasis process and plays a crucial role in mediating the development of aggressive tumor phenotypes. EMT is a multistage process in which cells lose their epithelial properties and undergo significant changes in morphology, adhesion, and migration capacity [65]. The core features of the EMT include decreased adhesion and increased motility [66]. Baculoviral IAP repeat-containing 7 (BIRC7) has been suggested to promote epithelial-mesenchymal transition and metastasis in papillary TC by inhibiting autophagy [67]. Lactate dehydrogenase A (LDHA) is an important enzyme involved in the Warburg effect. This leads to the formation of an acidic microenvironment in the tumor that promotes EMT and metastasis [68]. LDHA has been suggested to regulate autophagy to promote metastasis and tumorigenesis in PTC by inducing EMT gene transcription [68].
In addition, several other genes have been reported to influence the occurrence and development of thyroid disease through the autophagic pathway. FOXO3 belongs to the Forkhead Box (FOX) family of transcription factors [69]. The aberrant activation of FOXO3 has been extensively studied in cancer development and progression [70]. FOXO3 can promote autophagy through the transcriptional activation of autophagy-related genes, suggesting that FOXO3 can serve as a marker of autophagy [71]. The RNA binding motif protein 47 (RBM47)/small nucleolar RNA host gene 5 (SNHG5)/FOXO3 axis inhibits PTC cell proliferation by activating autophagy [72]. Furthermore, as another critical transcription factor of the FOX family [73], FOXK2 was found to promote the proliferation of PTC cells through the downregulation of autophagy [74]. BIRC7 is a newly identified member of the IAP family that is largely absent in normal tissues but is expressed at elevated levels in a range of tumor types [75]. The overexpression and expression of BIRC7 in tumors are related to the increased resistance to chemotherapy and decreased patient survival [76]. BIRC7 is considered a potential new target for thyroid tumor therapy. As a regulator of autophagy and lysosomal biogenesis, the transcription factor E3 (TFE3) belongs to the microphthalmia/transcription factor E (MiT/TFE) family, located on the short arm of the X chromosome 11.22 [77]. TFE3 can contribute to the invasion and metastasis of PTC by regulating autophagy [78]. As a member of the Ca2+/calmodulin-regulated serine/threonine kinases family, death-associated protein kinase 2 (DAPK2) is a tumor suppressor that affects various cellular activities, including cellular immune function and cell death. Recent studies have shown that DAPK2 can participate in autophagy and activate NF-κB through the autophagy-dependent degradation of IκBα, affecting TC development and progression [79].
With continuous advances in high-throughput genome sequencing technology, we have observed that 90% of the human genome could undergo transcription [80]. However, not all RNAs can be translated into proteins [81]. We refer to genes that do not directly encode proteins as non-coding RNAs (ncRNAs). ncRNAs play vital roles in human disease progression by regulating gene expression [82]. ncRNAs can contribute to mRNA degradation and protein translation failure by mediating post-transcriptional gene silencing [83]. In addition, ncRNAs also remodel chromatin structure by altering heterochromatin formation [84], thus enhancing or repressing gene expression [83]. These regulations can affect cellular function and help maintain homeostasis in vivo [85]. Several ncRNAs influence the onset of thyroid-related diseases through autophagy in thyroid-related diseases.
Long non-coding RNAs (lncRNAs) are classical ncRNAs: a heterogeneous family of RNA molecules greater than 200 nucleotides in length. They have gained widespread attention for their potential roles in organism development and disease [86]. Aberrant lncRNA expression has been observed in various cancers [87]. LncRNAs in PTC carcinogenesis and development play a vital role in thyroid disease development through autophagic mechanisms. LncRNA SLC26A4-AS1 can inhibit PTC progression by recruiting ETS1 to promote inositol 1,4,5-trisphosphate receptor type 1 (ITPR1)-mediated autophagy [88]. The lncRNA distal-less homeobox 6 antisense RNA 1 (DLX6-AS1) can interact with microRNA-193b-3p to inhibit TC progression by suppressing homeobox A1 (HOXA1) and enhancing autophagy and apoptosis in TC cells [89]. The lncRNA TNRC6C-AS1 promotes serine/threonine kinase 4 (STK4) methylation and inhibits TC cell autophagy through the hippo signaling pathway [90]. Activating transcription factor-2 (ATF2)-inducible lncRNA growth arrest-specific 8 (GAS8)-AS1 promotes TC cell progression by targeting miR-1343-3p/ATG7 and miR-187-3p/ATG5 axes to promote autophagy in TC cells [91]. LncRNA RP11-476D10.1 can enhance autophagy in PTC cells while inhibiting their proliferation through microRNA-138-5p-dependent inhibition of LRRK2 [92]. Furthermore, SNHG9, an exosome-rich lncRNA in PTC cells, can inhibit autophagy through the YBOX3/P21 pathway in normal thyroid epithelial cells [93].
MicroRNAs (miRNAs) are a class of non-coding RNAs of 18–24 nucleotides in length involved in various physiological processes. miRNAs are involved in target gene regulation by inhibiting protein production through binding to complementary mRNAs and are tissue- and stage-specific [94]. miRNA-524-5p inhibits the progression of PTC cells by targeting FOXE1 and ITGA3 in the cellular autophagy and recycling pathways [95].
2.2. The Role of Inhibitors, Substances, and Autophagy in Thyroid-Related Diseases
Natural products have been used as alternative therapies for various diseases, including inflammation and cancer. They have received attention from the academic community because of their cost and relatively few side effects [96,97].
Curcumin is widely used in traditional medicine and is a phytochemical isolated from the spice turmeric (Curcuma longa) [98]. Curcumin, used primarily as an adjuvant in cancer treatment, is undergoing extensive clinical trials resulting in favorable results [99,100,101]. Curcumin reportedly induces autophagic death in human thyroid cancer cells [102].
ATC is the most lethal subtype of TC. Lacking of sodium iodide synthetics (NIS) is a characteristic of the highly dedifferentiated state of ATC115. Therefore, radioactive iodine (RAI) therapy, which relies on iodine uptake by NIS channels, cannot be applied to ATC patients [103,104]. However, severe side effects can develop in TC patients treated with chemotherapy, such as high blood pressure, hypocalcemia, and hypoalbuminemia and drug resistance [105]. This means that finding new targeted drugs or exploring multidrug combinations may be a new research direction for the treatment of TC. As novel orally targeted TKIs, apatinib can the inhibit vascular endothelial growth factor receptor 2 (VEGFR2) with high selectivity. Importantly, apatinib has shown promising efficacy in few patients with radioiodine-refractory differentiated TC [106]. Apatinib has been shown to inhibit proliferation and induce autophagy through the PI3K/Akt/mTOR signaling pathway in human PTC cells [107]. Apatinib has been suggested to induce autophagy by downregulating p-AKT and p-mTOR signaling through the AKT/mTOR pathway in human ATC cells [108]. It has been reported that apatinib induces autophagy and apoptosis in human ATC cells by blocking the Akt/GSK3/ANG pathway to inhibit angiogenesis in mesenchymal TC [109]. In addition, as a natural alkaloid derived from Capsicum spp. [110], Capsaicin (trans-8-methyl-N-vanillyl-6-nonenamide, CAP) is a natural alkaloid [110], which inhibits the stemness of mesenchymal TC cells by activating autophagic lysosomal-mediated degradation of OCT4A [111]. The action of CAP on ATC helps us explore new ATC-targeting drugs [108,112,113,114].
Herbal plants have a long history of clinical application in China. Prunella vulgaris L. (PV) is a traditional herbal medicine used in ancient China to treat thyroid disorders [115]. Under environmental stress conditions, AMPK, the mammalian target of rapamycin (mTOR), and unc-51-like autophagy-activated kinase 1 (ULK1) constitute a pathway that can initiate cellular autophagy [116]. PV aqueous extract can inhibit the growth of papillary thyroid carcinomas through the induction of autophagy in vitro and in vivo, possibly due to being autophagy-mediated by the AMPK/ mTOR/ULK1 pathway [117].
In the past, the vast majority of the world’s population consumed iodized salt to reduce iodine deficiency disorders, leaving most of the population in a state of iodine excess [118,119]. High iodine levels may affect the occurrence of PTC through BRAF gene mutations [120]. V600EBRAF can render PTC more susceptible to extrathyroidal infiltration and lymph node metastasis by aberrantly activating the BRAF/MEK/ERK (MAPK) signaling pathway [121]. High iodine levels are an important risk factor in the formation of V600EBRAF mutation-associated tumors, followed by increased overexpression and activity of BRAF kinase [122]. Studies by several authors indicate that autophagy induced by BRAF kinase in PTC cells is involved in anti-apoptosis, and promotes proliferation and migration at high iodine concentrations [123].
In addition, some specific inhibitors, or biochemicals, may also affect thyroid-related diseases through the autophagic pathway. Di-isonylphthalate (DINP) has a wide range of applications in artificial leather and coated fabrics [124]. It has been suggested that DINP exacerbates autoimmune thyroid disease in Wistar rats by inhibiting autophagy via the activation of the Akt/mTOR pathway [125]. Furthermore, some researchers have suggested that the adenosine 5’-monophosphate-activated protein kinase-dependent mTOR pathway is involved in Flavokawain BLHPP (FKB)-induced autophagy in TC cells [126].
