
Three-Dimensional Microtumor Formation of Infantile Hemangioma-Derived Endothelial Cells for Mechanistic Exploration and Drug Screening


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
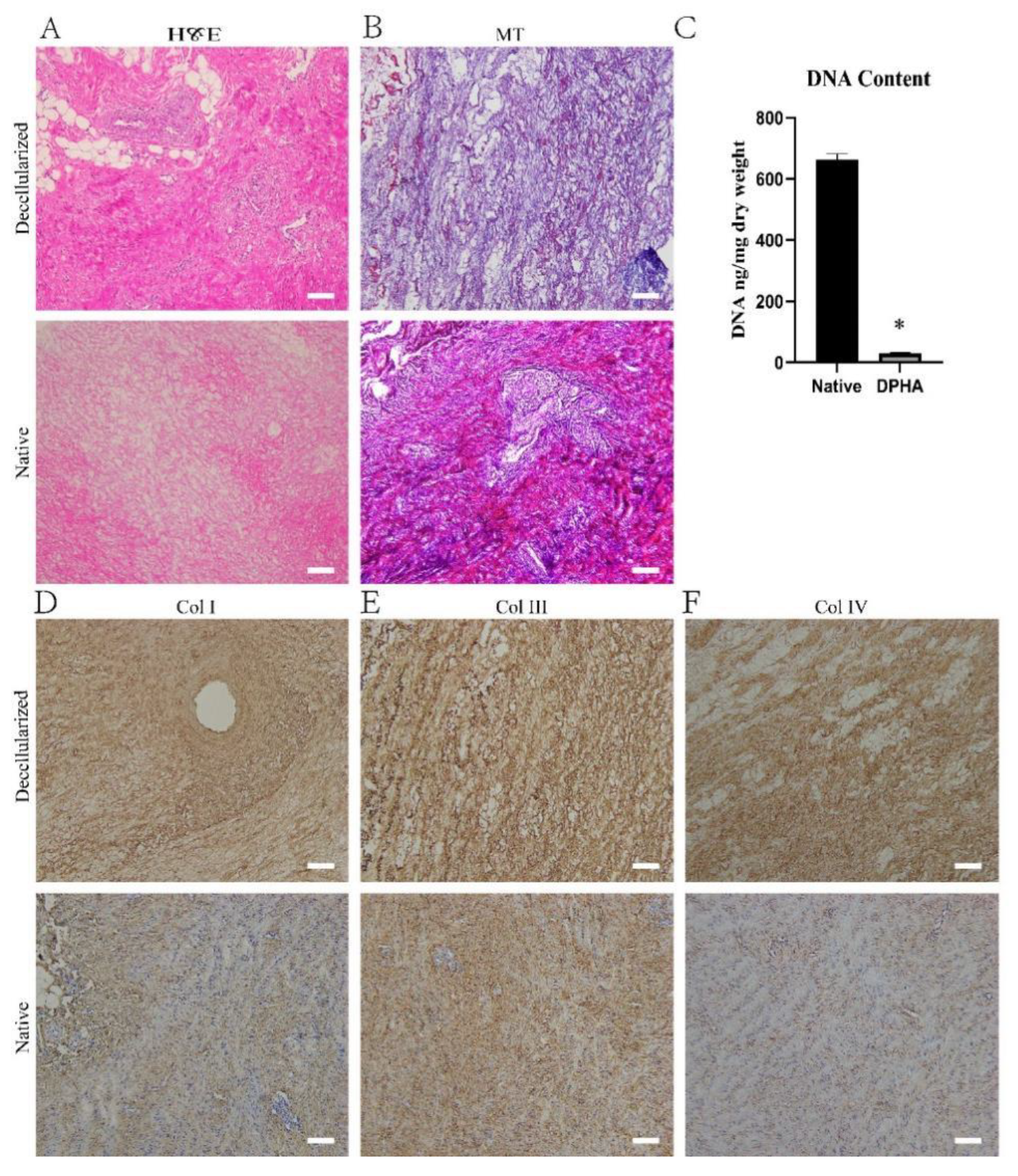
2.1. Characterization of DPHASs
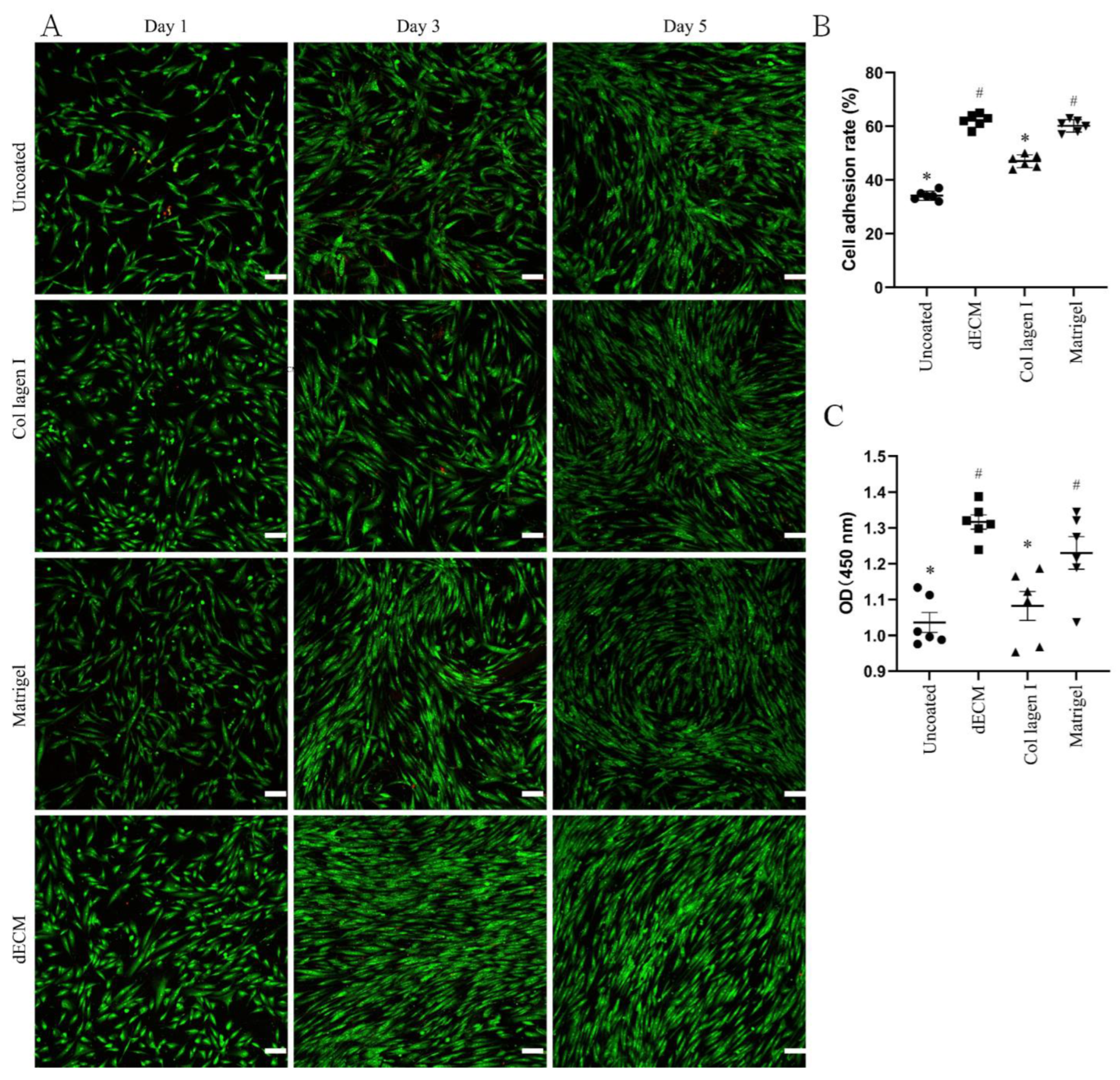
2.2. Evaluation of dECM-Coated Plate Characteristics
2.3. Specific Features of the Micropattern Array
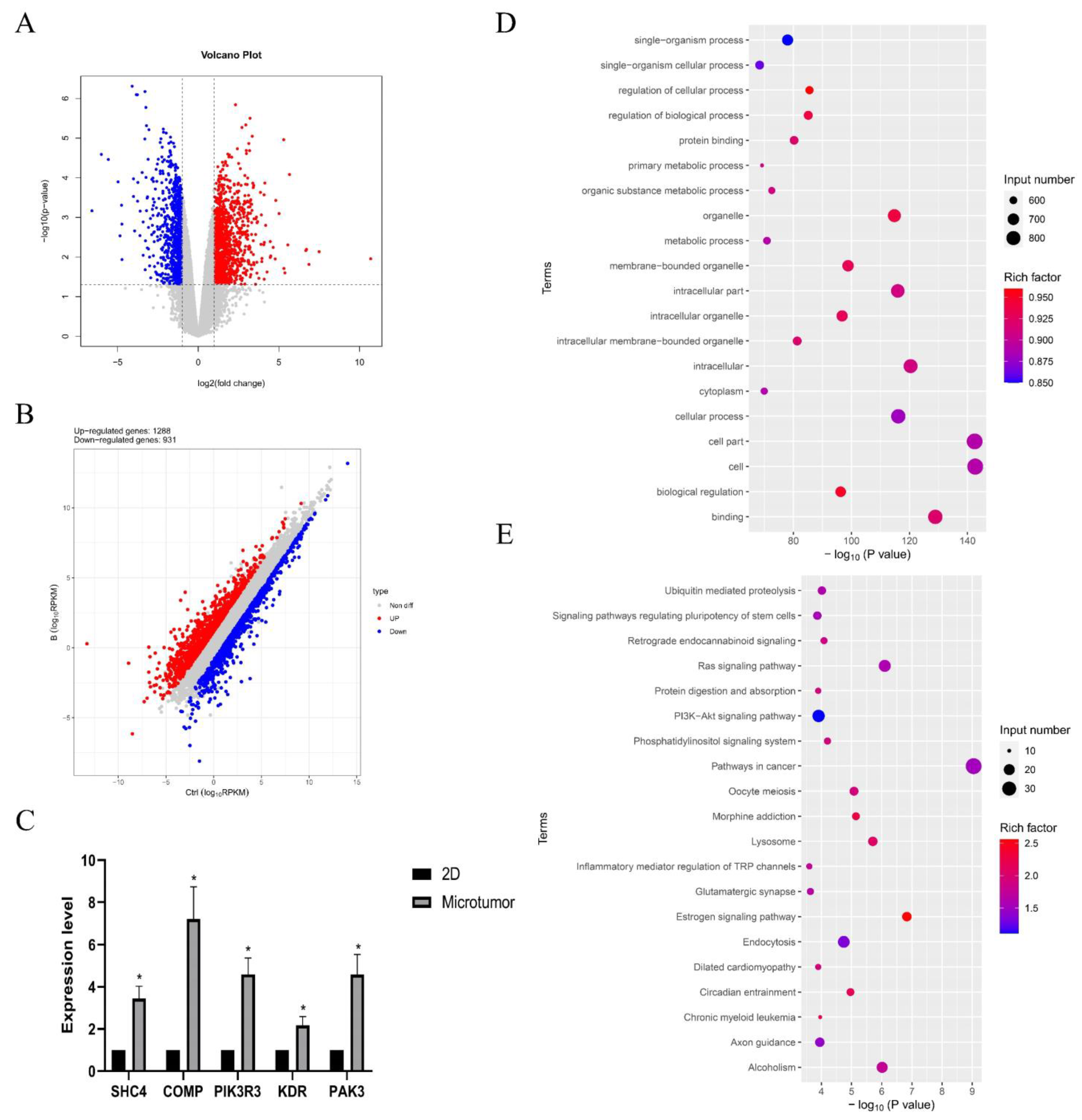
2.4. Results of the RNA Sequencing
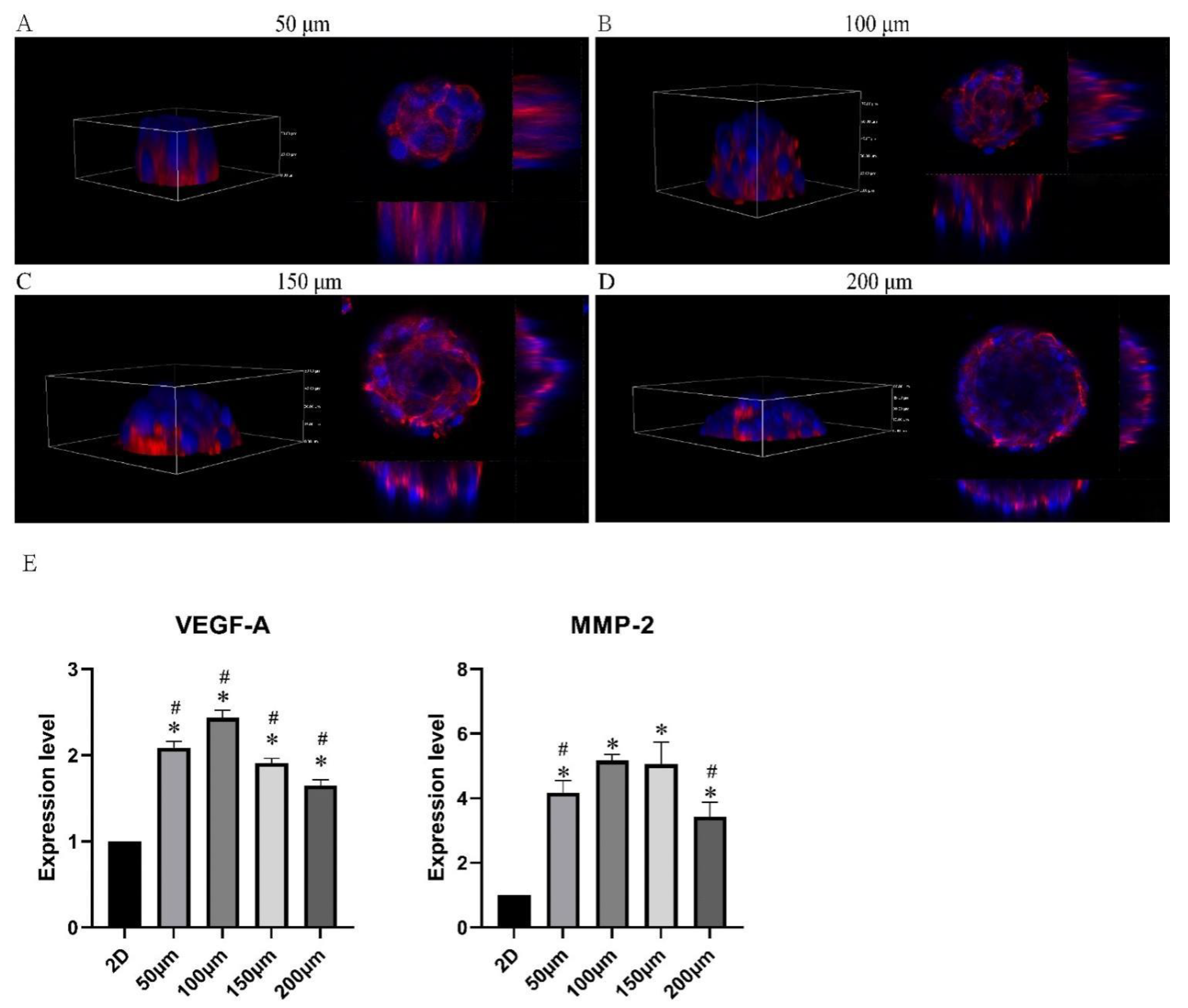
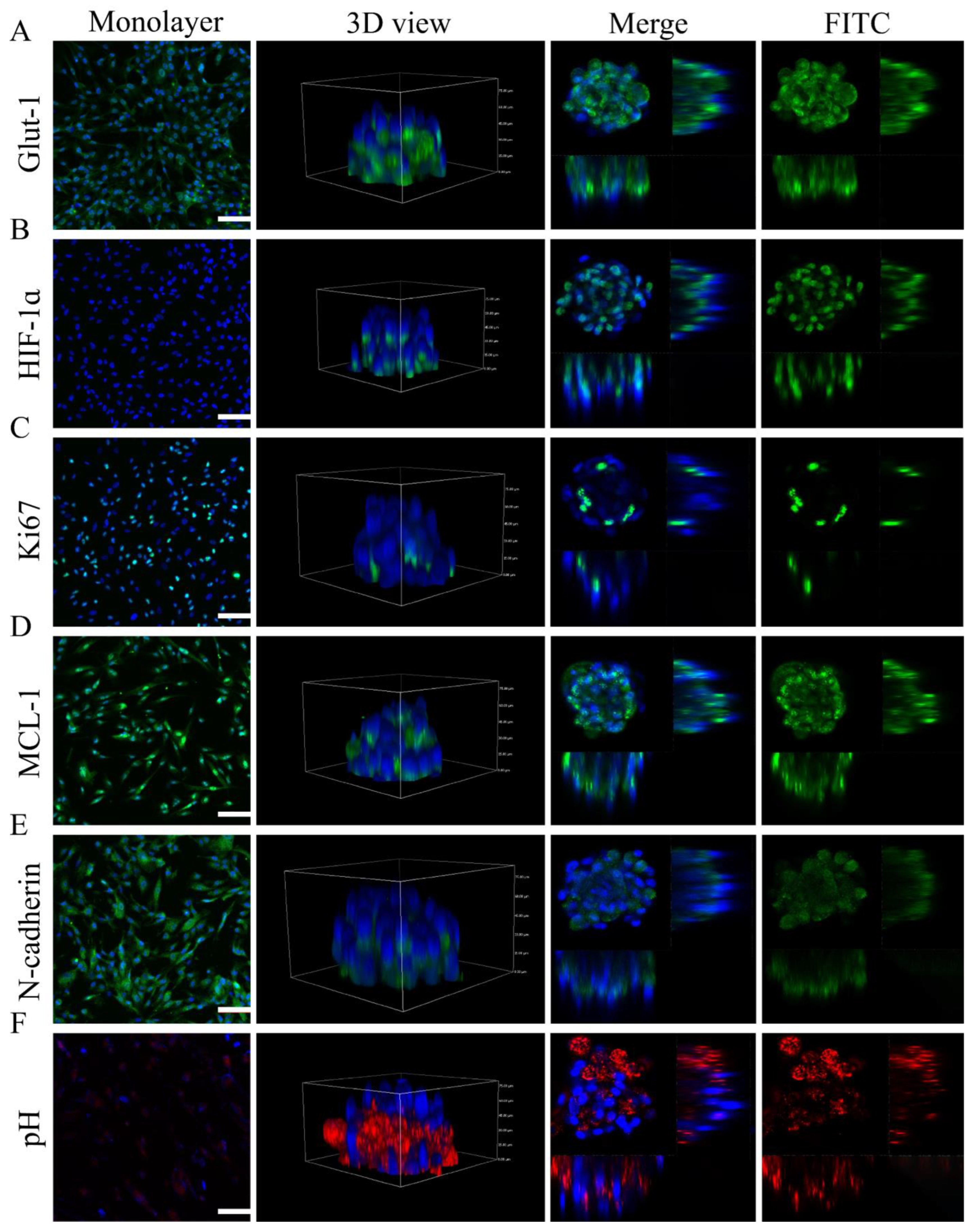
2.5. Microtumor Characteristics
2.6. Drug Screening
3. Materials and Methods
3.1. Decellularization of Porcine Heart Aortic Tissue
3.2. Preparation of the Vascular-Specific dECM Hydrogel
3.3. Cell Culture
3.4. Evaluation of Plate Coating and Features
3.5. Printing of Micropattern Arrays
3.6. Culturing of CD31+ HemEC Microtumors
3.7. RNA Sequencing
3.8. Quantitative Real-Time Polymerase Chain Reaction (qRT–PCR)
3.9. Features of CD31+ HemEC Microtumors
3.10. Evaluation of Chemotherapeutic Medication Cytotoxicity
3.11. Data Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moisan, F.; Oucherif, S.; Kaulanjan-Checkmodine, P.; Prey, S.; Rousseau, B.; Bonneu, M.; Claverol, S.; Gontier, E.; Lacomme, S.; Dousset, L.; et al. Critical role of Aquaporin-1 and telocytes in infantile hemangioma response to propranolol beta blockade. Proc. Natl. Acad. Sci. USA 2021, 118, e2018690118. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.R.; Schoch, J.J.; Lohse, C.M.; Hand, J.L.; Davis, D.M.; Tollefson, M.M. Increasing incidence of infantile hemangiomas (IH) over the past 35 years: Correlation with decreasing gestational age at birth and birth weight. J. Am. Acad. Dermatol. 2016, 74, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, C.; Solman, L.; Flohr, C. A national audit of oral propranolol for the treatment of infantile haemangiomas. Br. J. Dermatol. 