
Human Umbilical Cord MSC Delivered-Soluble TRAIL Inhibits the Proliferation and Promotes Apoptosis of B-ALL Cell In Vitro and In Vivo

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. rhTRAIL Inhibits the Proliferation of B-ALL Cells
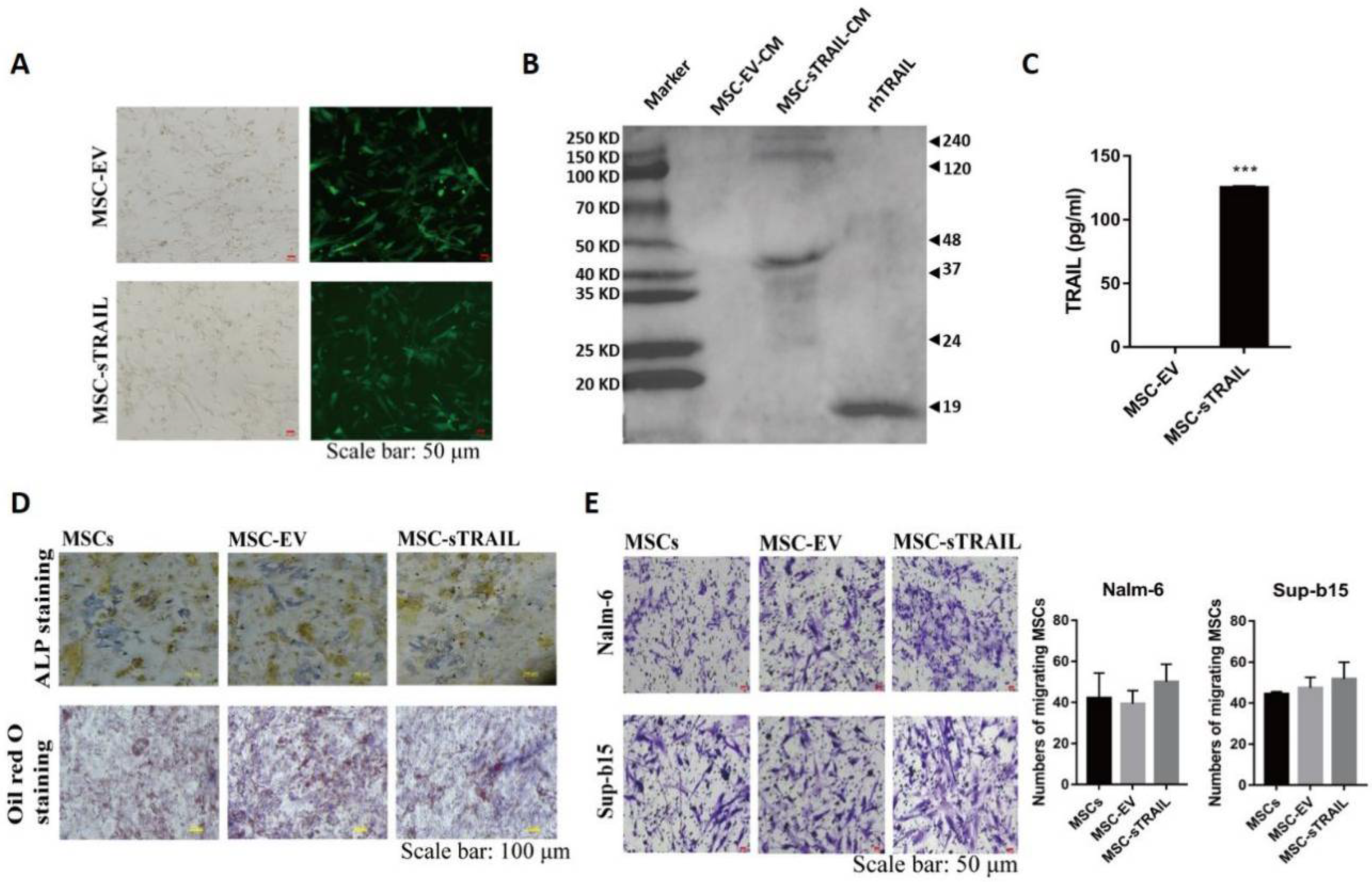
2.2. Gene Modified UC-MSCs Can Secrete Soluble TRAIL (sTRAIL) without Apparent Interference on Their Biological Properties
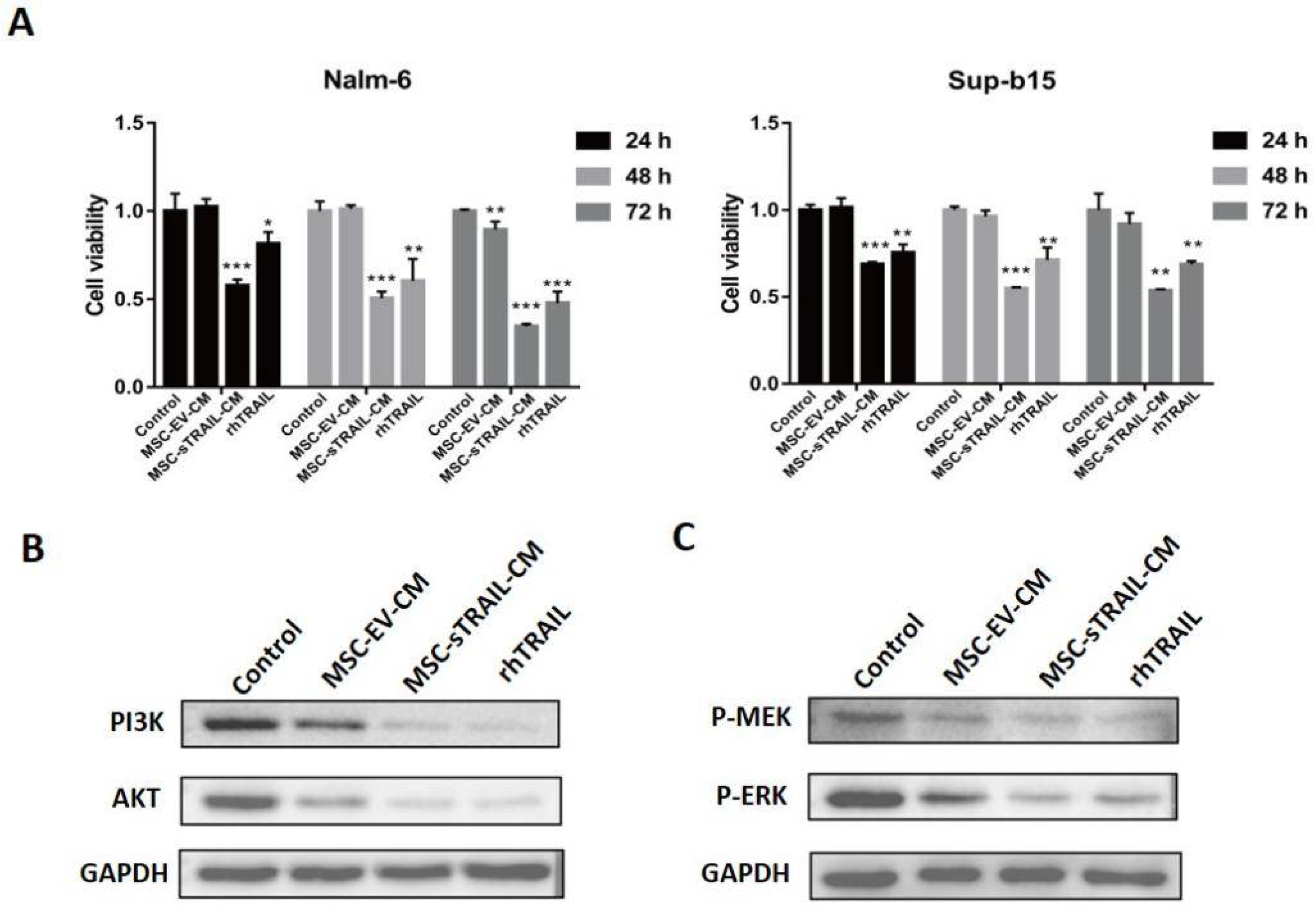
2.3. MSC-sTRAIL Inhibits the Proliferation of B-ALL Cells In Vitro
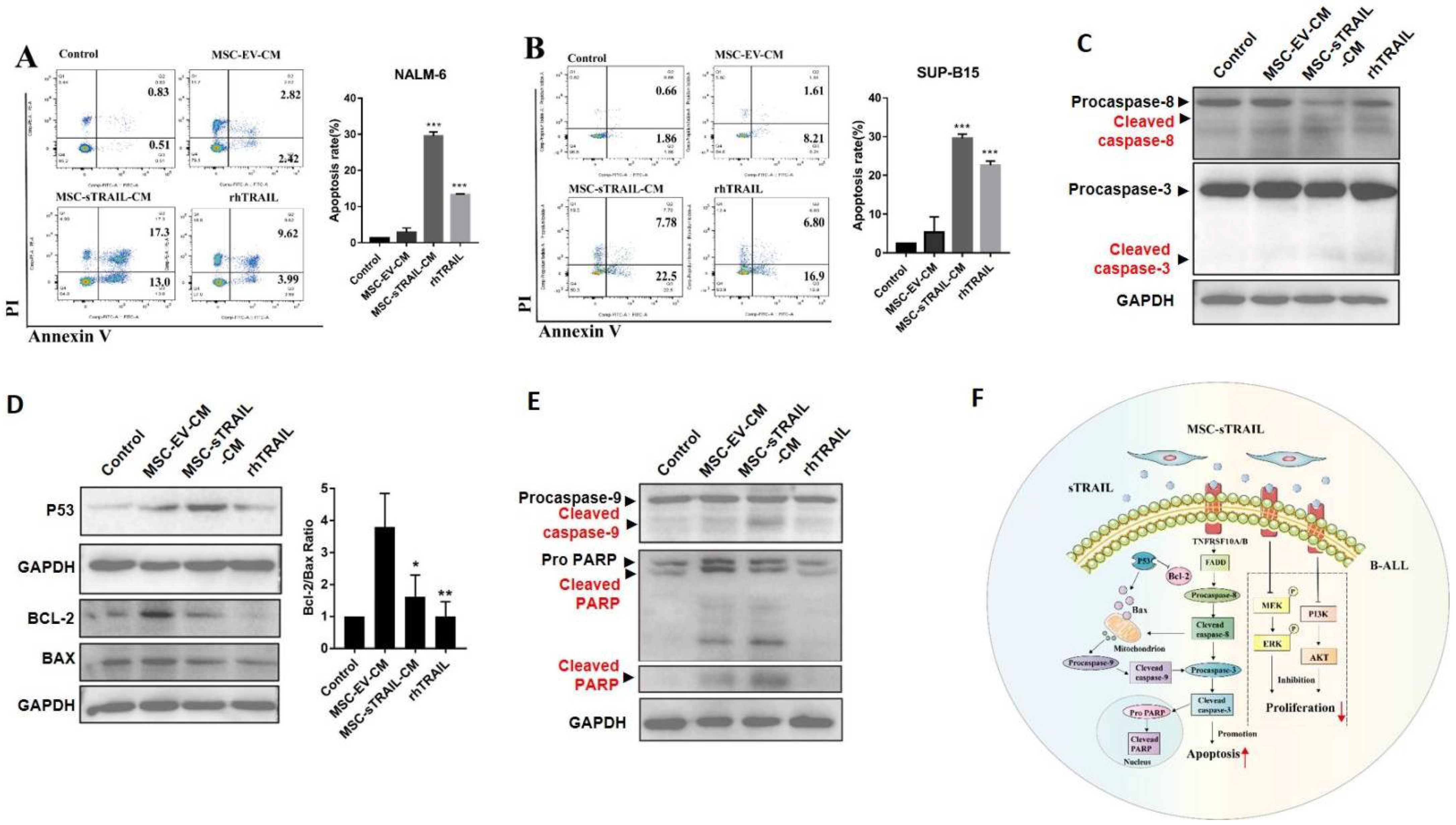
2.4. MSC-sTRAIL Induces Apoptosis of B-ALL Cells In Vitro
2.5. MSC-sTRAIL Showed Little Effects on the Expression Levels of TRAIL Receptors of B-ALL Cells
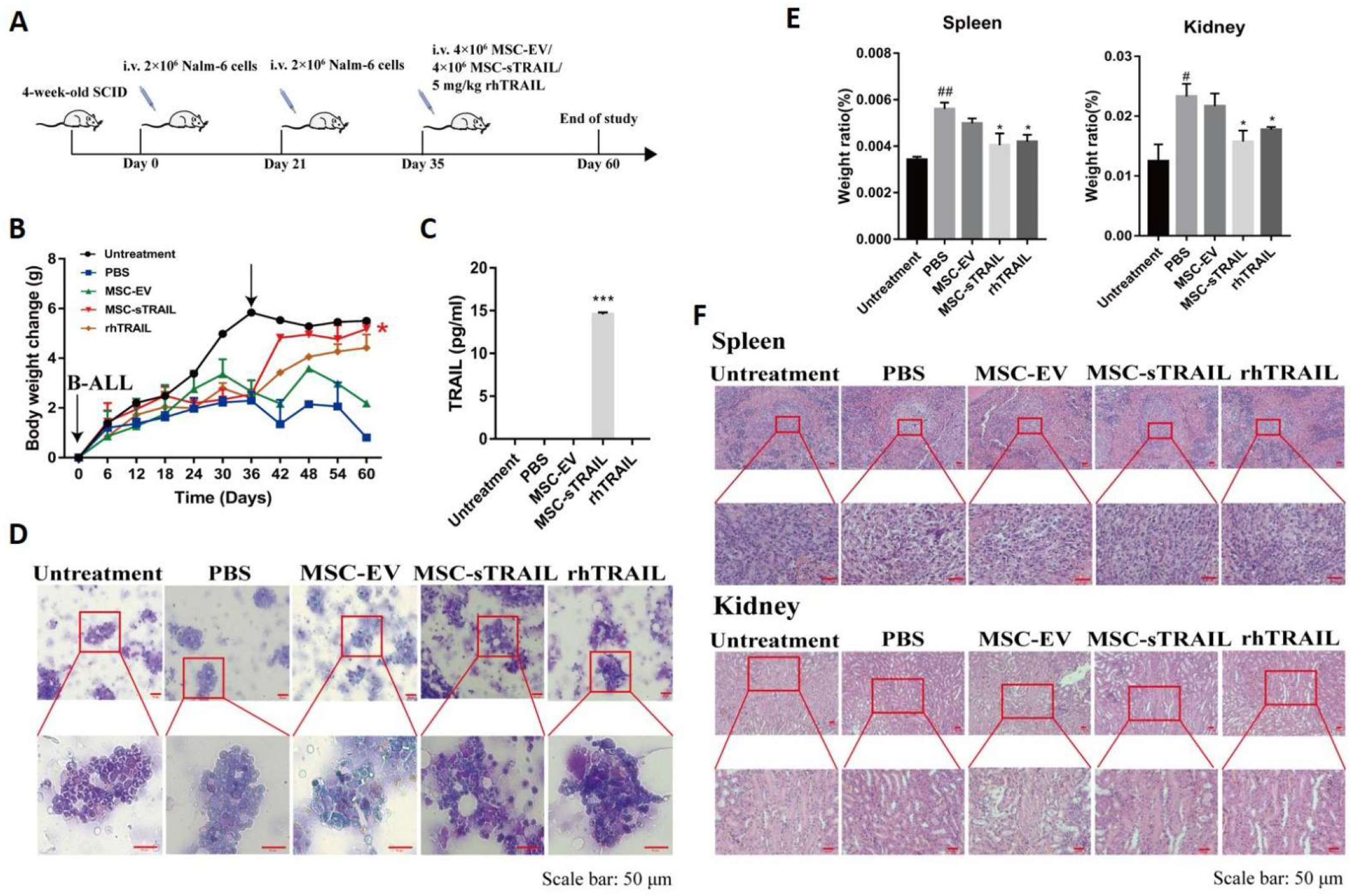
2.6. MSC-sTRAIL Delivered in Mice Efficiently Control the B-ALL Cell Levels in Bone Marrow
2.7. MSC-sTRAIL-Delivery Alleviates B-ALL Cell-Induced Tissue Injuries in Mice
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animal
4.3. Cell Viability Assay
4.4. Lentivial Plasmid Construction and Transduction
4.5. Conditioned Medium Preparation
4.6. Western Blotting
4.7. Measurement of TRAIL Levels by Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Hematoxylin and Eosin (H&E) Staining
4.9. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.W.; Cho, B. Prognostic factors and treatment of pediatric acute lymphoblastic leukemia. Korean J. Pediatr. 