2.3. Role of Drug Resistance and Autophagy in Thyroid Disease
Drug resistance is a classic theme in cancer therapy, and its development may be based on multiple mechanisms [127,128,129]. Autophagy, however, is a new cause of drug resistance in tumors that has been proposed in recent years [130,131,132,133,134]. Many therapeutic regimens induce cytoprotective autophagy, rendering cancer cells less sensitive to these drugs. Our exploration of the mechanisms of autophagy helps us uncover new targets, which may lead to breakthroughs in drug resistance to anticancer therapies [135].
The presence of V600EBRAF mutations is strongly associated with rapid TC progression, extrathyroidal infiltration, lymph node metastasis, and tumor recurrence [50,136]. Several V600EBRAF inhibitors (BRAFi), such as vemurafenib and dabrafenib, have been marketed for approval. Targeting V600EBRAF therapies have resulted in benefits to many patients [137]. However, a significant proportion of patients still develop resistance to BRAFi and progress to more advanced diseases. Melanoma is considered one of the most aggressive forms of skin cancer and the use of BRAF inhibitors, such as vemurafenib and dabrafenib, is revolutionizing the treatment of melanoma. Unfortunately, the duration of response to these drugs is limited due to the development of acquired resistance [138]. V600EBRAF in TC cells inhibition has been shown to induce cytoprotective autophagy via the AMPK-ULK1 pathway [139]. This provides a deeper insight into the mechanisms of resistance to BRAFi.
3. Ferroptosis and Thyroid Disease
Ferroptosis is a type of NAPCD that relies on the continuous accumulation of lipid peroxides in the cell membrane, which ultimately leads to cell death [140]. The ferroptosis pathway is responsible for reducing lipid peroxides, mediated by the inactivation of glutathione peroxidase 4 (GPX4) [140]. In recent years, a growing number of studies have shown that ferroptosis is strongly related to cancer development and progression and opens up new possibilities for cancer therapy [141]. Targeting ferroptosis is an emerging anticancer strategy [142]. In previous studies from our team, we found that ferroptosis plays an important role in thyroid tumors [143].
A recent study examined several genes associated with ferroptosis that may influence immune infiltration and progression of TC, including the arachidonic acid 5-lipoxygenase-activating protein (ALOX5AP), B-cell CLL/lymphoma 3 (BCL3), and apolipoprotein E (APOE) [144]. Ferroptosis-associated ALOX5AP, BCL3, and APOE gene polymorphisms have been associated with TC risk [145]. These results help to better understand the relationship between TC susceptibility and genetic polymorphisms of ferroptosis-related genes.
As an essential dietary vitamin derived from fruits and vegetables, vitamin C can protect healthy cells from oxidative damage and act as a scavenger of free radicals in the body [146]. Vitamin C has been shown to induce ferroptosis in mesenchymal TC cells through ferritin phage activation [147]. This suggests that finding genes related to ferroptosis or autophagy may provide new targets for ATC therapy.
4. Pyroptosis and Thyroid Disease
Pyroptosis is a NAPCD that has only recently begun to attract the attention of scientists. Pyroptosis is believed to exist principally as a defense against pathogens by triggering an antimicrobial response through the release of immunogenic cellular content, including damage-associated molecular patterns (DAMPs) and inflammatory cytokines, which can lead to programmed cell death in various contexts [148,149]. Unlike other NAPCDs, cellular pyroptosis has a complex effect on the microenvironment. On the one hand, pyroptosis can affect the tumor immune microenvironment by affecting immune cells; on the other hand, many inflammatory factors are released during pyroptosis as normal cells are stimulated [150,151]. Pyroptosis was initially thought to be a primitive immune response to pathogens or their products and occurs in dendritic cells, monocytes, macrophages, and T cells [152]. The characteristic cell death pattern of pyroptosis also includes cell swelling, plasma membrane damage, and massive cytoplasmic leakage, particularly of IL-1β [153]. It has been reported that pyroptosis usually results from the activation of inflammatory cystathionase, resulting in gasdermin D protein hydrolysis cleavage [154].
Melittin, an isolated water-soluble peptide derived from honeybee venom, is used to alleviate chronic inflammation [155]. Apatinib, in the presence of melittin, induces recruitment and activation of inflammatory vesicles and leads to pyroptosis and enhanced antitumor effects of apatinib [156]. This suggests that apatinib in mesenchymal or invasive TC shows promising therapeutic benefits. Furthermore, low-dose apatinib synergistically achieves comparable therapeutic potential with melittin, thereby reducing adverse events. The positive feedback modulation may improve the therapeutic efficacy of antiangiogenic targeted agents, offering new prospects for targeted therapy [156].
AIT is a classic, organ-specific autoimmune disease. Epidemiological investigations have shown that environmental triggers and genetic susceptibility contribute to decreased tolerance and disease progression [157]. Cytokine secretion and release from thyroid follicular cells are mediated by enhanced AIM2, NLRC4, and NLRP1, and NLRP3 inflammasomes are related to autoimmune thyroiditis [158].
Hashimoto’s thyroiditis (HT) is a chronic form of autoimmune thyroiditis. The main manifestations of HT are autoimmune hypothyroidism and lymphocyte infiltration of the thyroid tissue [15]. Excessive iodine intake is a major risk factor for HT [159]. In the NOD-H-2h4 mouse model (spontaneous autoimmune thyroiditis model) [160], excessive iodine induces thyroid follicular cell scorch death and the production of unbalanced reactive oxygen species (ROS) in a mouse model, thereby inducing autoimmune thyroiditis via ROS [161]. This reveals a new cellular mechanism of abnormal cell scorching death in HT, thus contributing to our understanding of the mechanisms involved in the occurrence of scorching death and providing further insight into the HT mechanism.
Subacute thyroiditis (SAT) is a self-limiting inflammatory thyroid disease [162]. SAT is caused by the destruction of thyroid follicles, leading to the leakage of stored colloids, which results in thyrotoxicosis and elevated sedimentation rate with discomfort and fever [163]. For many years, SAT has been a rare disease that has been treated with NSAIDs or corticosteroids [164]. In recent years, there has been hope to devise a novel approach to analgesia, antiviral therapy, inflammation reduction, and the use of hormones without hormone dependence. Lidocaine is the original antiarrhythmic drug [165]. Some researchers have suggested that lidocaine treats subacute thyroiditis by inhibiting the pyroptosis pathway to inhibit adenovirus-induced apoptosis of thyroid follicular epithelial cells [166].
5. Other Non-Apoptotic Cell Death Mechanisms and Thyroid Disease
ICD is another NAPCD induced by certain chemotherapeutic agents, lytic viruses, physical chemotherapy, photodynamic therapy, and radiation therapy [37].
The coatomer protein complex zeta 1 (COPZ1) is involved in the retrograde transport of proteins in the endoplasmic reticulum Golgi secretory pathway [167] and lipid homeostasis [168]. COPZ1-deficiency cells have been found to initiate IFN/viral mimicry responses, and ICD, in turn, exacerbates inflammation and cell death. The link between ICD and the type-I interferon pathway is well established [169]. COPZ1 deficiency triggers type-I IFN responses and immunogenic cell death in thyroid tumor cells [170]. Immunotherapy, represented by immune checkpoint blockade, has changed the cancer treatment paradigm. The immune co-inhibitory receptor (CIR) and its corresponding ligands are essential components of the tumor microenvironment [171]. T-cell immunoglobulin and mucin-domain containing-3 (TIM-3), T-cell immunoglobulin and ITIM domain (TIGIT), lymphocyte activation gene-3 (LAG-3), cytotoxic T-lymphocyte antigen 4 (CTLA-4), and Protein-1 (PD-1) are considered to be the major immune CIRs and the most promising immunotherapeutic targets in cancer therapy [172].
MTC is a relatively rare malignant neuroendocrine tumor that exhibits aggressive clinical progression [173]. In a large cohort study of MTC, positive TIGIT, LAG-3, CTLA-4, PD-1, and TIM-3 expression was detected in 6 (3.0 %), 6 (3.0 %), 25 (12.5 %), 27 (13.5 %), and 96 (48.0%) patients, respectively, with a positive correlation between TIM-3, PD-1, and CTLA-4 expression. This suggests that TIM-3, CTLA-4 positivity, and PD1/PD-L1 co-positivity may be potential immune features related to structural tumor recurrence [174].
6. Summary
Thyroid disorders, a hot spot among endocrine-related diseases, have been extensively studied in recent years. We have observed that NAPCD has excellent potential for diagnosing and managing thyroid diseases. In our study, we have reviewed recently published studies related to NAPCD and thyroid-related diseases to further elucidate the relationship between autophagy, ferroptosis, pyroptosis, ICD, and the development and progression of thyroid-related diseases (Table 1). These studies have laid the theoretical foundation for clarifying the mechanisms of disease development, searching for new therapeutic agents, identifying drug resistance mechanisms, and exploring targets. With the continuous exploration of the role of NAPCD in diseases, we have a better understanding of benign and malignant thyroid diseases. NAPCD plays a distinct role in the pathogenesis of thyroid tumors. Its use to effectively inhibit cancer cell proliferation and achieve precise treatment of thyroid-related diseases deserves further study by the academic community. In addition, we found that NAPCD may be associated with tumor drug resistance, which may help provide a new research direction for us to explore the mechanism of reversing drug resistance. Furthermore, elucidating the role of NAPCD in thyroid disease will help us explore novel drugs and targets. Consequently, exploring the underlying mechanisms in thyroid tumors and inflammation-related diseases has helped us better understand the self-mechanisms of NAPCD.
