
The Role of Habitat Heterogeneity in Structuring Mangrove Bird Assemblages

Abstract
:1. Introduction
2. Methods
2.1. Study Sites, Vegetation and Bird Sampling Methods
2.2. Habitat Heterogeneity Measurements
2.3. Data Analysis
2.3.1. Principle Component Analysis
2.3.2. Generalised Linear Models
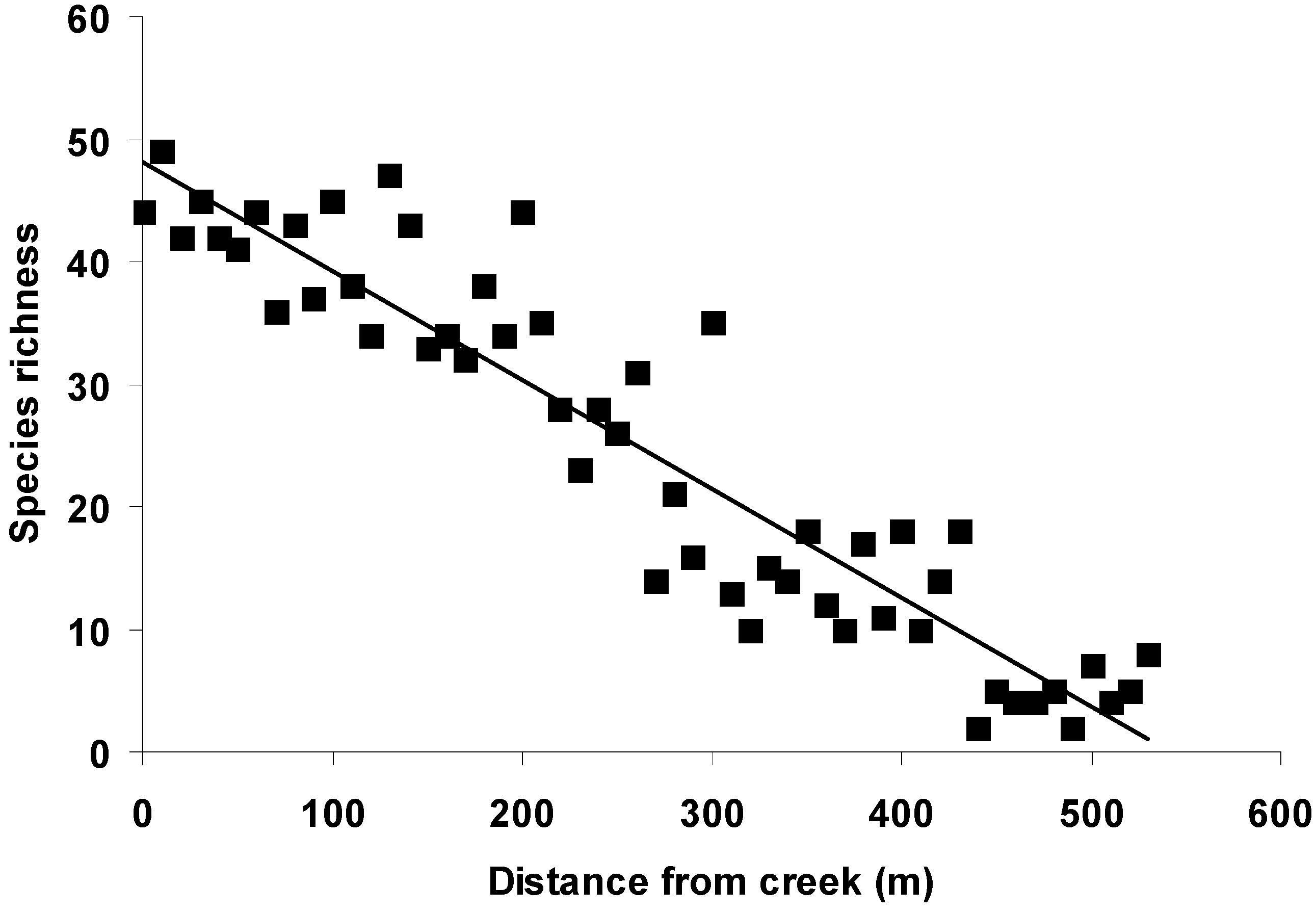
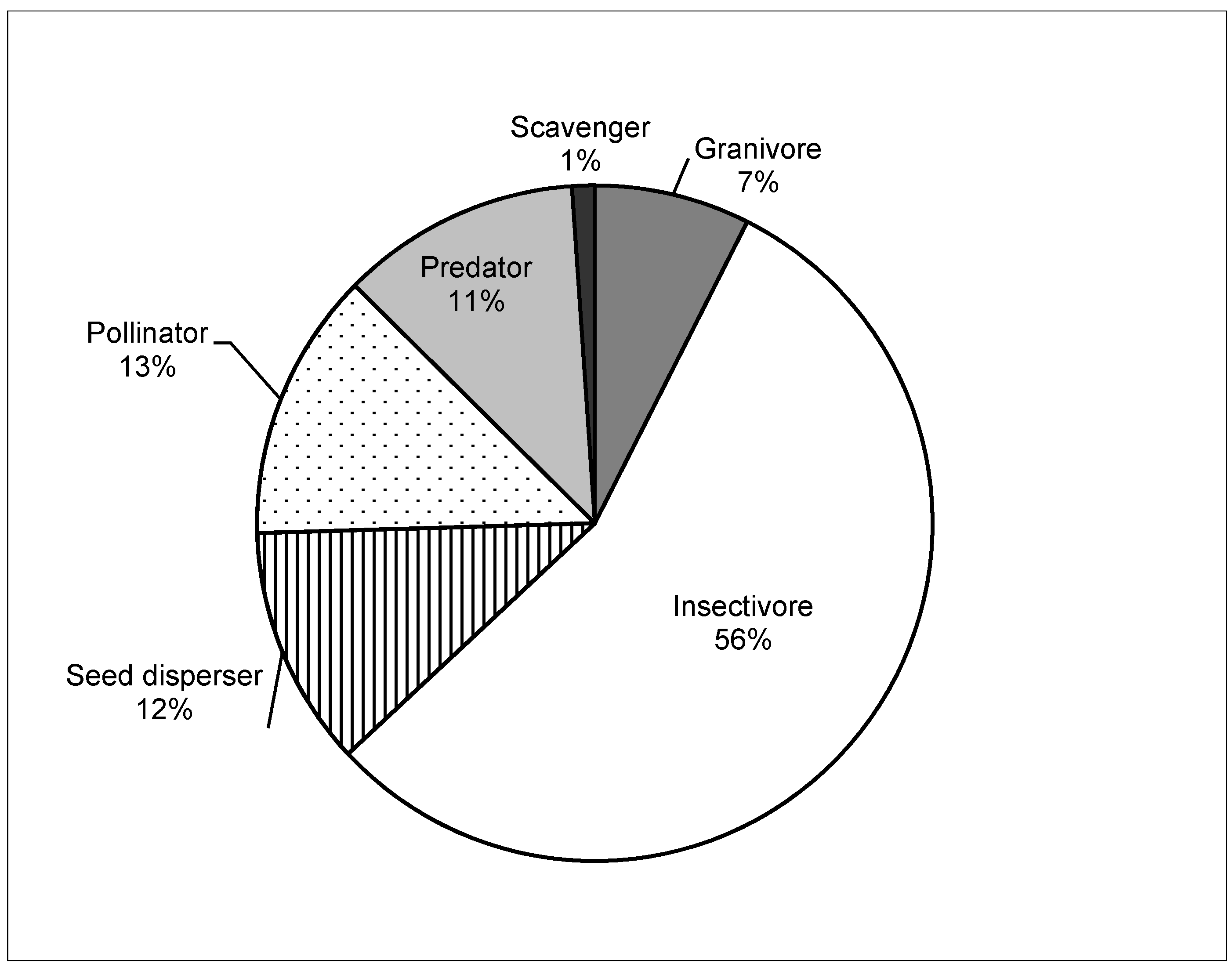
3. Results
3.1. Habitat Heterogeneity



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat Variable | BC | BV | CCR | CDNP | EAB | EAH | EAT | EP | LLA | LNR | MDL | NGLF | RC | Range | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Transect length (m) | 820 | 410 | 440 | 2110 | 420 | 750 | 830 | 560 | 560 | 800 | 60 | 260 | 800 | 60–2110 | |
| Density by DBH class (stems/0.025 ha) | |||||||||||||||
| DBH > 10 | 3.8 | 1.7 | 3.3 | 1.65 | 1.2 | 2.3 | 1.0 | 2.0 | 2.5 | 2.8 | 3.1 | 2.2 | 2.1 | 1.2–3.6 | |
| DBH > 2 | 49.5 | 73.1 | 33.0 | 45.3 | 45.0 | 18.0 | 51.5 | 16.8 | 18.5 | 40.0 | 8.0 | 20.0 | 19.0 | 8–73.1 | |
| DBH < 2 | 848.6 | 626.0 | 379.8 | 944.4 | 823.8 | 264.8 | 822.2 | 83.1 | 61.7 | 89.6 | 0 | 72.0 | 669.7 | 0–944.4 | |
| Density by height class (stems/0.025 ha) | |||||||||||||||
| HC1 (0–2) | 775.1 | 540.6 | 309.7 | 968.2 | 934. | 265.0 | 806.7 | 106.0 | 74.2 | 94.4 | 0 | 72.2 | 370.6 | 0–968.2 | |
| HC2 (2–4) | 29.8 | 57.9 | 14.5 | 17.2 | 26.5 | 11.1 | 48.6 | 6.6 | 1.0 | 27.20 | 0.8 | 7.0 | 24.5 | 0.8–58.0 | |
| HC3 (4–6) | 3.9 | 2.0 | 5.0 | 1.8 | 3.9 | 2.3 | 7.7 | 3.0 | 5.0 | 2.80 | 2.4 | 1.4 | 1.5 | 1.4–7.7 | |
| HC4 (6–8) | 1.5 | 0.1 | 2.0 | 0.7 | 0.2 | 0.7 | 1.4 | 0.8 | 2.1 | 0 | 2.1 | 1.0 | 0.3 | 0–2.02 | |
| HC5 (8–10) | 1.6 | 0 | 0.4 | 0.2 | 0 | 0 | 0.6 | 0.7 | 0.6 | 0 | 0.5 | 0.2 | 0.4 | 0–1.6 | |
| HC6 (10–12) | 1.6 | 0 | 0.10 | 0 | 0 | 0 | 0 | 0.1 | 1.5 | 0 | 1.1 | 0.3 | 0.2 | 0–1.6 | |
| HC7 (12–14) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.5 | 0 | 0 | 0 | 0 | 0–0.5 | |
| Vertical profile (%) | |||||||||||||||
| VP (0–2 m) | 45.0 | 4.00 | 37.0 | 50.5 | 36.6 | 42.0 | 41.1 | 45.6 | 20.2 | 40.1 | 21.1 | 29.2 | 45.6 | 20.2–50.5 | |
| VP (2–4 m) | 40.3 | 44.9 | 39.8 | 33.2 | 45.1 | 51.1 | 40.3 | 31.4 | 30.3 | 38.8 | 28.7 | 24.4 | 37.9 | 24.4–51.0 | |
| VP (4–6 m) | 32.6 | 26.5 | 32.6 | 20.7 | 28.3 | 36.6 | 25.3 | 29.1 | 36.4 | 16.4 | 45.0 | 30.7 | 24.7 | 16.4–45.0 | |
| VP (6–8 m) | 25.5 | 5.0 | 19.8 | 11.5 | 10.0 | 10.8 | 12.9 | 27.1 | 29.7 | 0 | 45.9 | 28.5 | 18.2 | 10.0–45.7 | |
| VP (8–10 m) | 12.9 | 0 | 10.0 | 5.7 | 0 | 0 | 9.1 | 14.8 | 35.8 | 0 | 42.1 | 18.0 | 1.0. | 0–42.1 | |