2017, 60, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, S.; Zhang, J.; Liu, C.; Li, X.; Guo, W.; Duan, Y.; Chen, X.; Zong, S.; Zheng, J.; et al. Elucidating minimal residual disease of paediatric B-cell acute lymphoblastic leukaemia by single-cell analysis. Nat. Cell Biol. 2022, 24, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Terwilliger, T.; Abdul-Hay, M. Acute lymphoblastic leukemia: A comprehensive review and 2017 update. Blood Cancer J. 2017, 7, e577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt, K.D.; Bram, R.J. Immunotherapy in pediatric B-cell acute lymphoblastic leukemia. Hum. Immunol. 2019, 80, 400–408. [Google Scholar] [CrossRef]
- Xu, X.; Sun, Q.; Liang, X.; Chen, Z.; Zhang, X.; Zhou, X.; Li, M.; Tu, H.; Liu, Y.; Tu, S.; et al. Mechanisms of Relapse After CD19 CAR T-Cell Therapy for Acute Lymphoblastic Leukemia and Its Prevention and Treatment Strategies. Front. Immunol. 2019, 10, 2664. [Google Scholar] [CrossRef] [Green Version]
- Riegler, L.L.; Jones, G.P.; Lee, D.W. Current approaches in the grading and management of cytokine release syndrome after chimeric antigen receptor T-cell therapy. Ther. Clin. Risk Manag. 2019, 15, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Newsom-Davis, T.; Prieske, S.; Walczak, H. Is TRAIL the holy grail of cancer therapy? Apoptosis 2009, 14, 607–623. [Google Scholar] [CrossRef]
- Kelley, S.K.; Ashkenazi, A. Targeting death receptors in cancer with Apo2L/TRAIL. Curr. Opin. Pharmacol. 2004, 4, 333–339. [Google Scholar] [CrossRef]
- Tazzari, P.L.; Tabellini, G.; Ricci, F.; Papa, V.; Bortul, R.; Chiarini, F.; Evangelisti, C.; Martinelli, G.; Bontadini, A.; Cocco, L.; et al. Synergistic proapoptotic activity of recombinant TRAIL plus the Akt inhibitor Perifosine in acute myelogenous leukemia cells. Cancer Res. 2008, 68, 9394–9403. [Google Scholar] [CrossRef] [Green Version]