Author Contributions
F.J. contributed to the conception and study design, conducted the experiment, collected data, analyzed the data, and drafted the manuscript. X.Q. was responsible for revising and correcting the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the Thermal Ablation of Thyroid Nodules International Joint Laboratory (Henan Province) (YUKEWAI [2016] NO.11).
Institutional Review Board Statement
The study, carried out in accordance with the Declaration of Principles of Helsinki, won approval from the Ethics Committee of First Affiliated Hospital of Zhengzhou University. And the license number of the ethical review for the study is 2021-KY-0202-002.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Acknowledgments
We acknowledge assistance with the access of analytic instruments from the Translational Medical Center at The First Affiliated Hospital of Zhengzhou University.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
non-apoptotic programmed cell death (NAPCD); thyroid cancer (TC); papillary thyroid cancer (PTC); follicular thyroid cancer (FTC); medullary thyroid cancer (MTC); radioactive iodine (RAI, 131I); tyrosine kinase inhibitors (TKIs); autoimmune thyroiditis (AIT); Graves’ disease (GD); Hashimoto’s thyroiditis (HT); programmed cell death (PCD); accidental cell death (ACD); immunogenic cell death (ICD); Wilms’s tumor 1 (WT1)2; Sequestosome 1 (SQSTM1); glucose transporter protein 1 (GLUT1); phospholysine phosphohistidine inorganic pyrophosphate phosphatase (LHPP); sonic hedgehog (Shh); TGF-β-activated kinase (TAK1); Epithelial-mesenchymal transition (EMT); Baculoviral IAP repeat-containing 7 (BIRC7); Lactate dehydrogenase A (LDHA); Forkhead Box (FOX); RNA binding motif protein 47 (RBM47); small nucleolar RNA host gene 5 (SNHG5); transcription factor E3 (TFE3); microphthalmia/transcription factor E (MiT/TFE); death-associated protein kinase 2 (DAPK2); non-coding RNAs (ncRNAs); long non-coding RNAs (lncRNAs); distal-less homeobox 6 antisense RNA 1 (DLX6-AS1); homeobox A1 (HOXA1); serine/threonine kinase 4 (STK4); Activating transcription factor-2 (ATF2); growth arrest-specific 8 (GAS8); microRNAs (miRNAs); Prunella vulgaris L. (PV); adenosine monophos-phate-activated protein kinase (AMPK); unc-51-like autoph-agy-activated kinase 1 (ULK1); sodium iodide synthetics (NIS); radioactive iodine (RAI); vascular endothelial growth factor receptor 2 (VEGFR2); Di-isonylphthalate (DINP); Flavokawain BLHPP (FKB); glutathione peroxidase 4 (GPX4); arachidonic acid 5-lipoxygenase-activating protein (ALOX5AP); B-cell CLL/lymphoma 3 (BCL3); apolipoprotein E (APOE); damage-associated molecular patterns (DAMPs); Hashimoto’s thyroiditis (HT); reactive oxygen species (ROS); Subacute thyroiditis (SAT); coatomer protein complex zeta 1 (COPZ1); T-cell immunoglobulin and mucin-domain containing-3 (TIM-3); T-cell immunoglobulin and ITIM domain (TIGIT); lymphocyte activation gene-3 (LAG-3); cytotoxic T-lymphocyte antigen 4 (CTLA-4); Protein-1 (PD-1).
References
- Colin, I.M.; Denef, J.F.; Lengele, B.; Many, M.C.; Gerard, A.C. Recent insights into the cell biology of thyroid angiofollicular units. Endocr. Rev. 2013, 34, 209–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigneri, R.; Malandrino, P.; Vigneri, P. The changing epidemiology of thyroid cancer: Why is incidence increasing? Curr. Opin. Oncol. 2015, 27, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fagin, J.A.; Wells, S.A., Jr. Biologic and Clinical Perspectives on Thyroid Cancer. N. Engl. J. Med. 2016, 375, 1054–1067. [Google Scholar] [CrossRef] [Green Version]
- Carling, T.; Udelsman, R. Thyroid cancer. Annu Rev. Med. 2014, 65, 125–137. [Google Scholar] [CrossRef]
- Barletta, J.A.; Nose, V.; Sadow, P.M. Genomics and Epigenomics of Medullary Thyroid Carcinoma: From Sporadic Disease to Familial Manifestations. Endocr. Pathol. 2021, 32, 35–43. [Google Scholar] [CrossRef]
- Nelkin, B. Recent advances in the biology and therapy of medullary thyroid carcinoma. F1000Res 2017, 6, 2184. [Google Scholar] [CrossRef] [Green Version]
- Cote, G.J.; Grubbs, E.G.; Hofmann, M.C. Thyroid C-Cell Biology and Oncogenic Transformation. Recent Results Cancer Res. 2015, 204, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Durante, C.; Haddy, N.; Baudin, E.; Leboulleux, S.; Hartl, D.; Travagli, J.P.; Caillou, B.; Ricard, M.; Lumbroso, J.D.; De Vathaire, F.; et al. Long-term outcome of 444 patients with distant metastases from papillary and follicular thyroid carcinoma: Benefits and limits of radioiodine therapy. J. Clin. Endocrinol. Metab. 2006, 91, 2892–2899. [Google Scholar] [CrossRef]
- Ibrahimpasic, T.; Ghossein, R.; Carlson, D.L.; Nixon, I.; Palmer, F.L.; Shaha, A.R.; Patel, S.G.; Tuttle, R.M.; Shah, J.P.; Ganly, I. Outcomes in patients with poorly differentiated thyroid carcinoma. J. Clin. Endocrinol. Metab. 2014, 99, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.P.; Shaha, A.R. Anaplastic thyroid cancer. Oral. Oncol. 2013, 49, 702–706. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.A.; Kuo, J.H. Advancements in the treatment of differentiated thyroid cancer. Ther. Adv. Endocrinol. Metab. 2021, 12, 20420188211000251. [Google Scholar] [CrossRef] [PubMed]
- Gild, M.L.; Tsang, V.H.M.; Clifton-Bligh, R.J.; Robinson, B.G. Multikinase inhibitors in thyroid cancer: Timing of targeted therapy. Nat. Rev. Endocrinol. 2021, 17, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Ferrari, S.M.; Corrado, A.; Di Domenicantonio, A.; Fallahi, P. Autoimmune thyroid disorders. Autoimmun. Rev. 2015, 14, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Chen, L.; Lian, X.; Liu, C.; Shi, B.; Shi, L.; Tong, N.; Wang, S.; Weng, J.; Zhao, J.; et al. Iodine Status and Prevalence of Thyroid Disorders After Introduction of Mandatory Universal Salt Iodization for 16 Years in China: A Cross-Sectional Study in 10 Cities. Thyroid 2016, 26, 1125–1130. [Google Scholar] [CrossRef]
- Liu, H.; Zheng, T.; Mao, Y.; Xu, C.; Wu, F.; Bu, L.; Mou, X.; Zhou, Y.; Yuan, G.; Wang, S.; et al. gammadelta Tau cells enhance B cells for antibody production in Hashimoto’s thyroiditis, and retinoic acid induces apoptosis of the gammadelta Tau cell. Endocrine 2016, 51, 113–122. [Google Scholar] [CrossRef]
- Hollowell, J.G.; Staehling, N.W.; Flanders, W.D.; Hannon, W.H.; Gunter, E.W.; Spencer, C.A.; Braverman, L.E. Serum TSH, T(4), and thyroid antibodies in the United States population (1988 to 1994): National Health and Nutrition Examination Survey (NHANES III). J. Clin. Endocrinol. Metab. 2002, 87, 489–499. [Google Scholar] [CrossRef]
- Sawicka-Gutaj, N.; Gruszczynski, D.; Zawalna, N.; Nijakowski, K.; Muller, I.; Karpinski, T.; Salvi, M.; Ruchala, M. Microbiota Alterations in Patients with Autoimmune Thyroid Diseases: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 13450. [Google Scholar] [CrossRef]
- Ragusa, F.; Fallahi, P.; Elia, G.; Gonnella, D.; Paparo, S.R.; Giusti, C.; Churilov, L.P.; Ferrari, S.M.; Antonelli, A. Hashimotos’ thyroiditis: Epidemiology, pathogenesis, clinic and therapy. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101367. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Vandenabeele, P.; Abrams, J.; Alnemri, E.S.; Baehrecke, E.H.; Blagosklonny, M.V.; El-Deiry, W.S.; Golstein, P.; Green, D.R.; et al. Classification of cell death: Recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ. 2009, 16, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daley-Bauer, L.P.; Roback, L.; Crosby, L.N.; McCormick, A.L.; Feng, Y.; Kaiser, W.J.; Mocarski, E.S. Mouse cytomegalovirus M36 and M45 death suppressors cooperate to prevent inflammation resulting from antiviral programmed cell death pathways. Proc. Natl. Acad. Sci. USA 2017, 114, E2786–E2795. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, Y.; Steller, H. Live to die another way: Modes of programmed cell death and the signals emanating from dying cells. Nat. Rev. Mol. Cell Biol. 2015, 16, 329–344. [Google Scholar] [CrossRef]