| VP (10–12 m) | 6.4 | 0 | 6.6 | 0 | 0 | 0 | 0 | 0 | 23.9 | 0 | 8.1 | 7.0 | 5.0 | 0–23.9 | |
| VP (12–14 m) | 0 | 0 | 3.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0–3.3 | |
| Grass cover (%) | 16.1 | 0 | 18.9 | 9.8 | 13.5 | 0 | 0 | 21.1 | 21.7 | 38.5 | 0 | 0 | 0 | 0–38.5 | |
| Canopy cover (%) | 49.3 | 43.3 | 44.2 | 43.5 | 46.1 | 46.9 | 41.1 | 43.7 | 39.6 | 40.8 | 44.3 | 39.8 | 46.4 | 39.6–49.3 | |
| Plant species richness | 25.0 | 7.0 | 27.0 | 9.0 | 11.0 | 9.0 | 8.0 | 27.0 | 7.0 | 3.0 | 5.0 | 7 | 7.0 | 3.0–27.0 | |
| Saltpan size (ha) | 9.2 | 0 | 0.5 | 17.4 | 0.7 | 3.1 | 3.6 | 0.8 | 2.2 | 8.0 | 0 | 0 | 1.8 | 0–17.4 | |
| No. of pencil roots (Avicennia sp.) | 620.0 | 216.0 | 385.6 | 522.6 | 112.0 | 1212.2 | 435.6 | 2637.0 | 2308.6 | 4227.2 | 1166.4 | 1762.4 | 1699.6 | 112–2637 | |
| No. of knee roots (Bruguiera sp.) | 123.4 | 159.4 | 13.6 | 79.8 | 609.6 | 0 | 214.8 | 38.1 | 27.2 | 0 | 150.4 | 0 | 108.0 | 0–609.6 | |
| No. of zones | 5 | 4 | 5 | 4 | 4 | 5 | 5 | 5 | 4 | 1 | 2 | 3 | 5.0 | 1–5 | |
| *No of garbage items | 0.2 | 4.4 | 1.4 | 0.1 | 0 | 0.9 | 0.2 | 1.5 | 3.4 | 5.5 | 3.0 | 0.3 | 9.8 | 0–5.5 | |
| *No of tracks | 0.1 | 0 | 0.3 | 0 | 0.3 | 4.5 | 0 | 3.5 | 0.4 | 1.0 | 0.5 | 0 | 0.03 | 0–4.5 | |
| *Bare ground (%) | 43.1 | 38.6 | 36.9 | 41.2 | 40.6 | 39.4 | 43.9 | 45.8 | 40.3 | 45.5 | 48.9 | 38.6 | 44.7 | 37.0–48.9 | |
| *Vegetation diversity (Fisher’s α) | 6.2 | 1.4 | 6.2 | 1.4 | 1.9 | 1.7 | 1.4 | 7.8 | 1.8 | 0.7 | 2.1 | 1.8 | 1.2 | 0.65–6.19 | |
3.2. Habitat Heterogeneity and Bird Species Composition
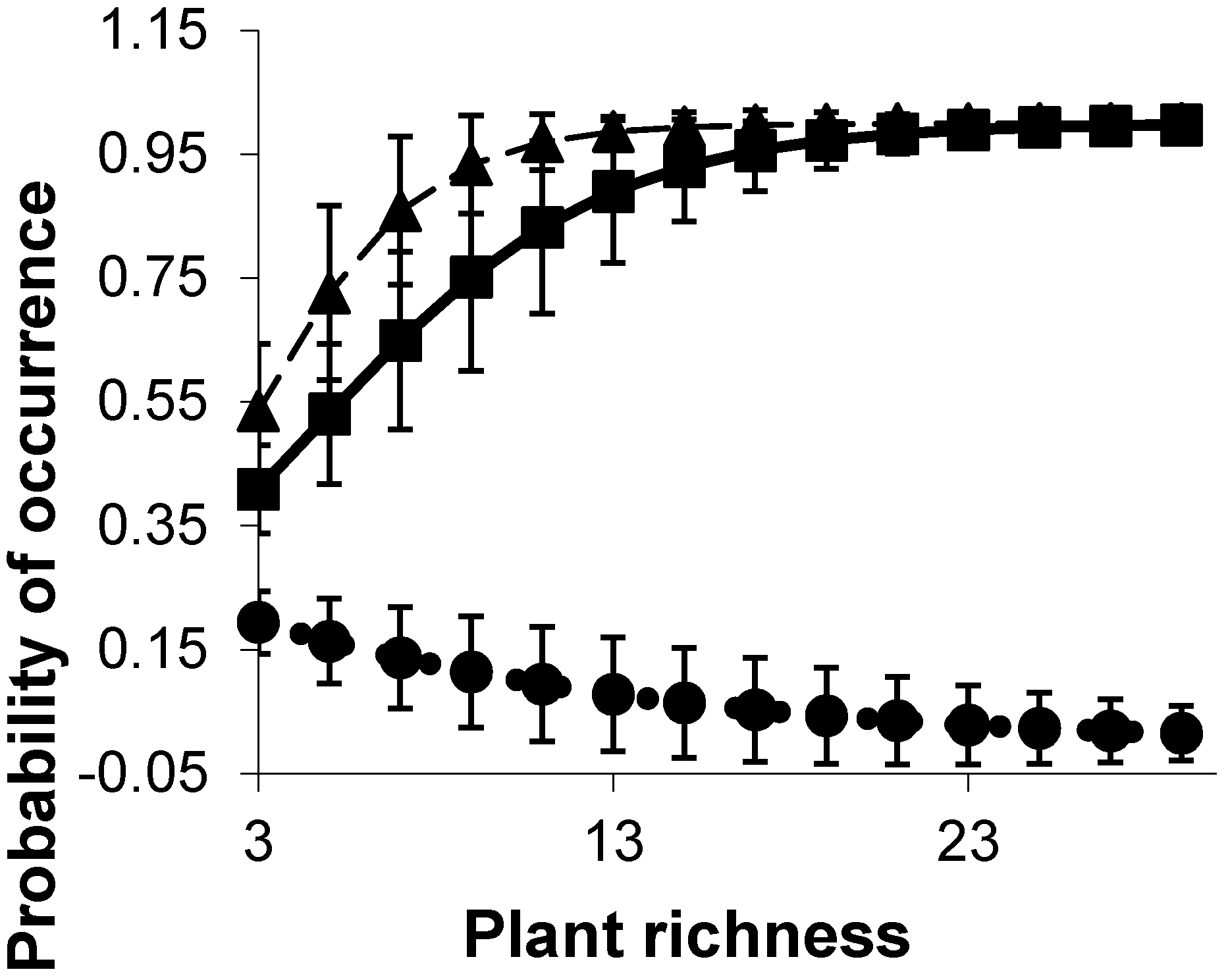
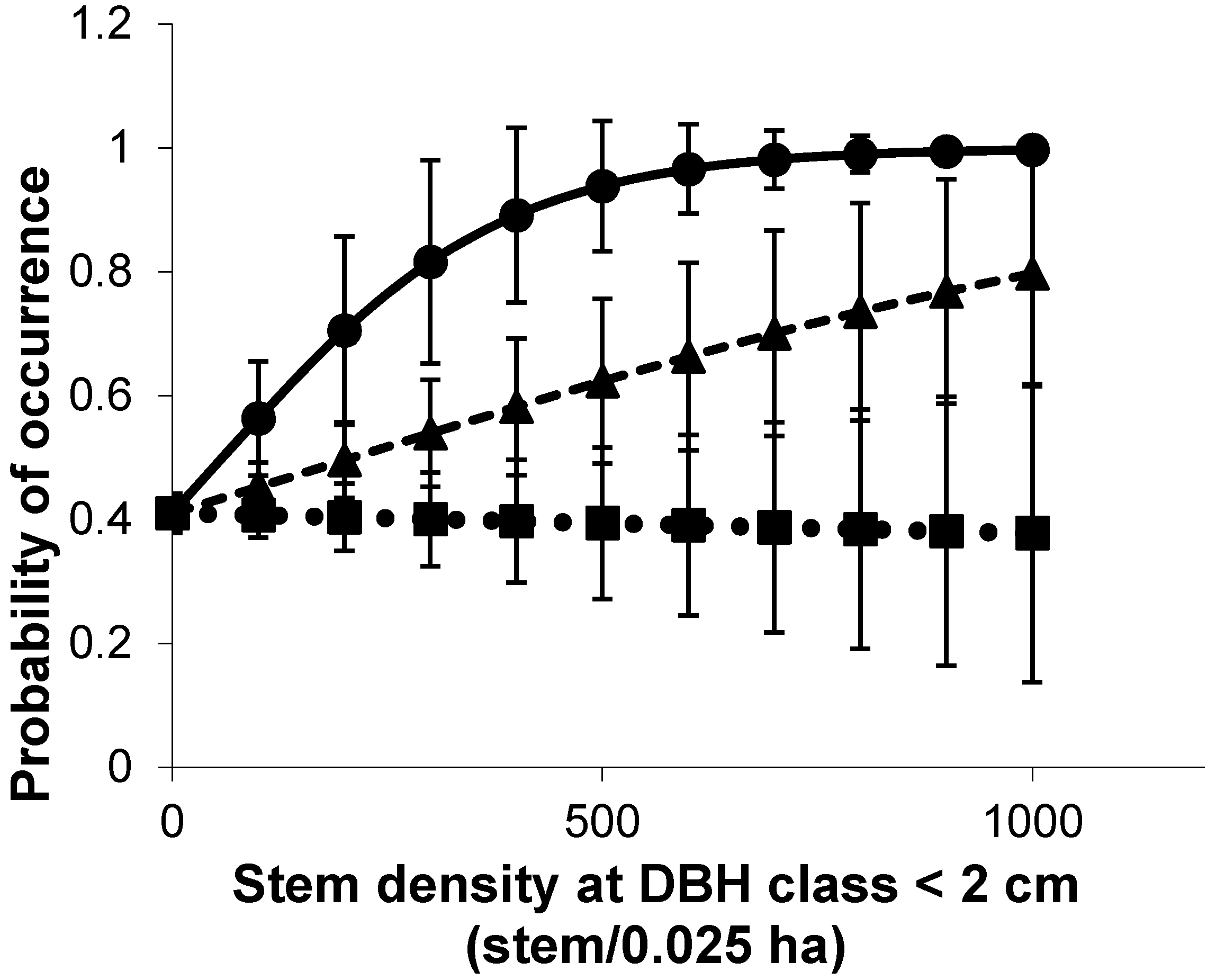
3.3. Habitat Heterogeneity Predictors of Bird Species Richness and Density
| Category | Eigenvalue | % of Variance | Broken-Stick Eigenvalue | % of Variables Selected | No. of Species |
|---|---|---|---|---|---|
| Overall Species | 15.4 | 38.7 | 4.8 | 29 | 70 |
| Interior species | 13.2 | 41.0 | 4.7 | 38 | 61 |
| Mangrove dependent species (MDS) | 4.7 | 65.5 | 3.0 | 21 | 12 |
| Species density | 13.3 | 34.3 | 4.8 | 38 | 70 |
| (a) Interior Species | (b) Species Density | (c) Overall Species Richness | (d) MDS Richness | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variables | Axis | R2 | Variables | Axis | R2 | Variables | Axis | R2 | Variables | Axis | R2 |
| DBH < 2 | 1 | 0.38 | VP 4–6 | 1 | 0.37 | HC5 | 1 | 0.48 | DBH < 2 | 1 | 0.28 |
| HC1 | 1 | 0.38 | VP 12–14 | 1 | 0.32 | Zones | 1 | 0.38 | HC1 | 1 | 0.27 |