2021, 186, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Itinteang, T.; Tan, S.T.; Brasch, H.; Day, D.J. Primitive mesodermal cells with a neural crest stem cell phenotype predominate proliferating infantile haemangioma. J. Clin. Pathol. 2010, 63, 771–776. [Google Scholar] [CrossRef] [Green Version]
- de Jong, S.; Itinteang, T.; Withers, A.H.; Davis, P.F.; Tan, S.T. Does hypoxia play a role in infantile hemangioma? Arch. Dermatol. Res. 2016, 308, 219–227. [Google Scholar] [CrossRef]
- Xia, H.F.; Zhu, J.Y.; Wang, J.N.; Ren, J.G.; Cai, Y.; Wang, F.Q.; Zhang, W.; Chen, G.; Zhao, Y.-F.; Zhao, J.-H. Association of ATF4 Expression With Tissue Hypoxia and M2 Macrophage Infiltration in Infantile Hemangioma. J. Histochem. Cytochem. 2017, 65, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Itinteang, T.; Withers, A.H.; Davis, P.F.; Tan, S.T. Biology of infantile hemangioma. Front. Surg. 2014, 1, 38. [Google Scholar] [CrossRef] [Green Version]
- Itinteang, T.; Marsh, R.; Davis, P.F.; Tan, S.T. Angiotensin II causes cellular proliferation in infantile haemangioma via angiotensin II receptor 2 activation. J. Clin. Pathol. 2015, 68, 346–350. [Google Scholar] [CrossRef]
- Itinteang, T.; Chudakova, D.A.; Dunne, J.C.; Davis, P.F.; Tan, S.T. Expression of Cathepsins B, D, and G in Infantile Hemangioma. Front. Surg. 2015, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Leaute-Labreze, C.; de la Roque, E.D.; Hubiche, T.; Boralevi, F.; Thambo, J.B.; Taieb, A. Propranolol for severe hemangiomas of infancy. N. Engl. J. Med. 2008, 358, 2649–2651. [Google Scholar] [CrossRef]
- Sebaratnam, D.F.; Bandera, A.L.R.; Wong, L.F.; Wargon, O. Infantile hemangioma. Part 2: Management. J. Am. Acad. Dermatol. 