- Ke, B.; Tian, M.; Li, J.; Liu, B.; He, G. Targeting Programmed Cell Death Using Small-Molecule Compounds to Improve Potential Cancer Therapy. Med. Res. Rev. 2016, 36, 983–1035. [Google Scholar] [CrossRef]
- Hartman, M.L. Non-Apoptotic Cell Death Signaling Pathways in Melanoma. Int. J. Mol. Sci. 2020, 21, 2980. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Liu, J.; Kang, R.; Klionsky, D.J.; Kroemer, G.; Tang, D. Ferroptosis is a type of autophagy-dependent cell death. Semin. Cancer Biol. 2020, 66, 89–100. [Google Scholar] [CrossRef]
- Wu, J.; Ye, J.; Kong, W.; Zhang, S.; Zheng, Y. Programmed cell death pathways in hearing loss: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2020, 53, e12915. [Google Scholar] [CrossRef]
- Ye, J.; Zhang, R.; Wu, F.; Zhai, L.; Wang, K.; Xiao, M.; Xie, T.; Sui, X. Non-apoptotic cell death in malignant tumor cells and natural compounds. Cancer Lett. 2018, 420, 210–227. [Google Scholar] [CrossRef]
- Denton, D.; Kumar, S. Autophagy-dependent cell death. Cell Death Differ. 2019, 26, 605–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushima, N.; Levine, B. Autophagy in Human Diseases. N. Engl. J. Med. 2020, 383, 1564–1576. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Wang, J.; Wu, J.; He, D.; Zhang, C.; Duan, C.; Li, B. Ferroptosis, a new form of cell death: Opportunities and challenges in cancer. J. Hematol. Oncol. 2019, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beier, J.I.; Banales, J.M. Pyroptosis: An inflammatory link between NAFLD and NASH with potential therapeutic implications. J. Hepatol. 2018, 68, 643–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, I.; Miao, E.A. Pyroptotic cell death defends against intracellular pathogens. Immunol. Rev. 2015, 265, 130–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Tait, S.W.G. Targeting immunogenic cell death in cancer. Mol. Oncol. 2020, 14, 2994–3006. [Google Scholar] [CrossRef]
- Garg, A.D.; Galluzzi, L.; Apetoh, L.; Baert, T.; Birge, R.B.; Bravo-San Pedro, J.M.; Breckpot, K.; Brough, D.; Chaurio, R.; Cirone, M.; et al. Molecular and Translational Classifications of DAMPs in Immunogenic Cell Death. Front. Immunol. 2015, 6, 588. [Google Scholar] [CrossRef] [Green Version]
- Cetti, E.; Di Marco, T.; Mauro, G.; Mazzoni, M.; Lecis, D.; Minna, E.; Gioiosa, L.; Brich, S.; Pagliardini, S.; Borrello, M.G.; et al. Mitosis perturbation by MASTL depletion impairs the viability of thyroid tumor cells. Cancer Lett. 2019, 442, 362–372. [Google Scholar] [CrossRef]
- Varinelli, L.; Caccia, D.; Volpi, C.C.; Caccia, C.; De Bortoli, M.; Taverna, E.; Gualeni, A.V.; Leoni, V.; Gloghini, A.; Manenti, G.; et al. 4-IPP, a selective MIF inhibitor, causes mitotic catastrophe in thyroid carcinomas. Endocr. Relat. Cancer 2015, 22, 759–775. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, T.; Yoshimori, T. Autophagosome biogenesis and human health. Cell Discov. 2020, 6, 33. [Google Scholar] [CrossRef]
- Valencia, M.; Kim, S.R.; Jang, Y.; Lee, S.H. Neuronal Autophagy: Characteristic Features and Roles in Neuronal Pathophysiology. Biomol. Ther. 2021, 29, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Chmurska, A.; Matczak, K.; Marczak, A. Two Faces of Autophagy in the Struggle against Cancer. Int. J. Mol. Sci. 2021, 22, 2981. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V. Autophagy in inflammation, infection, and immunometabolism. Immunity 2021, 54, 437–453. [Google Scholar] [CrossRef]
- Chen, C.; Gao, H.; Su, X. Autophagy-related signaling pathways are involved in cancer (Review). Exp. Ther. Med. 2021, 22, 710. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in major human diseases. EMBO J. 2021, 40, e108863. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.R.; Thorburn, A. Autophagy and organelle homeostasis in cancer. Dev. Cell 2021, 56, 906–918. [Google Scholar] [CrossRef] [PubMed]
- Bialik, S.; Dasari, S.K.; Kimchi, A. Autophagy-dependent cell death—Where, how and why a cell eats itself to death. J. Cell Sci. 2018, 131, jcs215152. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.Q.; Kumar, A.V.; Mills, J.; Lapierre, L.R. Autophagy in aging and longevity. Hum. Genet. 2020, 139, 277–290. [Google Scholar] [CrossRef]
- Yamazaki, T.; Bravo-San Pedro, J.M.; Galluzzi, L.; Kroemer, G.; Pietrocola, F. Autophagy in the cancer-immunity dialogue. Adv. Drug Deliv. Rev. 2021, 169, 40–50. [Google Scholar] [CrossRef]
- Xing, M. BRAF mutation in papillary thyroid cancer: Pathogenic role, molecular bases, and clinical implications. Endocr. Rev. 2007, 28, 742–762. [Google Scholar] [CrossRef]
- Cracolici, V.; Mujacic, I.; Kadri, S.; Alikhan, M.; Niu, N.; Segal, J.P.; Rosen, L.E.; Sarne, D.H.; Morgan, A.; Desouky, S.; et al. Synchronous and Metastatic Papillary and Follicular Thyroid Carcinomas with Unique Molecular Signatures. Endocr. Pathol. 2018, 29, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, X.L.; Zhang, Y.F.; Wang, D.; Wang, Q.; Zhao, C.K.; Li, M.X.; Wei, Q.; Ji, G.; Xu, H.X. Ultrasound validation of predictive model for central cervical lymph node metastasis in papillary thyroid cancer on BRAF. Future Oncol. 2020, 16, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Eozenou, C.; Gonen, N.; Touzon, M.S.; Jorgensen, A.; Yatsenko, S.A.; Fusee, L.; Kamel, A.K.; Gellen, B.; Guercio, G.; Singh, P.; et al. Testis formation in XX individuals resulting from novel pathogenic variants in Wilms’ tumor 1 (WT1) gene. Proc. Natl. Acad. Sci. USA 2020, 117, 13680–13688. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lin, S.; Lin, Y.; Wu, S.; Zhuo, M.; Zhang, A.; Zheng, J.; You, Z. BRAF-activated WT1 contributes to cancer growth and regulates autophagy and apoptosis in papillary thyroid carcinoma. J. Transl. Med. 2022, 20, 79. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, M.D.; Galic, S.; Marcinko, K.; Sikkema, S.; Pulinilkunnil, T.; Chen, Z.P.; O’Neill, H.M.; Ford, R.J.; Palanivel, R.; O’Brien, M.; et al. Single phosphorylation sites in Acc1 and Acc2 regulate lipid homeostasis and the insulin-sensitizing effects of metformin. Nat. Med. 2013, 19, 1649–1654. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Hou, L.; Song, H.; Xu, P.; Sun, Y.; Wu, K. Akt/AMPK/mTOR pathway was involved in the autophagy induced by vitamin E succinate in human gastric cancer SGC-7901 cells. Mol. Cell Biochem. 2017, 424, 173–183. [Google Scholar] [CrossRef]
- Sun, W.; Qian, K.; Guo, K.; Chen, L.; Xiang, J.; Li, D.; Wu, Y.; Ji, Q.; Sun, T.; Wang, Z. LHPP inhibits cell growth and migration and triggers autophagy in papillary thyroid cancer by regulating the AKT/AMPK/mTOR signaling pathway. Acta Biochim. Biophys. Sin. 2020, 52, 382–389. [Google Scholar] [CrossRef]
- Liang, Z.; Yuan, Z.; Guo, J.; Wu, J.; Yi, J.; Deng, J.; Shan, Y. Ganoderma lucidum Polysaccharides Prevent Palmitic Acid-Evoked Apoptosis and Autophagy in Intestinal Porcine Epithelial Cell Line via Restoration of Mitochondrial Function and Regulation of MAPK and AMPK/Akt/mTOR Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 478. [Google Scholar] [CrossRef] [Green Version]