- Asati, V.; Mahapatra, D.K.; Bharti, S.K. PI3K/Akt/mTOR and Ras/Raf/MEK/ERK signaling pathways inhibitors as anticancer agents: Structural and pharmacological perspectives. Eur. J. Med. Chem. 2016, 109, 314–341. [Google Scholar] [CrossRef]
- Jin, Y.; Qiu, S.; Shao, N.; Zheng, J. Fucoxanthin and Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL) Synergistically Promotes Apoptosis of Human Cervical Cancer Cells by Targeting PI3K/Akt/NF-κB Signaling Pathway. Med. Sci. Monit. 2018, 24, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Plastaras, J.P.; Dorsey, J.F.; Carroll, K.; Kim, S.-H.; Birnbaum, M.J.; El-Deiry, W.S. Role of PI3K/Akt signaling in TRAIL- and radiation-induced gastrointestinal apoptosis. Cancer Biol. Ther. 2008, 7, 2047–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso Alves, L.; Corazza, N.; Micheau, O.; Krebs, P. The multifaceted role of TRAIL signaling in cancer and immunity. FEBS J. 2021, 288, 5530–5554. [Google Scholar] [CrossRef]
- Yuan, X.; Gajan, A.; Chu, Q.; Xiong, H.; Wu, K.; Wu, G.S. Developing TRAIL/TRAIL death receptor-based cancer therapies. Cancer Metastasis Rev. 2018, 37, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Chaudhry, G.-E. Understanding Apoptosis and Apoptotic Pathways Targeted Cancer Therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Ray, S.; Hussein, M.A.; Srkalovic, G.; Almasan, A. Role of Apo2L/TRAIL and Bcl-2-family proteins in apoptosis of multiple myeloma. Leuk. Lymphoma 2003, 44, 1209–1214. [Google Scholar] [CrossRef]