2021, 85, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.A.; Melero-Martin, J.M.; Wu, X.; Paruchuri, S.; Boscolo, E.; Mulliken, J.B.; Bischoff, J. Endothelial progenitor cells from infantile hemangioma and umbilical cord blood display unique cellular responses to endostatin. Blood 2006, 108, 915–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Z.A.; Boscolo, E.; Picard, A.; Psutka, S.; Melero-Martin, J.M.; Bartch, T.C.; Mulliken, J.B.; Bischoff, J. Multipotential stem cells recapitulate human infantile hemangioma in immunodeficient mice. J. Clin. Investig. 2008, 118, 2592–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguli, A.; Mostafa, A.; Saavedra, C.; Kim, Y.; Le, P.; Faramarzi, V.; Feathers, R.W.; Berger, J.; Ramos-Cruz, K.P.; Adeniba, O.; et al. Three-dimensional microscale hanging drop arrays with geometric control for drug screening and live tissue imaging. Sci. Adv. 2021, 7, eabc1323. [Google Scholar] [CrossRef]
- Kim, C.H.; Suhito, I.R.; Angeline, N.; Han, Y.; Son, H.; Luo, Z.; Kim, T.-H. Vertically Coated Graphene Oxide Micro-Well Arrays for Highly Efficient Cancer Spheroid Formation and Drug Screening. Adv. Healthc. Mater. 2020, 9, e1901751. [Google Scholar] [CrossRef]
- Hu, S.; Chen, T.H.; Zhao, Y.; Wang, Z.; Lam, R.H.W. Protein-Substrate Adhesion in Microcontact Printing Regulates Cell Behavior. Langmuir 2018, 34, 1750–1759. [Google Scholar] [CrossRef]
- Zhang, Z.; Chen, Y.C.; Urs, S.; Chen, L.; Simeone, D.M.; Yoon, E. Scalable Multiplexed Drug-Combination Screening Platforms Using 3D Microtumor Model for Precision Medicine. Small 2018, 14, e1703617. [Google Scholar] [CrossRef]
- Wang, Z.; Lang, B.; Qu, Y.; Li, L.; Song, Z.; Wang, Z. Single-cell patterning technology for biological applications. Biomicrofluidics 2019, 13, 061502. [Google Scholar] [CrossRef]
- Kabirian, F.; Mozafari, M. Decellularized ECM-derived bioinks: Prospects for the future. Methods 2020, 171, 108–118. [Google Scholar] [CrossRef]
- Multhaupt, H.A.; Leitinger, B.; Gullberg, D.; Couchman, J.R. Extracellular matrix component signaling in cancer. Adv. Drug Deliv. Rev. 2016, 97, 28–40. [Google Scholar] [CrossRef]