- Waters, S.; Marchbank, K.; Solomon, E.; Whitehouse, C.A. Autophagic receptors Nbr1 and p62 coregulate skeletal remodeling. Autophagy 2010, 6, 981–983. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Ma, R.; Liu, C.; Zhang, L.; Feng, K.; Wang, M.; Yin, D. SQSTM1/p62 Promotes Cell Growth and Triggers Autophagy in Papillary Thyroid Cancer by Regulating the AKT/AMPK/mTOR Signaling Pathway. Front Oncol. 2021, 11, 638701. [Google Scholar] [CrossRef]
- Cui, L.; Gong, X.; Tang, Y.; Kong, L.; Chang, M.; Geng, H.; Xu, K.; Wang, F. Relationship between the LHPP Gene Polymorphism and Resting-State Brain Activity in Major Depressive Disorder. Neural Plast. 2016, 2016, 9162590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, X.; Zhou, X.; Zhang, X. LHPP suppresses bladder cancer cell proliferation and growth via inactivating AKT/p65 signaling pathway. Biosci. Rep. 2019, 39, BSR20182270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeng, K.S.; Chang, C.F.; Lin, S.S. Sonic Hedgehog Signaling in Organogenesis, Tumors, and Tumor Microenvironments. Int. J. Mol. Sci. 2020, 21, 758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, J.; Lu, Y.; Zhao, Y.; Prinz, R.A.; Xu, X. Inhibition of the sonic hedgehog pathway activates TGF-beta-activated kinase (TAK1) to induce autophagy and suppress apoptosis in thyroid tumor cells. Cell Death Dis. 2021, 12, 459. [Google Scholar] [CrossRef]
- Liang, J.; Li, X.; Li, Y.; Wei, J.; Daniels, G.; Zhong, X.; Wang, J.; Sfanos, K.; Melamed, J.; Zhao, J.; et al. LEF1 targeting EMT in prostate cancer invasion is mediated by miR-181a. Am. J. Cancer Res. 2015, 5, 1124–1132. [Google Scholar]
- Baum, B.; Settleman, J.; Quinlan, M.P. Transitions between epithelial and mesenchymal states in development and disease. Semin. Cell Dev. Biol. 2008, 19, 294–308. [Google Scholar] [CrossRef]
- Liu, K.; Yu, Q.; Li, H.; Xie, C.; Wu, Y.; Ma, D.; Sheng, P.; Dai, W.; Jiang, H. BIRC7 promotes epithelial-mesenchymal transition and metastasis in papillary thyroid carcinoma through restraining autophagy. Am. J. Cancer Res. 2020, 10, 78–94. [Google Scholar]
- Hou, X.; Shi, X.; Zhang, W.; Li, D.; Hu, L.; Yang, J.; Zhao, J.; Wei, S.; Wei, X.; Ruan, X.; et al. LDHA induces EMT gene transcription and regulates autophagy to promote the metastasis and tumorigenesis of papillary thyroid carcinoma. Cell Death Dis. 2021, 12, 347. [Google Scholar] [CrossRef]
- Calissi, G.; Lam, E.W.; Link, W. Therapeutic strategies targeting FOXO transcription factors. Nat. Rev. Drug Discov. 2021, 20, 21–38. [Google Scholar] [CrossRef]
- Chen, W.; Jiang, J.; Gong, L.; Shu, Z.; Xiang, D.; Zhang, X.; Bi, K.; Diao, H. Hepatitis B virus P protein initiates glycolytic bypass in HBV-related hepatocellular carcinoma via a FOXO3/miRNA-30b-5p/MINPP1 axis. J. Exp. Clin. Cancer Res. 2021, 40, 1. [Google Scholar] [CrossRef]
- Zhao, M.; Gao, J.; Cui, C.; Zhang, Y.; Jiang, X.; Cui, J. Inhibition of PTEN Ameliorates Secondary Hippocampal Injury and Cognitive Deficits after Intracerebral Hemorrhage: Involvement of AKT/FoxO3a/ATG-Mediated Autophagy. Oxid. Med. Cell Longev. 2021, 2021, 5472605. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Sun, W.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Lv, C.; Zhang, H. RBM47/SNHG5/FOXO3 axis activates autophagy and inhibits cell proliferation in papillary thyroid carcinoma. Cell Death Dis. 2022, 13, 270. [Google Scholar] [CrossRef] [PubMed]
- Nestal de Moraes, G.; Carneiro, L.D.T.; Maia, R.C.; Lam, E.W.; Sharrocks, A.D. FOXK2 Transcription Factor and Its Emerging Roles in Cancer. Cancers 2019, 11, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, P.; Ju, H.; Zhu, T.; Shi, J.; Huang, Y. FOXK2 promotes the proliferation of papillary thyroid cancer cell by down-regulating autophagy. J. Cancer 2022, 13, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Shen, L.D.; Li, K.; Yang, R.X.; Zhang, Q.Y.; Chen, Y.; Gao, C.L.; Dong, C.; Bi, Q.; Tao, J.N.; et al. Inhibition of livin expression suppresses cell proliferation and enhances chemosensitivity to cisplatin in human lung adenocarcinoma cells. Mol. Med. Rep. 2015, 12, 547–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Yang, D.; Wang, S.; Che, X.; Wang, J.; Li, X.; Zhang, Z.; Chen, X.; Song, X. Livin regulates prostate cancer cell invasion by impacting the NF-kappaB signaling pathway and the expression of FN and CXCR4. IUBMB Life 2012, 64, 274–283. [Google Scholar] [CrossRef]
- Perera, R.M.; Stoykova, S.; Nicolay, B.N.; Ross, K.N.; Fitamant, J.; Boukhali, M.; Lengrand, J.; Deshpande, V.; Selig, M.K.; Ferrone, C.R.; et al. Transcriptional control of autophagy-lysosome function drives pancreatic cancer metabolism. Nature 2015, 524, 361–365. [Google Scholar] [CrossRef]
- Lu, H.; Zhu, C.; Ruan, Y.; Fan, L.; Wei, K.; Yang, Z.; Chen, Q. TFE3 Regulates the Function of the Autophagy-Lysosome Pathway to Drive the Invasion and Metastasis of Papillary Thyroid Carcinoma. Anal. Cell Pathol. 2021, 2021, 3081491. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, J.; Xu, H.; Zhou, X.; He, L.; Zhu, C. DAPK2 activates NF-kappaB through autophagy-dependent degradation of I-kappaBalpha during thyroid cancer development and progression. Ann. Transl. Med. 2021, 9, 1083. [Google Scholar] [CrossRef]
- Consortium, E.P.; Birney, E.; Stamatoyannopoulos, J.A.; Dutta, A.; Guigo, R.; Gingeras, T.R.; Margulies, E.H.; Weng, Z.; Snyder, M.; Dermitzakis, E.T.; et al. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature 2007, 447, 799–816. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Dutta, A. MicroRNAs in cancer. Annu. Rev. Pathol. 2009, 4, 199–227. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.; Corey, D.R. Non-coding RNAs as drug targets. Nat. Rev. Drug Discov. 2017, 16, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K.; Miyoshi, T.; Kugou, K.; Hoffman, C.S.; Shibata, T.; Ohta, K. Stepwise chromatin remodelling by a cascade of transcription initiation of non-coding RNAs. Nature 2008, 456, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Gulia, C.; Baldassarra, S.; Signore, F.; Rigon, G.; Pizzuti, V.; Gaffi, M.; Briganti, V.; Porrello, A.; Piergentili, R. Role of Non-Coding RNAs in the Etiology of Bladder Cancer. Genes 2017, 8, 339. [Google Scholar] [CrossRef] [Green Version]
- Batista, P.J.; Chang, H.Y. Long noncoding RNAs: Cellular address codes in development and disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [Green Version]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Li, W.; Zhang, B.; Liu, X. Overexpression of lncRNA SLC26A4-AS1 inhibits papillary thyroid carcinoma progression through recruiting ETS1 to promote ITPR1-mediated autophagy. J. Cell Mol. Med. 2021, 25, 8148–8158. [Google Scholar] [CrossRef]
- Feng, L.; Wang, R.; Wang, Y.; Shen, X.; Shi, Q.; Lian, M.; Ma, H.; Fang, J. Silencing long non-coding RNA DLX6-AS1 or restoring microRNA-193b-3p enhances thyroid carcinoma cell autophagy and apoptosis via depressing HOXA1. J. Cell Mol. Med. 2021, 25, 9319–9330. [Google Scholar] [CrossRef]