- Zhang, C.; Ni, J.; Fan, W.; Hou, J. Nutlin-3 Promotes TRAIL-Induced Liver Cancer Cells Apoptosis by Activating p53 to Inhibit bcl-2 and Surviving Expression. Ann. Clin. Lab. Sci. 2022, 52, 601–610. [Google Scholar]
- Shankar, S.; Srivastava, R.K. Enhancement of therapeutic potential of TRAIL by cancer chemotherapy and irradiation: Mechanisms and clinical implications. Drug Resist. Updates 2004, 7, 139–156. [Google Scholar] [CrossRef]
- Herbst, R.S.; Mendolson, D.S.; Ebbinghaus, S.; Gordon, M.S.; O’Dwyer, P.; Lieberman, G.; Ing, J.; Kurzrock, R.; Novotny, W.; Eckhardt, G. A phase I safety and pharmacokinetic (PK) study of recombinant Apo2L/TRAIL, an apoptosis-inducing protein in patients with advanced cancer. J. Clin. Oncol. 2006, 24, 3013. [Google Scholar] [CrossRef]
- Lim, S.M.; Kim, T.H.; Jiang, H.H.; Park, C.W.; Lee, S.; Chen, X.; Lee, K.C. Improved biological half-life and anti-tumor activity of TNF-related apoptosis-inducing ligand (TRAIL) using PEG-exposed nanoparticles. Biomaterials 2011, 32, 3538–3546. [Google Scholar] [CrossRef]
- Yuan, X.-F.; Peng, H.-W.; Ding, Y.-H.; Yan, C.-H.; Zhang, Y.-J.; Yang, M.; Xiong, D.-S. Gene therapy in B-NHL cell line using adenovirus-mediated transfer of secretable trimeric TRAIL gene expression driven by CD20 promoter. Exp. Hematol. 2013, 41, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Clodi, K.; Wimmer, D.; Li, Y.; Goodwin, R.; Jaeger, U.; Mann, G.; Gadner, H.; Younes, A. Expression of tumour necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) receptors and sensitivity to TRAIL-induced apoptosis in primary B-cell acute lymphoblastic leukaemia cells. Br. J. Haematol. 2000, 111, 580–586. [Google Scholar] [PubMed]