- Mulliken, J.B.; Zetter, B.R.; Folkman, J. In vitro characteristics of endothelium from hemangiomas and vascular malformations. Surgery 1982, 92, 348–353. [Google Scholar] [PubMed]
- Bischoff, J. Progenitor cells in infantile hemangioma. J. Craniofac. Surg. 2009, 20 (Suppl. S1), 695–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, H.M.; Zheng, J.W.; Wang, Y.A.; Yang, X.J.; Zhou, Q.; Qin, Z.P.; Li, K.-L. CD133 selected stem cells from proliferating infantile hemangioma and establishment of an in vivo mice model of hemangioma. Chin. Med. J. 2013, 126, 88–94. [Google Scholar] [PubMed]
- O’Connell, L.; Winter, D.C. Organoids: Past Learning and Future Directions. Stem Cells Dev. 2020, 29, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Boscolo, E.; Durham, J.T.; Mulliken, J.B.; Herman, I.M.; Bischoff, J. Propranolol targets the contractility of infantile haemangioma-derived pericytes. Br. J. Dermatol. 2014, 171, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Li, K.; Xiao, X.; Zheng, S.; Xu, T.; Chen, S. Effects of propranolol on the proliferation and apoptosis of hemangioma-derived endothelial cells. J. Pediatr. Surg. 2012, 47, 2216–2223. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, X.; Chen, L.; Chen, S.; Ji, Y. Microarray expression profile of mRNAs and long noncoding RNAs and the potential role of PFK-1 in infantile hemangioma. Cell Div. 2021, 16, 1. [Google Scholar] [CrossRef]
- Zhu, X.; Li, Y.; Yang, Y.; He, Y.; Gao, M.; Peng, W.; Wu, Q.; Zhang, G.; Zhou, Y.; Chen, F.; et al. Ordered micropattern arrays fabricated by lung-derived dECM hydrogels for chemotherapeutic drug screening. Mater. Today Bio. 2022, 15, 100274. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Harbi, S.; Wang, R.; Gregory, M.; Hanson, N.; Kobylarz, K.; Ryan, K.; Deng, Y.; Lopez, P.; Chiriboga, L.; Mignatti, P. Infantile Hemangioma Originates From A Dysregulated But Not Fully Transformed Multipotent Stem Cell. Sci. Rep. 2016, 6, 35811. [Google Scholar] [CrossRef]