- Peng, X.; Ji, C.; Tan, L.; Lin, S.; Zhu, Y.; Long, M.; Luo, D.; Li, H. Long non-coding RNA TNRC6C-AS1 promotes methylation of STK4 to inhibit thyroid carcinoma cell apoptosis and autophagy via Hippo signalling pathway. J. Cell Mol. Med. 2020, 24, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Sun, W.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Shao, L.; Zhang, H. ATF2-Induced lncRNA GAS8-AS1 Promotes Autophagy of Thyroid Cancer Cells by Targeting the miR-187-3p/ATG5 and miR-1343-3p/ATG7 Axes. Mol. Ther. Nucleic Acids 2020, 22, 584–600. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, L.; Li, J.; Zhong, L. Silencing of long noncoding RNA RP11-476D10.1 enhances apoptosis and autophagy while inhibiting proliferation of papillary thyroid carcinoma cells via microRNA-138-5p-dependent inhibition of LRRK2. J. Cell Physiol. 2019, 234, 20980–20991. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Liu, W.L.; Lu, Z.W.; Cao, Y.M.; Ji, Q.H.; Wei, W.J. SNHG9, a Papillary Thyroid Cancer Cell Exosome-Enriched lncRNA, Inhibits Cell Autophagy and Promotes Cell Apoptosis of Normal Thyroid Epithelial Cell Nthy-ori-3 Through YBOX3/P21 Pathway. Front Oncol. 2021, 11, 647034. [Google Scholar] [CrossRef] [PubMed]
- Ali Syeda, Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, X.; Lin, T.; Chen, X.; Yan, J.; Jiang, S. MicroRNA-524-5p suppresses the progression of papillary thyroid carcinoma cells via targeting on FOXE1 and ITGA3 in cell autophagy and cycling pathways. J. Cell Physiol. 2019, 234, 18382–18391. [Google Scholar] [CrossRef] [Green Version]
- Gordaliza, M. Natural products as leads to anticancer drugs. Clin. Transl. Oncol. 2007, 9, 767–776. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. The influence of natural products upon drug discovery. Nat. Prod Rep. 2000, 17, 215–234. [Google Scholar] [CrossRef] [Green Version]
- Yallapu, M.M.; Jaggi, M.; Chauhan, S.C. Curcumin nanomedicine: A road to cancer therapeutics. Curr. Pharm. Des. 2013, 19, 1994–2010. [Google Scholar] [CrossRef]
- Howells, L.; Malhotra Mukhtyar, R.; Theofanous, D.; Pepper, C.; Thomas, A.; Brown, K.; Khan, S. A Systematic Review Assessing Clinical Utility of Curcumin with a Focus on Cancer Prevention. Mol. Nutr. Food Res. 2021, 65, e2000977. [Google Scholar] [CrossRef]
- Henrotin, Y.; Dierckxsens, Y.; Delisse, G.; Maes, N.; Albert, A. Curcuma longa and Boswellia serrata extract combination for hand osteoarthritis: An open-label pre-post trial. Pharm. Biol. 2022, 60, 2295–2299. [Google Scholar] [CrossRef]
- Sangouni, A.A.; Taghdir, M.; Mirahmadi, J.; Sepandi, M.; Parastouei, K. Effects of curcumin and/or coenzyme Q10 supplementation on metabolic control in subjects with metabolic syndrome: A randomized clinical trial. Nutr. J. 2022, 21, 62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, S.; Cheng, X.; Wu, J.; Wu, L.; Wang, Y.; Wang, X.; Bao, J.; Yu, H. Curcumin induces autophagic cell death in human thyroid cancer cells. Toxicol. In Vitro 2022, 78, 105254. [Google Scholar] [CrossRef] [PubMed]
- Xing, M. Molecular pathogenesis and mechanisms of thyroid cancer. Nat. Rev. Cancer 2013, 13, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Valerio, L.; Pieruzzi, L.; Giani, C.; Agate, L.; Bottici, V.; Lorusso, L.; Cappagli, V.; Puleo, L.; Matrone, A.; Viola, D.; et al. Targeted Therapy in Thyroid Cancer: State of the Art. Clin. Oncol. R Coll Radiol. 2017, 29, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Crispo, F.; Notarangelo, T.; Pietrafesa, M.; Lettini, G.; Storto, G.; Sgambato, A.; Maddalena, F.; Landriscina, M. BRAF Inhibitors in Thyroid Cancer: Clinical Impact, Mechanisms of Resistance and Future Perspectives. Cancers 2019, 11, 1388. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Wang, C.; Gao, W.; Cui, R.; Liang, J. Overwhelming rapid metabolic and structural response to apatinib in radioiodine refractory differentiated thyroid cancer. Oncotarget 2017, 8, 42252–42261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Wang, H.; Zhao, J.; Hu, L.; Zhi, J.; Wei, S.; Ruan, X.; Hou, X.; Li, D.; Zhang, J.; et al. Apatinib Inhibits Cell Proliferation and Induces Autophagy in Human Papillary Thyroid Carcinoma via the PI3K/Akt/mTOR Signaling Pathway. Front Oncol. 2020, 10, 217. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Cheng, X.; Kuang, J.; Chen, L.; Yuen, S.; Shi, M.; Liang, J.; Shen, B.; Jin, Z.; Yan, J.; et al. Apatinib-induced protective autophagy and apoptosis through the AKT-mTOR pathway in anaplastic thyroid cancer. Cell Death Dis. 2018, 9, 1030. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Cheng, X.; Feng, H.; Kuang, J.; Yang, W.; Peng, C.; Shen, B.; Qiu, W. Apatinib Inhibits Angiogenesis Via Suppressing Akt/GSK3beta/ANG Signaling Pathway in Anaplastic Thyroid Cancer. Cell Physiol. Biochem. 2017, 44, 1471–1484. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, K. Biological Activities of Red Pepper (Capsicum annuum) and Its Pungent Principle Capsaicin: A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 1488–1500. [Google Scholar] [CrossRef]
- Wu, L.; Xu, S.; Cheng, X.; Zhang, L.; Wang, Y.; Wu, J.; Bao, J.; Yu, H.; Lu, R. Capsaicin inhibits the stemness of anaplastic thyroid carcinoma cells by triggering autophagy-lysosome mediated OCT4A degradation. Phytother. Res. 2022, 36, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Subbiah, V.; Baik, C.; Kirkwood, J.M. Clinical Development of BRAF plus MEK Inhibitor Combinations. Trends Cancer 2020, 6, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.M.; Centanni, M.; Virili, C.; Miccoli, M.; Ferrari, P.; Ruffilli, I.; Ragusa, F.; Antonelli, A.; Fallahi, P. Sunitinib in the Treatment of Thyroid Cancer. Curr. Med. Chem. 2019, 26, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Shi, X.; Dong, Q.; Yu, Y.; Hou, X.; Song, X.; Wei, X.; Chen, L.; Gao, M. Antitumor effects of anlotinib in thyroid cancer. Endocr. Relat. Cancer 2019, 26, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wu, Y.; Chen, L.; Hu, L.; Liu, X. Initial treatment combined with Prunella vulgaris reduced prednisolone consumption for patients with subacute thyroiditis. Ann. Transl. Med. 2019, 7, 45. [Google Scholar] [CrossRef]
- Carroll, B.; Dunlop, E.A. The lysosome: A crucial hub for AMPK and mTORC1 signalling. Biochem. J. 2017, 474, 1453–1466. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Zhang, Z.; Hu, Y.; Li, Z.; Wan, Y.; Liu, J.; Chu, X.; Wei, Q.; Zhao, M.; Yang, X. An aqueous extract of Prunella vulgaris L. inhibits the growth of papillary thyroid carcinoma by inducing autophagy in vivo and in vitro. Phytother. Res. 2021, 35, 2691–2702. [Google Scholar] [CrossRef]
- Leung, A.M.; Braverman, L.E. Consequences of excess iodine. Nat. Rev. Endocrinol. 2014, 10, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Speeckaert, M.M.; Speeckaert, R.; Wierckx, K.; Delanghe, J.R.; Kaufman, J.M. Value and pitfalls in iodine fortification and supplementation in the 21st century. Br. J. Nutr. 2011, 106, 964–973. [Google Scholar] [CrossRef] [Green Version]
- Guan, H.; Ji, M.; Bao, R.; Yu, H.; Wang, Y.; Hou, P.; Zhang, Y.; Shan, Z.; Teng, W.; Xing, M. Association of high iodine intake with the T1799A BRAF mutation in papillary thyroid cancer. J. Clin. Endocrinol. Metab. 2009, 94, 1612–1617. [Google Scholar] [CrossRef]
- Li, D.D.; Zhang, Y.F.; Xu, H.X.; Zhang, X.P. The role of BRAF in the pathogenesis of thyroid carcinoma. Front. Biosci. 2015, 20, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Park, H.K.; Byun, D.W.; Suh, K.; Yoo, M.H.; Min, Y.K.; Kim, S.W.; Chung, J.H. Iodine intake as a risk factor for BRAF mutations in papillary thyroid cancer patients from an iodine-replete area. Eur. J. Nutr. 2018, 57, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Xu, X.; Li, J.; Yang, X.; Sun, J.; Wu, Y.; Qiao, H. High iodine effects on the proliferation, apoptosis, and migration of papillary thyroid carcinoma cells as a result of autophagy induced by BRAF kinase. Biomed. Pharm. 2019, 120, 109476. [Google Scholar] [CrossRef] [PubMed]