| Zones | 1 | 0.28 | Zones | 1 | 0.43 | Plant richness | 1 | 0.60 | VP 0–2 | 1 | 0.36 |
| Saltpan | 1 | 0.36 | Plant richness | 1 | 0.49 | Canopy cover | 1 | 0.34 | Saltpan | 1 | 0.68 |
| VP 4–6 | 2 | 0.48 | Canopy cover | 1 | 0.32 | DBH > 10 | 1 | 0.28 | DBH > 2 | 2 | 0.25 |
| VP 6–8 | 2 | 0.31 | Pencil root | 1 | 0.31 | Grass (%) | 2 | 0.35 | |||
| Saltpan | 2 | 0.51 | DBH > 2 | 2 | 0.27 | Pencil root | 2 | 0.48 | |||
| Grass (%) | 2 | 0.41 | Grass (%) | 2 | 0.64 | ||||||
| Pencil root | 2 | 0.27 | Pencil root | 2 | 0.31 | ||||||
| Model | ΔAICc | −2ln(L(θ)) | AICc | K | wi | Explained Deviance |
|---|---|---|---|---|---|---|
| (a) Overall species richness | ||||||
| Plant richness | 0 | 7.6 | 12.8 | 2 | 0.21 | 38.5 |
| Canopy cover | 1.6 | 9.3 | 14.5 | 2 | 0.09 | 25.3 |
| Zones | 1.8 | 9.5 | 14.7 | 2 | 0.08 | 23.9 |
| HC5 | 2.5 | 10.1 | 15.3 | 2 | 0.06 | 18.7 |
| Zones + grass | 2.5 | 6.7 | 15.3 | 3 | 0.06 | 46.5 |
| Plant richness + canopy cover | 2.6 | 6.8 | 15.5 | 3 | 0.06 | 45.4 |
| Plant richness + grass | 3.0 | 7.2 | 15.8 | 3 | 0.05 | 42.5 |
| Grass | 3.1 | 10.8 | 16.0 | 2 | 0.04 | 13.5 |
| Plant richness + pencil root | 3.4 | 7.6 | 16.2 | 3 | 0.04 | 39.2 |
| (b) Interior species richness | ||||||
| Grass | 0 | 3.0 | 8.2 | 2 | 0.11 | 19.7 |
| Zones | 0.03 | 3.1 | 8.3 | 2 | 0.11 | 19.0 |
| VP 4–6 | 0.1 | 3.2 | 8.4 | 2 | 0.10 | 16.4 |
| DBH < 2 | 0.4 | 3.4 | 8.6 | 2 | 0.09 | 10.0 |
| VP 6–8 | 0.4 | 3.4 | 8.6 | 2 | 0.09 | 9.3 |
| HC1 | 0.5 | 3.5 | 8.7 | 2 | 0.09 | 7.3 |
| Saltpan | 0.6 | 3.7 | 8.9 | 2 | 0.08 | 3.1 |
| Zones + grass | 2.4 | 1.9 | 10.6 | 3 | 0.03 | 48.8 |
| (c) MDS richness | ||||||
| DBH < 2 | 0 | 2.5 | 7.7 | 2 | 0.16 | 45.0 |
| HC1 | 0.1 | 2.5 | 7.7 | 2 | 0.15 | 43.1 |
| VP 0–2 | 0.1 | 2.5 | 7.7 | 2 | 0.15 | 42.9 |
| DBH > 2 | 0.7 | 3.1 | 8.3 | 2 | 0.12 | 30.3 |
| DBH < 2 + VP 0–2 | 3.1 | 2.0 | 10.7 | 3 | 0.04 | 54.3 |
| DBH < 2 + saltpan | 3.5 | 2.4 | 11.1 | 3 | 0.03 | 45.3 |
| DBH < 2 + HC1 | 3.5 | 2.4 | 11.1 | 3 | 0.03 | 45.2 |
| (d) Species density | ||||||
| Pencil root | 0 | 14.6 | 20.0 | 2 | 0.21 | 38.9 |
| Pencil root + plant richness | 1.2 | 12.1 | 21.1 | 3 | 0.12 | 49.4 |
| Grass | 1.6 | 16.2 | 21.5 | 2 | 0.10 | 32.3 |
| Pencil root + zones | 2.1 | 13.1 | 22.1 | 3 | 0.07 | 45.4 |
| Pencil root + grass | 2.9 | 13.8 | 22.8 | 3 | 0.05 | 42.3 |
| Pencil root + VP 12–14 | 3.0 | 14.0 | 23.0 | 3 | 0.05 | 41.5 |
| Pencil root + canopy cover | 3.0 | 14.0 | 23.0 | 3 | 0.05 | 41.5 |
| w+ | Unconditional SE | |
|---|---|---|
| Overall richness | ||
| Plant richness | 0.49 | 0.0048 |