- Xu, D.; Teresa, M.O.; Shartava, A.; Fowles, T.C.; Yang, J.; Fink, L.M.; Ward, D.C.; Mihm, M.C.; Waner, M.; Ma, Y. Isolation, characterization, and in vitro propagation of infantile hemangioma stem cells and an in vivo mouse model. J. Hematol. Oncol. 2011, 4, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heide, C.D.; Dalm, S.U. Radionuclide imaging and therapy directed towards the tumor microenvironment: A multi-cancer approach for personalized medicine. Eur. J. Nucl. Med. Mol. Imaging 2022, 49, 4616–4641. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Choi, N.; Kim, K.; Koo, H.J.; Choi, J.; Kim, H.N. Chemoresistance of Cancer Cells: Requirements of Tumor Microenvironment-mimicking In Vitro Models in Anti-Cancer Drug Development. Theranostics 2018, 8, 5259–5275. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Xu, J.; Wang, W.; Liang, C.; Hua, J.; Liu, J.; Zhang, B.; Meng, Q.; Yu, X.; Shi, S. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: New findings and future perspectives. Mol. Cancer 2021, 20, 131. [Google Scholar] [CrossRef]
- Hinshaw, D.C.; Shevde, L.A. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res. 2019, 79, 4557–4566. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Walker, C.; Mojares, E.; del Rio Hernandez, A. Role of Extracellular Matrix in Development and Cancer Progression. Int. J. Mol. Sci. 2018, 19, 3028. [Google Scholar] [CrossRef] [Green Version]
- North, P.E.; Waner, M.; Mizeracki, A.; Mrak, R.E.; Nicholas, R.; Kincannon, J.; Suen, J.Y.; Mihm, M.C. A unique microvascular phenotype shared by juvenile hemangiomas and human placenta. Arch. Dermatol. 2001, 137, 559–570. [Google Scholar]
- Fu, C.; Yang, K.; Zou, Y.; Huo, R. Identification of Key microRNAs and Genes in Infantile Hemangiomas. Front. Genet. 2022, 13, 766561. [Google Scholar] [CrossRef]