- Labusca, L.; Herea, D.D.; Mashayekhi, K. Stem cells as delivery vehicles for regenerative medicine-challenges and perspectives. World J. Stem Cells 2018, 10, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Shokat, Z.; Ali, A.; Ahmed, U.; Javed, M.R.; Qasim, M.; Tariq, M.; Ahmed, M.R.; Masoud, M.S. Mesenchymal Stem Cells: From Regeneration to Drug Delivery Systems. Crit. Rev. Ther. Drug Carr. Syst. 2021, 38, 33–73. [Google Scholar] [CrossRef] [PubMed]
- Spano, C.; Grisendi, G.; Golinelli, G.; Rossignoli, F.; Prapa, M.; Bestagno, M.; Candini, O.; Petrachi, T.; Recchia, A.; Miselli, F.; et al. Soluble TRAIL Armed Human MSC As Gene Therapy For Pancreatic Cancer. Sci. Rep. 2019, 9, 1788. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Li, S.; Li, Z.; Peng, H.; Yuan, X.; Jiang, L.; Zhang, Y.; Fan, D.; Hu, X.; Yang, M.; et al. Human umbilical cord mesenchymal stem cells as vehicles of CD20-specific TRAIL fusion protein delivery: A double-target therapy against non-Hodgkin’s lymphoma. Mol. Pharm. 2013, 10, 142–151. [Google Scholar] [CrossRef]
- Pitti, R.M.; Marsters, S.A.; Ruppert, S.; Donahue, C.J.; Moore, A.; Ashkenazi, A. Induction of apoptosis by Apo-2 ligand, a new member of the tumor necrosis factor cytokine family. J. Biol. Chem. 1996, 271, 12687–12690. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yoshida, T.; Zhang, B. TRAIL induces endocytosis of its death receptors in MDA-MB-231 breast cancer cells. Cancer Biol. Ther. 2009, 8, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Fang, B. Mechanisms of resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther. 2005, 12, 228–237. [Google Scholar] [CrossRef]