| %Grass | 0.29 | 0.0023 |
| %Canopy cover | 0.28 | 0.0026 |
| Zones (n) | 0.22 | 0.0023 |
| HC5 | 0.11 | 0.0015 |
| DBH >10 cm | 0.09 | 0.0010 |
| Pencil root | 0.08 | 0.0009 |
| Interior species | ||
| %Grass | 0.25 | 0.0019 |
| Zones (n) | 0.25 | 0.0019 |
| VP 4–6 m | 0.17 | 0.0017 |
| DBH < 2 cm | 0.15 | 0.0015 |
| VP 6–8 m | 0.12 | 0.0014 |
| HC1 | 0.12 | 0.0014 |
| Saltpan (ha) | 0.09 | 0.0012 |
| Pencil root | 0.05 | 0.0005 |
| MDS richness | ||
| DBH < 2 cm | 0.33 | 0.0046 |
| VP 0–2 m | 0.29 | 0.0043 |
| HC1 | 0.26 | 0.0043 |
| DBH > 2 cm | 0.22 | 0.0034 |
| Saltpan (ha) | 0.11 | 0.0020 |
| Species density | ||
| Pencil root | 0.79 | 0.0047 |
| %Grass | 0.28 | 0.0024 |
| Plant richness | 0.20 | 0.0021 |
| DBH > 2 cm | 0.16 | 0.0018 |
| VP 4–6 m | 0.12 | 0.0015 |
| Zones (n) | 0.10 | 0.0013 |
| %Canopy cover | 0.06 | 0.0008 |
| VP 12–14 m | 0.06 | 0.0008 |


4. Conclusion and Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; p. 203. [Google Scholar]
- Tews, K.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The important of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Acevedo, M.A.; Aide, T.M. Bird community dynamics and habitat associations in karsts, mangrove and Pterocarpous forest fragments in an urban zone in Puerto Rico. Caribb. J. Sci. 2008, 44, 402–416. [Google Scholar]
- Lindenmayer, D.B.; Wood, J.T.; McBurney, L.; Michael, D.; Crane, M.; MacGregor, C.; Montague-Drake, R. Comparing bird species richness and assemblage composition between montane ash eucalypt forest and cool temperate rainforest—An empirical study from Victoria, south-eastern Australia. Emu 2010, 110, 109–117. [Google Scholar] [CrossRef]
- Raman, T.R.S. Effects of habitat structure and adjacent habitats on birds in tropical rainforest fragments and shaded plantations in the Western Ghats, India. Biodivers. Conserv. 2006, 15, 1577–1607. [Google Scholar] [CrossRef]
- Trainor, C.R. Species richness, habitat use and conservation of birds of Alor Islands, Lesser Sundas, Indonesia. Emu 2005, 105, 127–135. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Tidemann, S.C.; Kerin, S. Birds in a tropical mosaic: The distribution of bird species in relation to vegetation patterns. Wildl. Res. 1988, 15, 171–196. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Fisher, A. Wildlife of Lancewood (Acacia shirlyi) thickets and woodlands in northern Australia. 2. Comparisons with other environments of the region (Acacia woodlands, Eucalyptus savanna woodlands and monsoon rainforest. Wildl. Res. 1995, 22, 413–443. [Google Scholar] [CrossRef]