- Hu, W.; Liu, Z.; Salato, V.; North, P.E.; Bischoff, J.; Kumar, S.N.; Fang, Z.; Rajan, S.; Hussain, M.M.; Miao, Q.R. NOGOB receptor-mediated RAS signaling pathway is a target for suppressing proliferating hemangioma. JCI Insight 2021, 6, e142299. [Google Scholar] [CrossRef]
- Fagiani, E.; Giardina, G.; Luzi, L.; Cesaroni, M.; Quarto, M.; Capra, M.; Germano, G.; Bono, M.; Capillo, M.; Pelicci, P.; et al. RaLP, A new member of the Src homology and collagen family, regulates cell migration and tumor growth of metastatic melanomas. Cancer Res. 2007, 67, 3064–3073. [Google Scholar] [CrossRef] [PubMed]
- Turco, M.Y.; Furia, L.; Dietze, A.; Diaz, L.F.; Ronzoni, S.; Sciullo, A.; Simeone, A.; Constam, D.; Faretta, M.; Lanfrancone, L. Cellular heterogeneity during embryonic stem cell differentiation to epiblast stem cells is revealed by the ShcD/RaLP adaptor protein. Stem Cells 2012, 30, 2423–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Yi, T.; Huang, D.; Chen, X.; Li, X.; Wan, Q.; Huang, H.; Huang, H.; Wei, H.; Song, Y.; et al. circ_PTN contributes to -cisplatin resistance in glioblastoma via PI3K/AKT signaling through the miR-542-3p/PIK3R3 pathway. Mol. Ther. Nucleic Acids 2021, 26, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Sun, X.; Luo, X.; Wang, J.; Hu, J.; Feng, Y. PIK3R3 inhibits cell senescence through p53/p21 signaling. Cell Death Dis. 2020, 11, 798. [Google Scholar] [CrossRef] [PubMed]
- Jinnin, M.; Medici, D.; Park, L.; Limaye, N.; Liu, Y.; Boscolo, E.; Bischoff, J.; Vikkula, M.; Boye, E.; Olsen, B.R. Suppressed NFAT-dependent VEGFR1 expression and constitutive VEGFR2 signaling in infantile hemangioma. Nat. Med. 2008, 14, 1236–1246. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Flint, A.F.; Mulliken, J.B.; Wu, J.K.; Bischoff, J. Endothelial progenitor cells in infantile hemangioma. Blood 2004, 103, 1373–1375. [Google Scholar] [CrossRef] [Green Version]