- Luciano, R.L.; Brewster, U.C. Kidney involvement in leukemia and lymphoma. Adv. Chronic Kidney Dis. 2014, 21, 27–35. [Google Scholar] [CrossRef]
- Jacobs, P. Splenomegaly and acute lymphoblastic leukaemia. South Afr. Med. J. 1981, 59, 171. [Google Scholar]
- Shen, C.-J.; Chan, T.-F.; Chen, C.-C.; Hsu, Y.-C.; Long, C.-Y.; Lai, C.-S. Human umbilical cord matrix-derived stem cells expressing interferon-β gene inhibit breast cancer cells via apoptosis. Oncotarget 2016, 7, 34172–34179. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Chen, Y.; Lu, L.; Hu, X.; Shao, C.; Zhang, Y.; Zhou, X.; Zhou, Y.; Wu, L.; Liu, R.; et al. Human umbilical cord blood-derived mesenchymal stem cells producing IL15 eradicate established pancreatic tumor in syngeneic mice. Mol. Cancer Ther. 2014, 13, 2127–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.H.; Moon, A.R.; Chang, J.H.; Bae, J.; Choi, J.M.; Lee, S.H.; Kim, T.-H. Erratum to: Potentiation of TRAIL killing activity by multimerization through isoleucine zipper hexamerization motif. BMB Rep. 2017, 50, 647. [Google Scholar] [PubMed]
- Liu, T.M. Stemness of Mesenchymal Stem Cells. J. Stem Cell Ther. Transplant. 2017, 1, 071–073. [Google Scholar] [CrossRef] [Green Version]
- Szegezdi, E.; O’Reilly, A.; Davy, Y.; Vawda, R.; Taylor, D.L.; Murphy, M.; Samali, A.; Mehmet, H. Stem cells are resistant to TRAIL receptor-mediated apoptosis. J. Cell. Mol. Med. 2009, 13, 4409–4414. [Google Scholar] [CrossRef]