- Metcalfe, K. The Biological Diversity, Recovery from Disturbance and Rehabilitation of Mangroves in Darwin Harbour, Northern Territory. Ph.D. Thesis, Charles Darwin University, Darwin, Australia, 2007. [Google Scholar]
- Woinarski, J.C.Z.; Fisher, A.; Brennan, K.; Morris, I.; Chatta, R. Patterns of bird species richness and composition on islands off Arnhem Land, Northern Territory, Australia. Austral Ecol. 2001, 26, 1–13. [Google Scholar]
- Connor, E.F.; McCoy, E.D. The statistics and biology of the species-area relationship. Am. Nat. 1979, 113, 791–833. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Pianka, E.R. On the optimal use of a patchy environment. Am. Nat. 1966, 100, 603–609. [Google Scholar] [CrossRef]
- Ford, J. Origin, evolution and speciation of birds specialized to mangroves in Australia. Emu 1982, 82, 12–23. [Google Scholar] [CrossRef]
- Berg, Å. Diversity and abundance of birds in relation to forest fragmentation, habitat quality and heterogeneity. Bird Stud. 1997, 44, 355–366. [Google Scholar]
- Cousin, J.A.; Phillips, R.D. Habitat complexity explains species-specific occupancy but not species richness in a Western Australian woodland. Aust. J. Zool. 2008, 56, 95–102. [Google Scholar] [CrossRef]
- Honkanen, M.; Roberge, J.-M.; Rajaärkkä, A.; Mönkkönen, M. Distangling the effects of area, energy and habitat heterogeneity on boreal forest bird species richness in protected areas. Glob. Ecol. Biogeogr. 2009, 18, 61–71. [Google Scholar]
- Bateman, B.L.; Kutt, A.S.; Vanderduys, E.P.; Kemp, J.E. Small-mammal species richness and abundance along a tropical altitudinal gradient: An Australian example. J. Trop. Ecol. 2010, 26, 139–149. [Google Scholar] [CrossRef]
- Cromsigt, J.P.G.M.; Prins, H.H.T.; Olff, H. Habitat heterogeneity as a driver of ungulate diversity and distribution patterns: Interaction of body mass and digestive strategy. Divers. Distrib. 2009, 15, 513–522. [Google Scholar] [CrossRef]
- Hunter, W.R.; Sayer, M.D.J. The comparative effects of habitat complexity on faunal assemblages of northern temperate artificial and natural reefs. ICES J. Mar. Sci. Adv. Access 2009, 66, 691–698. [Google Scholar] [CrossRef]
- Haddad, N.M.; Crutsinger, G.M.; Gross, K.; Haarstad, J.; Knops, J.M.H.; Tilman, D. Plant species loss decreases arthropod diversity and shift tropic structure. Ecol. Lett. 2009, 12, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Hodgkison, R.; Balding, S.T.; Zubaid, A.; Kunz, T.H. Habitat structure, wing morphology, and the vertical stratification of Malaysian fruit bats (Megachiroptera: Pteropodidae). J. Trop. Ecol. 2004, 20, 667–673. [Google Scholar] [CrossRef]