- Piccand, J.; Meunier, A.; Merle, C.; Jia, Z.; Barnier, J.V.; Gradwohl, G. Pak3 promotes cell cycle exit and differentiation of beta-cells in the embryonic pancreas and is necessary to maintain glucose homeostasis in adult mice. Diabetes 2014, 63, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.; Hou, H.; Liu, T.; Su, S.; Xi, X.; Liao, Y.; Xie, R.; Jin, G.; Liu, X.; Zhu, L.; et al. Cartilage Oligomeric Matrix Protein promotes epithelial-mesenchymal transition by interacting with Transgelin in Colorectal Cancer. Theranostics 2020, 10, 8790–8806. [Google Scholar] [CrossRef]
- Kleinman, M.E.; Greives, M.R.; Churgin, S.S.; Blechman, K.M.; Chang, E.I.; Ceradini, D.J.; Tepper, O.M.; Gurtner, G.C. Hypoxia-induced mediators of stem/progenitor cell trafficking are increased in children with hemangioma. Arter. Thromb. Vasc. Biol. 2007, 27, 2664–2670. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Li, Y.; Yang, K.; Chen, S.; Ji, Y. Infantile hepatic hemangiomas: Looking backwards and forwards. Precis. Clin. Med. 2022, 5, pbac006. [Google Scholar] [CrossRef]
- Rohwer, N.; Cramer, T. Hypoxia-mediated drug resistance: Novel insights on the functional interaction of HIFs and cell death pathways. Drug. Resist. Updat. 2011, 14, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Greenberger, S.; Bischoff, J. Infantile hemangioma-mechanism(s) of drug action on a vascular tumor. Cold Spring Harb. Perspect. Med. 2011, 1, a006460. [Google Scholar] [CrossRef] [Green Version]
- Potente, M.; Carmeliet, P. The Link Between Angiogenesis and Endothelial Metabolism. Annu. Rev. Physiol. 2017, 79, 43–66. [Google Scholar] [CrossRef] [PubMed]