- De Miguel, D.; Lemke, J.; Anel, A.; Walczak, H.; Martinez-Lostao, L. Onto better TRAILs for cancer treatment. Cell Death Differ. 2016, 23, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Cronier, L.; Crespin, S.; Strale, P.-O.; Defamie, N.; Mesnil, M. Gap junctions and cancer: New functions for an old story. Antioxid. Redox Signal. 2009, 11, 323–338. [Google Scholar] [CrossRef]
- Kouzi, F.; Zibara, K.; Bourgeais, J.; Picou, F.; Gallay, N.; Brossaud, J.; Dakik, H.; Roux, B.; Hamard, S.; Le Nail, L.-R.; et al. Disruption of gap junctions attenuates acute myeloid leukemia chemoresistance induced by bone marrow mesenchymal stromal cells. Oncogene 2020, 39, 1198–1212. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Lan, Y.-W.; Huang, S.-M.; Yen, C.-C.; Chen, W.; Wu, W.-J.; Staniczek, T.; Chong, K.-Y.; Chen, C.-M. Human amniotic fluid mesenchymal stem cells attenuate pancreatic cancer cell proliferation and tumor growth in an orthotopic xenograft mouse model. Stem Cell Res. Ther. 2022, 13, 235. [Google Scholar] [CrossRef]
- Vahedi PMoghaddamshahabi, R.; Webster, T.J.; Calikoglu Koyuncu, A.C.; Ahmadian, E.; Khan, W.S.; Jimale Mohamed, A.; Eftekhari, A. The Use of Infrapatellar Fat Pad-Derived Mesenchymal Stem Cells in Articular Cartilage Regeneration: A Review. Int. J. Mol. Sci. 2021, 22, 9215. [Google Scholar] [CrossRef] [PubMed]
- Schilling, T.; Nöth, U.; Klein-Hitpass, L.; Jakob, F.; Schütze, N. Plasticity in adipogenesis and osteogenesis of human mesenchymal stem cells. Mol. Cell. Endocrinol. 2007, 271, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Halberg, N.; Khan, M.; Magalang, U.J.; Scherer, P.E. Selective inhibition of hypoxia-inducible factor 1α ameliorates adipose tissue dysfunction. Mol. Cell. Biol. 2013, 33, 