- Rebelo, H.; Brito, J.C. Bat guild structure and habitat use in the Sahara desert. Afr. J. Ecol. 2006, 45, 228–230. [Google Scholar] [CrossRef]
- MacArthur, R.H. Population ecology of some warblers of north eastern coniferous forest. Ecology 1958, 39, 599–619. [Google Scholar] [CrossRef]
- MacArthur, R.H.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- Hulbert, A.H. Species-energy relationships and habitat complexity in bird communities. Ecol. Lett. 2004, 7, 714–720. [Google Scholar] [CrossRef]
- Hulbert, A.H. Linking species-area and species-energy relationships in Drosophila microcosms. Ecol. Lett. 2006, 9, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Wethered, R.; Lawes, M.J. Matrix effects on bird assemblages in fragmented Afromontane forest in South Africa. Biol. Conserv. 2003, 114, 327–340. [Google Scholar] [CrossRef]
- Pavey, C.R.; Nano, C.E.M. Bird assemblages of arid Australia: Vegetation patterns have greater effect than disturbance and resource pulses. J. Arid Environ. 2009, 73, 634–342. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Whittaker, R.J.; Dawson, T.P. Avifaunal response to habitat fragmentation in the threatened littoral forest of south-eastern Madagascar. J. Biogeogr. 2004, 31, 1791–1807. [Google Scholar] [CrossRef]
- Woinarski, J.C.Z.; Brock, C.; Amstrong, M.; Hempel, C.; Cheal, D.; Brennan, K. Bird distribution in riparian vegetation in the extensive natural landscape of Australia’s tropical savanna: A broad-scale survey and analysis of a distributional data base. J. Biogeogr. 2000, 27, 843–868. [Google Scholar] [CrossRef]
- Johnstone, R.E. Mangroves and Mangrove Birds of Western Australia; Western Australian Museum: Perth, Australia, 1990. [Google Scholar]
- Noske, R.A. Abundance, zonation and foraging ecology of birds in mangroves of Darwin Harbour, Northern Territory. Wildl. Res. 1996, 23, 443–474. [Google Scholar] [CrossRef]
- Mohd-Azlan, J.; Noske, R.A.; Lawes, M.J. Avian species-assemblage structure and indicator bird species of mangroves in the Australian monsoon tropics. Emu 2012, 112, 287–297. [Google Scholar] [CrossRef]
- Mohd-Azlan, J.; Lawes, M.J. The effect of the surrounding landscape matrix on mangrove bird community assembly in north Australia. Biol. Conserv. 2011, 144, 2134–2141. [Google Scholar] [CrossRef]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling; Oxford University Press: Oxford, UK, 2001; p. 432. [Google Scholar]