- De Bock, K.; Georgiadou, M.; Carmeliet, P. Role of endothelial cell metabolism in vessel sprouting. Cell. Metab. 2013, 18, 634–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wu, D.; Dong, Z.; Chen, A.; Liu, S. The expression and role of glycolysis-associated molecules in infantile hemangioma. Life Sci. 2020, 259, 118215. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Gou, Q.; Jin, J.; Shi, J.; Liu, Q.; Hou, Y. Metformin inhibits PPARdelta agonist-mediated tumor growth by reducing Glut1 and SLC1A5 expressions of cancer cells. Eur. J. Pharmacol. 2019, 857, 172425. [Google Scholar] [CrossRef]
- Popovic, K.J.; Popovic, D.J.; Miljkovic, D.; Popovic, J.K.; Lalosevic, D.; Posa, M.; Čapo, I. Disulfiram and metformin combination anticancer effect reversible partly by antioxidant nitroglycerin and completely by NF-kappaB activator mebendazole in hamster fibrosarcoma. Biomed. Pharmacother. 2021, 143, 112168. [Google Scholar] [CrossRef]
- Linehan, W.M.; Rouault, T.A. Molecular pathways: Fumarate hydratase-deficient kidney cancer—targeting the Warburg effect in cancer. Clin. Cancer Res. 2013, 19, 3345–3352. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Sawada, K.; Okumura, T. A case of liver hemangioma with markedly reduced tumor size after metformin treatment: A case report. Clin. J. Gastroenterol. 2017, 10, 63–67. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhu, X.; Kong, M.; Chen, S.; Bao, J.; Ji, Y. Three-Dimensional Microtumor Formation of Infantile Hemangioma-Derived Endothelial Cells for Mechanistic Exploration and Drug Screening. Pharmaceuticals 2022, 15, 1393. https://doi.org/10.3390/ph15111393
Li Y, Zhu X, Kong M, Chen S, Bao J, Ji Y. Three-Dimensional Microtumor Formation of Infantile Hemangioma-Derived Endothelial Cells for Mechanistic Exploration and Drug Screening. Pharmaceuticals. 2022; 15(11):1393. https://doi.org/10.3390/ph15111393
Chicago/Turabian StyleLi, Yanan, Xinglong Zhu, Meng Kong, Siyuan Chen, Ji Bao, and Yi Ji. 2022. "Three-Dimensional Microtumor Formation of Infantile Hemangioma-Derived Endothelial Cells for Mechanistic Exploration and Drug Screening" Pharmaceuticals 15, no. 11: 1393. https://doi.org/10.3390/ph15111393