904–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Li, X.; Yang, L.; Eckel-Mahan, K.; Tong, Q.; Gu, X.; Kolonin, M.G.; Sun, K. Transient Overexpression of Vascular Endothelial Growth Factor A in Adipose Tissue Promotes Energy Expenditure via Activation of the Sympathetic Nervous System. Mol. Cell. Biol. 2018, 38, e00242-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raulf, N.; El-Attar, R.; Kulms, D.; Lecis, D.; Delia, D.; Walczak, H.; Papenfuss, K.; Odell, E.; Tavassoli, M. Differential response of head and neck cancer cell lines to TRAIL or Smac mimetics is associated with the cellular levels and activity of caspase-8 and caspase-10. Br. J. Cancer 2014, 111, 1955–1964. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Zhong, X.; Dai, Q.; Li, K.; Zhang, W.; Wang, J.; Zhao, Y.; Shen, J.; Xiao, Z.; Xing, H.; et al. Human Umbilical Cord MSC Delivered-Soluble TRAIL Inhibits the Proliferation and Promotes Apoptosis of B-ALL Cell In Vitro and In Vivo. Pharmaceuticals 2022, 15, 1391. https://doi.org/10.3390/ph15111391
Chen F, Zhong X, Dai Q, Li K, Zhang W, Wang J, Zhao Y, Shen J, Xiao Z, Xing H, et al. Human Umbilical Cord MSC Delivered-Soluble TRAIL Inhibits the Proliferation and Promotes Apoptosis of B-ALL Cell In Vitro and In Vivo. Pharmaceuticals. 2022; 15(11):1391. https://doi.org/10.3390/ph15111391
Chicago/Turabian StyleChen, Fangshan, Xianmei Zhong, Qian Dai, Kuo Li, Wei Zhang, Jie Wang, Yueshui Zhao, Jing Shen, Zhangang Xiao, Hongyun Xing, and et al. 2022. "Human Umbilical Cord MSC Delivered-Soluble TRAIL Inhibits the Proliferation and Promotes Apoptosis of B-ALL Cell In Vitro and In Vivo" Pharmaceuticals 15, no. 11: 1391. https://doi.org/10.3390/ph15111391