- Sekercioglu, C.H. Increasing awareness of avian ecology function. Trends Ecol. Evol. 2006, 21, 455–471. [Google Scholar] [CrossRef]
- Kutt, A.S. Bird assemblage in a dune-mangrove mosaic, Cairns, Queensland. Aust. Zool. 2007, 34, 158–164. [Google Scholar] [CrossRef]
- Krüger, S.C.; Lawes, M.J. Edge effects at an induced forest-grassland boundary: Forest birds in the Ongoye Forest Reserve, KwaZulu-Natal. S. Afr. J. Zool. 1997, 32, 82–91. [Google Scholar]
- Kempton, R.A.; Wedderburn, R.W.M. A comparison of three measures of species diversity. Biometrics 1978, 34, 25–37. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity, 2nd ed.; Blackwell: Oxford, UK, 2004. [Google Scholar]
- Soetaert, K.; Heip, C. Sample-size dependence of diversity indices and the determination of sufficient sample size in a high-diversity deep-sea environment. Mar. Ecol. Press Ser. 1990, 59, 305–307. [Google Scholar] [CrossRef]
- Hutchings, P.; Saenger, P. Ecology of Mangroves; University of Queensland Press: Brisbane, Australia, 1987; p. 388. [Google Scholar]
- August, P.V. The role of habitat complexity and heterogeneity in structuring tropical mammal communities. Ecology 1983, 64, 1495–1507. [Google Scholar] [CrossRef]
- Brocklehurst, P.; Lewis, D.; Napier, D.; Lynch, D. Northern Territory Guidelines and Field Methodology for Vegetation Survey and Mapping; Northern Territory; Department of Natural Resources Environment and Arts: Darwin, Australia, 2007. [Google Scholar]
- Comley, B.W.T.; McGuinness, K.A. Above and below ground biomass, of four common northern Australian mangroves. Aust. J. Bot. 2005, 53, 431–436. [Google Scholar] [CrossRef]
- Schulte, R.P.O.; Lantinga, E.A.; Hawkins, M.J. A new family of Fisher-curves estimates Fisher’s alpha more accurately. J. Theor. Biol. 2005, 232, 305–313. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data; 4.34; MjM Software: Gleneden Beach, OR, USA, 1999. [Google Scholar]
- McCune, B.; Grace., J.B. Analysis of Ecological Communities; MjM Software: Gleneden Beach, OR, USA, 2002; p. 304. [Google Scholar]
- Jafari, M.; Chahouki, M.A.Z.; Tavili, A.; Azarnivand, H.; Amiri, G.Z. Effective environmental factors in the distribution of vegetation types in Poshtkouh rangelands of Yazd Province (Iran). J. Arid Environ. 2004, 56, 627–641. [Google Scholar] [CrossRef]
- VSN International GenStat for Windows, 13th ed; VSN International: Hemel Hempstead, UK, 2010.