- Kavlock, R.; Boekelheide, K.; Chapin, R.; Cunningham, M.; Faustman, E.; Foster, P.; Golub, M.; Henderson, R.; Hinberg, I.; Little, R.; et al. NTP Center for the Evaluation of Risks to Human Reproduction: Phthalates expert panel report on the reproductive and developmental toxicity of di-isononyl phthalate. Reprod Toxicol. 2002, 16, 679–708. [Google Scholar] [CrossRef]
- Duan, J.; Deng, T.; Kang, J.; Chen, M. DINP aggravates autoimmune thyroid disease through activation of the Akt/mTOR pathway and suppression of autophagy in Wistar rats. Environ. Pollut. 2019, 245, 316–324. [Google Scholar] [CrossRef]
- He, Q.; Liu, W.; Sha, S.; Fan, S.; Yu, Y.; Chen, L.; Dong, M. Adenosine 5′-monophosphate-activated protein kinase-dependent mTOR pathway is involved in flavokawain B-induced autophagy in thyroid cancer cells. Cancer Sci. 2018, 109, 2576–2589. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liu, X.; Jin, S.; Chen, Y.; Guo, R. Ferroptosis in cancer therapy: A novel approach to reversing drug resistance. Mol. Cancer 2022, 21, 47. [Google Scholar] [CrossRef]
- Nussinov, R.; Tsai, C.J.; Jang, H. Anticancer drug resistance: An update and perspective. Drug Resist. Updat. 2021, 59, 100796. [Google Scholar] [CrossRef]
- Smith, A.G.; Macleod, K.F. Autophagy, cancer stem cells and drug resistance. J. Pathol. 2019, 247, 708–718. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Jiang, L.; Miao, H.; Lv, Y.; Zhang, Q.; Ma, M.; Duan, W.; Huang, Y.; Wei, X. Autophagy regulation of ATG13 and ATG27 on biofilm formation and antifungal resistance in Candida albicans. Biofouling 2022, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, J.; Li, Q. Autophagy, an accomplice or antagonist of drug resistance in HCC? Cell Death Dis. 2021, 12, 266. [Google Scholar] [CrossRef]
- Chang, H.; Zou, Z. Targeting autophagy to overcome drug resistance: Further developments. J. Hematol. Oncol. 2020, 13, 159. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Song, D.; Yan, Y.; Huang, C.; Shen, C.; Lan, J.; Chen, Y.; Liu, A.; Wu, Q.; Sun, L.; et al. IL-6 regulates autophagy and chemotherapy resistance by promoting BECN1 phosphorylation. Nat. Commun. 2021, 12, 3651. [Google Scholar] [CrossRef] [PubMed]
- Amaravadi, R.K.; Kimmelman, A.C.; Debnath, J. Targeting Autophagy in Cancer: Recent Advances and Future Directions. Cancer Discov. 2019, 9, 1167–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Shong, Y.K.; Kim, T.Y.; Viola, D.; Elisei, R.; Bendlova, B.; Yip, L.; Mian, C.; et al. Association between BRAF V600E mutation and recurrence of papillary thyroid cancer. J. Clin. Oncol. 2015, 33, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklener, K.; Mazurek, M.; Wieteska, M.; Waclawska, M.; Bilski, M.; Mandziuk, S. New Directions in the Therapy of Glioblastoma. Cancers 2022, 14, 5377. [Google Scholar] [CrossRef]
- Pastwinska, J.; Karas, K.; Karwaciak, I.; Ratajewski, M. Targeting EGFR in melanoma—The sea of possibilities to overcome drug resistance. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188754. [Google Scholar] [CrossRef]
- Jimenez-Mora, E.; Gallego, B.; Diaz-Gago, S.; Lasa, M.; Baquero, P.; Chiloeches, A. (V600E)BRAF Inhibition Induces Cytoprotective Autophagy through AMPK in Thyroid Cancer Cells. Int. J. Mol. Sci. 2021, 22I, 6033. [Google Scholar] [CrossRef]
- Hassannia, B.; Vandenabeele, P.; Vanden Berghe, T. Targeting Ferroptosis to Iron Out Cancer. Cancer Cell 2019, 35, 830–849. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, Z.; Pan, K.; Li, J.; Chen, Q. The function and mechanism of ferroptosis in cancer. Apoptosis 2020, 25, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kang, R.; Kroemer, G.; Tang, D. Broadening horizons: The role of ferroptosis in cancer. Nat. Rev. Clin. Oncol. 2021, 18, 280–296. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.H.; Fu, X.H.; Li, G.Q.; He, Q.; Qiu, X.G. FTO Prevents Thyroid Cancer Progression by SLC7A11 m6A Methylation in a Ferroptosis-Dependent Manner. Front. Endocrinol. 2022, 13, 857765. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Fogarty, C.E.; Ma, B.; Li, H.; Ni, G.; Liu, X.; Yuan, J.; Wang, T. Identification of ferroptosis genes in immune infiltration and prognosis in thyroid papillary carcinoma using network analysis. BMC Genom. 2021, 22, 576. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhao, H. Ferroptosis-Related APOE, BCL3 and ALOX5AP Gene Polymorphisms are Associated with the Risk of Thyroid Cancer. Pharmgenomics Pers. Med. 2022, 15, 157–165. [Google Scholar] [CrossRef]
- Ngo, B.; Van Riper, J.M.; Cantley, L.C.; Yun, J. Targeting cancer vulnerabilities with high-dose vitamin C. Nat. Rev. Cancer 2019, 19, 271–282. [Google Scholar] [CrossRef]
- Wang, X.; Xu, S.; Zhang, L.; Cheng, X.; Yu, H.; Bao, J.; Lu, R. Vitamin C induces ferroptosis in anaplastic thyroid cancer cells by ferritinophagy activation. Biochem. Biophys. Res. Commun. 2021, 551, 46–53. [Google Scholar] [CrossRef]
- Fang, Y.; Tian, S.; Pan, Y.; Li, W.; Wang, Q.; Tang, Y.; Yu, T.; Wu, X.; Shi, Y.; Ma, P.; et al. Pyroptosis: A new frontier in cancer. Biomed. Pharm. 2020, 121, 109595. [Google Scholar] [CrossRef]
- De Vasconcelos, N.M.; Lamkanfi, M. Recent Insights on Inflammasomes, Gasdermin Pores, and Pyroptosis. Cold Spring Harb. Perspect. Biol. 2020, 12, a036392. [Google Scholar] [CrossRef]
- Du, T.; Gao, J.; Li, P.; Wang, Y.; Qi, Q.; Liu, X.; Li, J.; Wang, C.; Du, L. Pyroptosis, metabolism, and tumor immune microenvironment. Clin. Transl. Med. 2021, 11, e492. [Google Scholar] [CrossRef]
- Li, L.; Jiang, M.; Qi, L.; Wu, Y.; Song, D.; Gan, J.; Li, Y.; Bai, Y. Pyroptosis, a new bridge to tumor immunity. Cancer Sci. 2021, 112, 3979–3994. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, I.; Rayamajhi, M.; Miao, E.A. Programmed cell death as a defence against infection. Nat. Rev. Immunol. 2017, 17, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Son, D.J.; Lee, J.W.; Lee, Y.H.; Song, H.S.; Lee, C.K.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Feng, H.; Yang, Z.; Liang, J.; Jin, Z.; Chen, L.; Zhan, L.; Xuan, M.; Yan, J.; Kuang, J.; et al. The central role of a two-way positive feedback pathway in molecular targeted therapies-mediated pyroptosis in anaplastic thyroid cancer. Clin. Transl. Med. 2022, 12, e727. [Google Scholar] [CrossRef] [PubMed]
- Tomer, Y. Genetic susceptibility to autoimmune thyroid disease: Past, present, and future. Thyroid 2010, 20, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Wu, Y.; Hou, Y.; Liu, Y.; Liu, T.; Zhang, H.; Fan, C.; Guan, H.; Li, Y.; Shan, Z.; et al. Cytokine Secretion and Pyroptosis of Thyroid Follicular Cells Mediated by Enhanced NLRP3, NLRP1, NLRC4, and AIM2 Inflammasomes Are Associated With Autoimmune Thyroiditis. Front. Immunol. 2018, 9, 1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Liu, Y.; Lin, Y.; Liang, J. Radioactive Iodine-Refractory Differentiated Thyroid Cancer and Redifferentiation Therapy. Endocrinol. Metab. 2019, 34, 215–225. [Google Scholar] [CrossRef]
- Yu, S.; Medling, B.; Yagita, H.; Braley-Mullen, H. Characteristics of inflammatory cells in spontaneous autoimmune thyroiditis of NOD.H-2h4 mice. J. Autoimmun. 2001, 16, 37–46. [Google Scholar] [CrossRef]