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; Chapman and Hall: London, UK, 1989. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Kullback-Leiber information as a basis for strong inference in ecological studies. Wildl. Res. 2001, 28, 111–119. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and inference: A Practical Information—Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Anderson, D.R. Model Based Inference in the Life Sciences: A Primer on Evidence; Springer Verlag: New York, NY, USA, 2008; p. 184. [Google Scholar]
- MacArthur, R.H.; MacArthur, J.W.; Preer, J. On bird species diversity II prediction of bird cencus from habitat measurements. Am. Nat. 1962, 96, 167–774. [Google Scholar] [CrossRef]
- Rotenberry, J.T. The role of habitat in avian community composition: Physiognomy or floristics? Oecologia 1985, 67, 213–217. [Google Scholar] [CrossRef]
- Seymour, C.L.; Dean, W.R.J. The influence of changes in habitat structure on the species composition of bird assemblages in the southern Kalahari. Austral Ecol. 2010, 35, 581–592. [Google Scholar] [CrossRef]
- Tassicker, A.L.; Kutt, A.S.; Vanderduys, E.; Mangru, S. The effects of vegetation structure on the birds in a tropical savanna woodland in north-eastern Australia. Rangel. J. 2006, 28, 139–152. [Google Scholar] [CrossRef]
- Lees, A.C.; Peres, C.A. Rapid avifaunal collapse along the Amazonian deforestation frontier. Biol. Conserv. 2006, 133, 198–211. [Google Scholar] [CrossRef]
- Coupland, G.T. The Ecological Interaction between Insects and Mangroves in Darwin Harbor, Australia. Ph.D. Thesis, Northern Territory University, Darwin, Australia, 2002. [Google Scholar]
- Mulyani, Y.A. Reproductive Ecology of Tropical Mangrove-Dwelling Warblers: The Role of Nest Predation, Brood Parasitism, and Food Limitation. Ph.D. Thesis, Charles Darwin University, Darwin, Australia, 2004. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd-Azlan, J.; Noske, R.A.; Lawes, M.J. The Role of Habitat Heterogeneity in Structuring Mangrove Bird Assemblages. Diversity 2015, 7, 118-136. https://doi.org/10.3390/d7020118
Mohd-Azlan J, Noske RA, Lawes MJ. The Role of Habitat Heterogeneity in Structuring Mangrove Bird Assemblages. Diversity. 2015; 7(2):118-136. https://doi.org/10.3390/d7020118
Chicago/Turabian StyleMohd-Azlan, Jayasilan, Richard A. Noske, and Michael J. Lawes. 2015. "The Role of Habitat Heterogeneity in Structuring Mangrove Bird Assemblages" Diversity 7, no. 2: 118-136. https://doi.org/10.3390/d7020118