- Rasooly, L.; Burek, C.L.; Rose, N.R. Iodine-induced autoimmune thyroiditis in NOD-H-2h4 mice. Clin. Immunol. Immunopathol. 1996, 81, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, E.; Ohye, H.; Amino, N.; Takata, K.; Arishima, T.; Kudo, T.; Ito, M.; Kubota, S.; Fukata, S.; Miyauchi, A. Clinical characteristics of 852 patients with subacute thyroiditis before treatment. Intern Med. 2008, 47, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Fatourechi, V.; Aniszewski, J.P.; Fatourechi, G.Z.; Atkinson, E.J.; Jacobsen, S.J. Clinical features and outcome of subacute thyroiditis in an incidence cohort: Olmsted County, Minnesota, study. J. Clin. Endocrinol. Metab. 2003, 88, 2100–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koirala, K.P.; Sharma, V. Treatment of Acute Painful Thyroiditis with Low Dose Prednisolone: A Study on Patients from Western Nepal. J. Clin. Diagn Res. 2015, 9, MC01–MC03. [Google Scholar] [CrossRef] [PubMed]
- Dorian, P.; Cass, D.; Schwartz, B.; Cooper, R.; Gelaznikas, R.; Barr, A. Amiodarone as compared with lidocaine for shock-resistant ventricular fibrillation. N. Engl. J. Med. 2002, 346, 884–890. [Google Scholar] [CrossRef]
- Yang, X.; Yang, L.X.; Wu, J.; Guo, M.L.; Zhang, Y.; Ma, S.G. Treatment of lidocaine on subacute thyroiditis via restraining inflammatory factor expression and inhibiting pyroptosis pathway. J. Cell Biochem. 2019, 120, 10964–10971. [Google Scholar] [CrossRef]
- Razi, M.; Chan, E.Y.; Tooze, S.A. Early endosomes and endosomal coatomer are required for autophagy. J. Cell Biol. 2009, 185, 305–321. [Google Scholar] [CrossRef] [Green Version]
- Beller, M.; Sztalryd, C.; Southall, N.; Bell, M.; Jackle, H.; Auld, D.S.; Oliver, B. COPI complex is a regulator of lipid homeostasis. PLoS Biol. 2008, 6, e292. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.D.; Agostinis, P. Cell death and immunity in cancer: From danger signals to mimicry of pathogen defense responses. Immunol. Rev. 2017, 280, 126–148. [Google Scholar] [CrossRef]
- Di Marco, T.; Bianchi, F.; Sfondrini, L.; Todoerti, K.; Bongarzone, I.; Maffioli, E.M.; Tedeschi, G.; Mazzoni, M.; Pagliardini, S.; Pellegrini, S.; et al. COPZ1 depletion in thyroid tumor cells triggers type I IFN response and immunogenic cell death. Cancer Lett. 2020, 476, 106–119. [Google Scholar] [CrossRef]
- Lucca, L.E.; Dominguez-Villar, M. Modulation of regulatory T cell function and stability by co-inhibitory receptors. Nat. Rev. Immunol. 2020, 20, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and TIGIT: Co-inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef] [PubMed]
- Matias-Guiu, X.; De Lellis, R. Medullary thyroid carcinoma: A 25-year perspective. Endocr. Pathol. 2014, 25, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, C.W.; Tan, L.C.; Wen, S.S.; Liao, T.; Zhang, Y.; Chen, T.Z.; Ma, B.; Yu, P.C.; Lu, Z.W.; et al. Immune Co-inhibitory Receptors PD-1, CTLA-4, TIM-3, LAG-3, and TIGIT in Medullary Thyroid Cancers: A Large Cohort Study. J. Clin. Endocrinol. Metab. 2021, 106, 120–132. [Google Scholar] [CrossRef]
Figure 1.
The mechanisms of autophagy in thyroid cancer.


{kind=link}
Table 1.
The characteristic summary of NAPCD in thyroid cancer.
| Drugs or Inducers | Disease | Key Factor | Pathway | In Vivo/In Vitro Experimental Validation | Reference | ||
|---|---|---|---|---|---|---|---|
| autophagy | genes | V600EBRAF | papillary thyroid cancer | WT1 | AKT/mTOR pathway, ERK/P65 pathway | In vivo and in virto | [57] |
| SQSTM1/p62 | papillary thyroid cancer | LC3-II | AMPK/AKT/mTOR pathway | In vivo and in virto | [63] | ||
| V600EBRAF | papillary thyroid cancer | LKB1-AMPK-ULK1 pathway | In vivo and in virto | [113] | |||
| SIRT6 | papillary thyroid cancer | GLUT1 | Warburg effect | In vivo and in virto | [63] | ||
| BIRC7 | papillary thyroid cancer | ATG5, BECN1 | EMT | In vivo and in virto | [70] | ||
| GANT61 | anaplastic thyroid cancer | LC3-II, p62, TAK1, JNK, AMPK | Shh pathway | In vitro | [67] | ||
| LDHA | papillary thyroid cancer | H3K27, FX11 | EMT | In vivo and in virto | [71] | ||
| FOXK2 | papillary thyroid cancer | ULK1, VPS34, FOXO3 | In vitro | [78] | |||
| RBM47 | papillary thyroid cancer | LC3-II, p62 | SNHG5/FOXO3 pathway | In vivo and in virto | [76] | ||
| TFE3 | papillary thyroid cancer | P62, LC3, CTSL, CTSB | In vitro | [82] | |||
| DAPK2 | papillary thyroid cancer | NF-Κb, I-κBα | In vivo and in virto | [83] | |||
| compounds | Capsaicin | anaplastic thyroid cancer | TRPV1 | In vitro | [46] | ||
| Prunella vulgaris L. | papillary thyroid cancer | LC3-II, beclin-1, p62 | AMPK/mTOR/ULK1 pathway | In vivo and in virto | [49] | ||
| Apatinib | papillary thyroid cancer | ATG5 | PI3K/Akt/mTOR pathway | In vivo and in virto | [101] | ||
| Apatinib | anaplastic thyroid cancer | SC79 | AKT/mTOR pathway | In vivo and in virto | [103] | ||
| Iodine | papillary thyroid cancer | V600EBRAF | BRAF/MEK/ERK (MAPK) pathway | In vitro | [108] | ||
| DINP | autoimmune thyroid disease | IL-17 | Akt/mTOR pathway | In vivo | [112] | ||
| Flavokawain B | thyroid cancer | p-AMPK, αThr172 | AMPK/mTOR pathway | In vitro | [113] | ||
| Curcumin | papillary thyroid cancer | LC3-II, beclin-1, p62 | AMPK/AKT/mTOR pathway | In vitro | [41] | ||
| LHPP | papillary thyroid cancer | AMPK/AKT/mTOR pathway | In vivo and in virto | [60] | |||
| ncRNA | lncRNA SLC26A4- AS1 | papillary thyroid cancer | ITPR1, ETS1 | In vitro | [92] | ||
| lncRNA DLX6- AS1 | papillary thyroid cancer | microRNA-193b-3p, HOXA1 | In vivo and in virto | [93] | |||
| lncRNA TNRC6C-AS1 | papillary thyroid cancer | STK4 | Hippo pathway | In vivo and in virto | [94] | ||
| lncRNA GAS8-AS1 | papillary thyroid cancer | ATF2 | miR-187-3p/ATG5, miR-1343-3p/ATG7 | In vivo and in virto | [95] | ||
| lncRNA RP11-476D10.1 | papillary thyroid cancer | microRNA-138-5p, LRRK2 | In vitro | [96] | |||
| lncRNA SNHG9 | papillary thyroid cancer | SNHG9 | YBOX3/P21 pathway | In vitro | [97] | ||
| MicroRNA-524-5p | papillary thyroid cancer | FOXE1, ITGA3 | In vitro | [99] | |||
| Ferroptosis | genes | APOE | thyroid cancer | In vivo | [126] | ||
| BCL3 | thyroid cancer | In vivo | [126] | ||||
| ALOX5AP | thyroid cancer | In vivo | [126] | ||||
| compounds | Vitamin C | anaplastic thyroid cancer | GPX4 | In vitro | [128] | ||
| ncRNA | Circ_0067934 | thyroid cancer | miR-545-3p/SLC7A11 | In vitro | [130] | ||
| Pyroptosis | genes | NLRP3 | autoimmune thyroiditis | In vivo | [137] | ||
| NLRP1 | autoimmune thyroiditis | In vivo | [137] | ||||
| NLRC4 | autoimmune thyroiditis | In vivo | [137] | ||||
| AIM2 | autoimmune thyroiditis | In vivo | [137] | ||||
| compounds | Iodine | hashimoto’s thyroiditis | ROS | In vivo | [141] | ||
| lidocaine | subacute thyroiditis | In vitro | [146] | ||||
| Melittin | anaplastic thyroid cancer | apatinib | In vitro | [135] | |||
| Immunogenic cell death | genes | COPZ1 | papillary thyroid cancer | type I IFN | In vitro | [151] | |
| TIM-3 | medullary thyroid carcinoma | In vivo | [155] | ||||
| PD-1 | medullary thyroid carcinoma | In vivo | [155] | ||||
| CTLA-4 | medullary thyroid carcinoma | In vivo | [155] | ||||
| LAG-3 | medullary thyroid carcinoma | In vivo | [155] | ||||
| TIGIT | medullary thyroid carcinoma | In vivo | [155] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ji, F.; Qiu, X. Non-Apoptotic Programmed Cell Death in Thyroid Diseases. Pharmaceuticals 2022, 15, 1565. https://doi.org/10.3390/ph15121565
AMA Style
Ji F, Qiu X. Non-Apoptotic Programmed Cell Death in Thyroid Diseases. Pharmaceuticals. 2022; 15(12):1565. https://doi.org/10.3390/ph15121565
Chicago/Turabian StyleJi, Feihong, and Xinguang Qiu. 2022. "Non-Apoptotic Programmed Cell Death in Thyroid Diseases" Pharmaceuticals 15, no. 12: 1565. https://doi.org/10.3390/ph15121565
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.